6
Art and Dementia: How Degeneration of Some Brain Regions Can Lead to New Creative Impulses
The study of individuals with dementia has largely been focused on describing the deficits and changes in behavior that accompany neural degeneration. Most studies have compared affected individuals with their healthy counterparts, and emphasize loss rather than change. Certainly, the loss of cognitive functions such as memory, language, and decision-making devastates patients and their families. But in some cases, patients and their families develop coping strategies that lead to the paradoxical emergence of new and positive behaviors such as gardening, painting, and other forms of personal expression. These cases provide insight not only into the capacity of the mind to adapt and survive, but also into the creative process and the drive to communicate. As artistic expression is deeply personal and subjective, the study of creativity in patients with dementia will need to focus on the individual as well as the group.
Defining and measuring creativity has proven to be a monumental task. Taken together with the considerable individual differences in response to brain damage, and the subjectivity of artistic expression, evidence unambiguously demonstrating increased creativity in patients with degenerative dementia is scarce. Here, we focus on the emergence of previously unrecognized artistic and musical creativity in the context of neurodegenerative illnesses such as frontotemporal dementia (FTD) (Miller et al., 1998, 2000). Understanding which skills are retained or even emerge in the setting of dementia not only aids in differential diagnosis, but can also help clinicians design appropriate treatment options and diminish the frustration that patients and their caregivers experience on a daily basis by finding activities that bring satisfaction and pleasure to patients. Furthermore, as age is the biggest risk factor in the development of dementia, highlighting the positive changes that can accompany progressive neural degeneration may also aid in decreasing the prevalence of prejudicial attitudes toward the elderly.
Neural Circuits Affected in Neurodegenerative Disease
Current directions in neuroscience research have begun to move away from regional specialization and toward an understanding of how cognitive and other functions are served by neural circuits. Neurodegenerative diseases such as Alzheimer’s disease (AD) and FTD strike specific circuits; damage is not limited to one isolated region. Degeneration is not immediate, however, and the progressive nature of the disease provides neuroscientists with the opportunity to understand how circuits might cope with loss, and how the brain might reorganize functional specialization. In some patients, the reorganization can be striking, not only in terms of regions taking over certain functions, but also in the behavioral changes that reflect the mind’s adaptation to functional loss. The study of neural plasticity is entering a golden age and our understanding of the neurobiology of creativity will certainly benefit from this new focus.
One of the recent advances in cognitive neuroscience has provided the tools with which scientists can observe connectivity between regions during different mental states. Fox and colleagues (2005) demonstrated that while certain brain regions such as the frontal and parietal cortices act in concert during tasks demanding focused attention, other regions show correlated activity only when the mind is “at rest,” or not actively engaged in a task (Fox et al., 2005). The delineation of functional connectivity during different cognitive tasks is particularly fruitful in patients with neurodegenerative disease, as it allows scientists to understand the context in which new behaviors emerge and how functional reorganization with disease progression might occur.
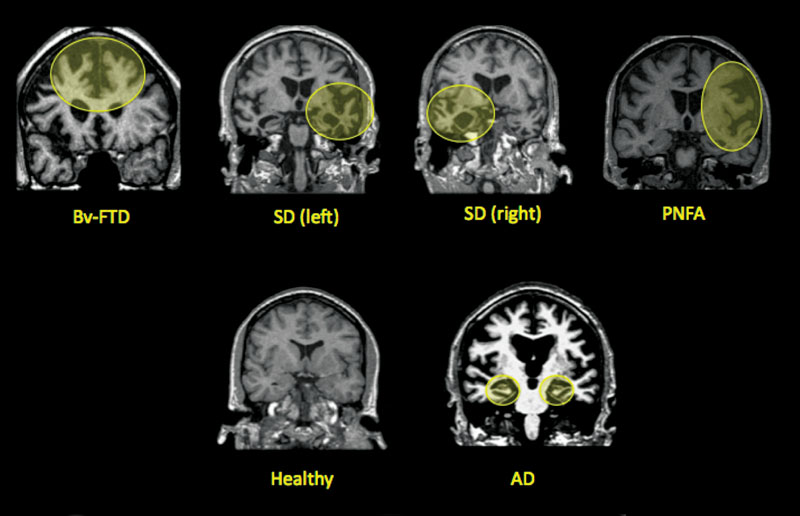
Functional connectivity analyses in patients with neurodegenerative diseases have begun to demonstrate that these diseases target specific large-scale neural networks (Seeley et al., 2009). AD is characterized by episodic memory dysfunction, and results in atrophy in the “default mode network” (Fox et al., 2005), including the posterior cingulate/precuneus, medial temporal lobe, and lateral temporoparietal regions. FTD is an umbrella term for a group of diseases that result in progressive degeneration of the frontal and temporal lobes. These disorders are categorized into three subtypes: behavioral-variant frontotemporal dementia (bvFTD) causes primarily behavioral symptoms, including disinhibition, apathy, emotional blunting, lack of insight, and poor decision making, and anterior cingulate, frontoinsular, striatal, and frontopolar degeneration; semantic dementia (SD) patients show a loss of word and object meaning, or conceptual knowledge, with primarily left-temporal pole and subgenual cingulate atrophy; and progressive nonfluent aphasia (PNFA) leads to nonfluent, effortful, and agrammatic speech, caused by degeneration in the left frontal operculum, dorsal anterior insula, and precentral gyrus (see figure 6.1; for a review, see Boxer & Miller, 2005).
Figure 6.1
Coronal MRI images showing areas affected by neurodegenerative disease. BvFTD: behavioral-variant frontotemporal dementia; SD: semantic dementia, left: left hemisphere, right: right hemisphere; PNFA: progressive nonfluent aphasia; AD: Alzheimer’s disease.
Networks Involved in Creativity
Although the relationship between these specific patterns of network degeneration and the emergence of creativity in specific patients remains murky, there is overlap between the networks affected by these diseases and those thought to underlie creativity. In particular, neuroscientists investigating creativity have moved away from the idea that creativity is primarily a right-hemisphere function (Bogen & Bogen, 1988), and toward the understanding that the interaction between the frontal and temporal lobes might be critical for generating and acting on creative impulses (Flaherty, 2005).
Providing a theoretical framework through which the neural networks involved in different types of creative output may be understood, Dietrich (2004) distinguishes between cognitive and emotional content, and spontaneous and deliberate creative processes. These two dimensions yield four distinct classes: insights can be cognitive and deliberate, cognitive and spontaneous, emotional and deliberate, or emotional and spontaneous. Furthermore, each class of creative thought is likely subserved by a different pattern of network interactions (see figure 6.2).
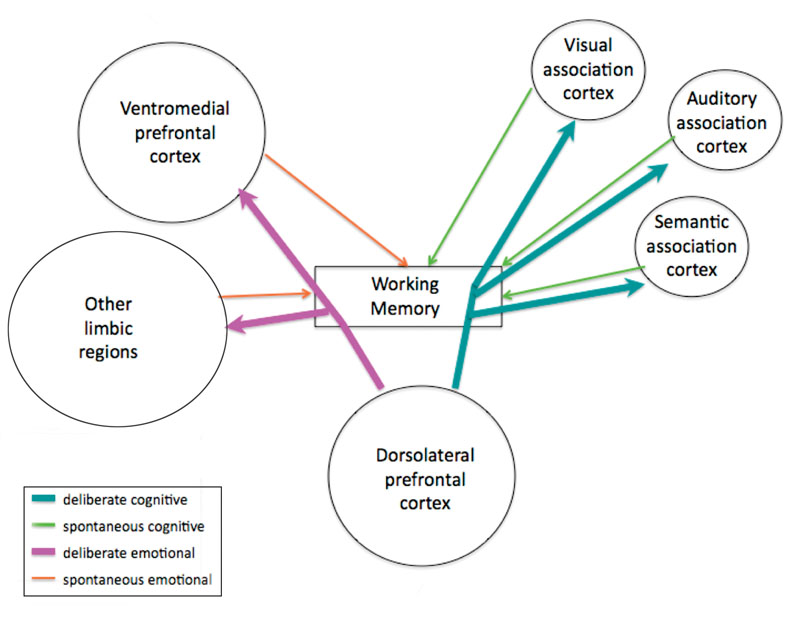
Figure 6.2
Types of creative output and the interactions between neural networks that support them. Thicker arrows indicate control, thinner arrows indicate information flow.
Cognitive deliberate insights likely rely heavily on dorsolateral prefrontal cortex (dlPFC) and its connections with domain-specific association cortex. Cognitive spontaneous insights reflect metabolic activity in the posterior association cortex. These thoughts seem to involve the basal ganglia, given its role in automatic behaviors, and enter consciousness, or working memory, during the periodic down-regulation of the frontal attentional system. Deliberate emotional insights require an interaction between the ventromedial PFC and the limbic system, including the amygdala and cingulate cortex. Spontaneous emotional insights are thought to be driven primarily by the activity of the limbic system, when the PFC is deactivated. Often, during biologically significant events and/or intense emotional experiences, information processed in emotion regions takes priority and overrides voluntary attention. Understanding how each of these types of insights is subserved by specific neural circuits will aid in the interpretation of the paradoxical facilitation of creativity with the degeneration of certain neural circuits.
The frontal cortex, in particular, receives information from regions throughout the brain, including motor, sensory, and emotion areas, and via reciprocal connections controls attention, language, memory, and other cognitive functions such as self-construct (Keenan et al., 2000), self-reflective consciousness (Courtney et al., 1998), complex social function (Damasio et al., 1994), cognitive flexibility, planning, willed action, source memory, and theory of mind. With changes in frontal neuroanatomy, one expects to see effects on creativity, although the exact nature of these effects is difficult to predict given the complexity of frontal lobe function.
Paradoxical Facilitation of Creativity with Neurodegenerative Disease
There have been some cases of patients in whom new or preserved musical or visual artistic abilities have been described in the setting of FTD (Miller et al., 1998, 2000). Given the central role that the frontal lobes play in creativity, the emergence of these abilities is often linked to degeneration of the anterior temporal lobe, particularly in the left hemisphere, as opposed to the frontal lobes. These patients, classified as having SD or PNFA, show progressive loss of conceptual knowledge or other language skills, and their creative output tends to be devoid of verbal or symbolic content (Miller et al., 2000).
These patients tend to paint realistic landscapes, animals, or detailed geometric designs that do not seem to contain abstract meaning beyond the superficial visual characteristics. Musicians continue to perform music, but when words are set to music, as is the case in popular songs, these patients do not use the words to express themselves. For example, one individual sang almost every day, and his primary pleasurable activity was leading elderly patients in a hospice setting in sing-alongs. He could sing any popular song in a large songbook, but when the words of a famous folk song (“My Bonny Lies Over the Ocean”) were altered to the point of being incomprehensible, he did not miss a beat (Sacks, 2007). In our experience, however, much of the work created by these patients contains some recollections from the past and tends to avoid verbal or symbolic representations.
To explore the connection between anterior temporal lobe degeneration and visual art, Rankin and colleagues (2007) used standardized tests to probe creative cognition and novel tests of visual art creation in patients with dementia. Patients with both bvFTD and SD produced artwork that was rated as more bizarre and distorted than patients with AD and healthy age-matched controls. SD patients were more likely than participants from any of the other groups to produce drawings devoid of meaning, and to choose conventional or obvious markings, such as closing an open-ended figure in standardized visuospatial creativity tests, which involved creating drawings based on incomplete meaningless doodles. In contrast, the paintings that they created, and indeed much of the artwork from this patient group, was striking in the aesthetic dimension, characterized by the unorthodox use of vivid, unconventional colors and intricate, repetitive geometrical designs, underscoring the unique perception as well as the obsessive nature of their work. This pattern of findings—that SD patients are less creative on standardized visuospatial creativity tests but less conventional in their self-generated depictions of the world—underscores the difficulty in measuring and categorizing creativity. Great artists share their unique subjective experience of the world in a way that both resonates with and surprises their audience, and by that measure, some of these SD patient artists produced work of substantial artistic merit.
In line with their language deficits, however, SD patients showed decreased fluency, originality, and elaboration in concert with a tendency toward ending the test prematurely on verbal tests of creativity. These findings suggest that the enhancements in creativity that may accompany a particular degenerative pattern are likely to remain domain specific, at least in the case of artists with neurodegenerative disease. Just as mastery of skills in one domain such as music can improve creative output in that domain but not necessarily transfer to other artistic realms, so too might the paradoxical facilitation of creativity with neurological disease remain tied to a single type of output. Dean Simonton (1994) introduced the idea that creative works that meet both the uniqueness and appropriateness criteria are almost always a function of a highly developed skill used in a novel fashion. Before a novel creative masterpiece is produced, artists spend years developing the necessary skills in painting, music, writing, and science (Ericsson, 2008).
The development of these necessary skills is enabled by the plastic nature of the brain. Cortical plasticity facilitates fine-tuning of motor and sensory skills and allows the formation of novel associations that make up the building blocks of creativity, and has been observed in both the frontal and medial temporal lobes. For example, extensive practice of a musical instrument results in reorganization of motor and sensory regions involved in the activity, with a larger proportion of cortical real estate devoted to the sensory and motor processing of the relevant body part (Elbert & Rockstroh, 2004). The flexible and dynamic nature of brain regions such as the hippocampus and prefrontal cortex, where plasticity is most pronounced, enables the formation of new associations, which form the basis of creativity. When, as might be the case in patients with SD, these regions are disrupted in a progressive manner, the brain’s efforts to compensate for loss of function might underlie the emergence of a new obsession and/or creative behavior.
The Case of Anne Adams
Recently, Seeley et al. (2008) described the work and disease progression of a visual artist, Anne Adams, who suffered from PNFA. Ms. Adams was a biologist by profession but painted as a hobby for much of her working life. As her disease progressed, her artistic drive increased so much that eventually painting became her primary daily activity. It is interesting to note that she might have chosen to spend her time engaged in any number of hobbies, but, like many similar patients, she chose to focus on creating art in the visual domain. Likewise, an SD patient who was a lawyer by profession and a musician by avocation chose to produce visual art when he became ill, rather than resorting to music, which would have been more familiar, or some other related activity (Miller et al., 1998).
Underscoring the paradoxical facilitation of creativity in the setting of disease, Anne’s most stunning works of art were produced at a time when the disease had already made a significant impact on her brain function. In PNFA, the main site of atrophy is the left fronto-opercular cortex, leaving patients with effortful, nonfluent, and apractic speech (Gorno-Tempini et al., 2004), and difficulties with grammar and articulation. Adding another dimension to the story is the observation that one of her masterpieces was an elaborate painting inspired by Maurice Ravel’s (1875–1937) famous Bolero (Seeley et al., 2008) composed at a time when Ravel himself is thought to have been suffering from a progressive aphasia. Ravel composed his most famous piece to the rhythms of the Moorish-Spanish dance, in the early stages of his disease, at age 53. The Bolero is a compulsively repetitive/perseverative piece (Amaducci et al., 2002), with the recurrence of a simple melodic theme, accompanied by a cyclical and simple bass line. Ravel considered the Bolero as “a piece for orchestra without music.” “I’m going to try and repeat [the theme] a number of times without any development, gradually increasing the orchestra as best I can” (Orenstein, 1991). Anne Adams was inspired to paint the Bolero, although she was not aware that she and Ravel shared a similar illness. She painted the piece in a precise and compulsive fashion, representing each repetition of the melodic theme as a sequence of squares in a set of rows, with the height of each row signifying the increasing texture and volume of the orchestra. A retrospective of her art production throughout her lifetime shows a movement away from symbolic representations and toward detailed, obsessive interest in geometric patterns (see figure 6.3).
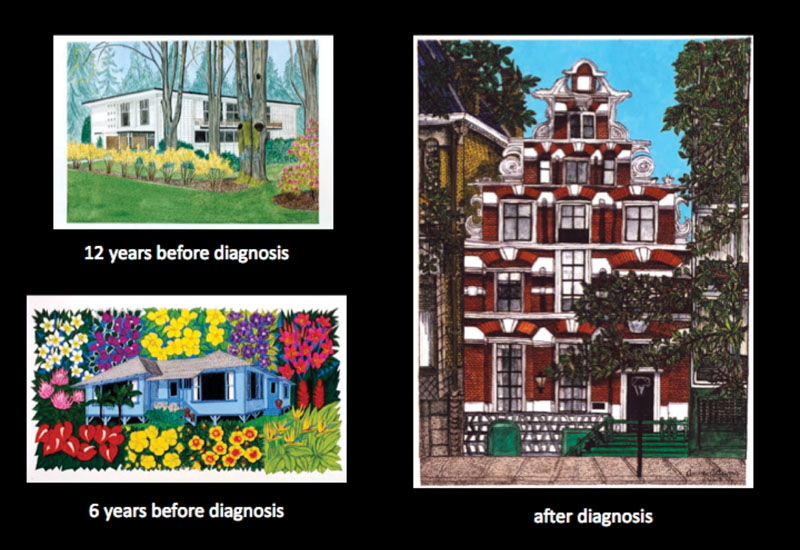
Figure 6.3
Representative artwork by Anne Adams as her disease progressed. Used with permission.
The case of Anne Adams is fascinating not only from the behavioral perspective, but also in terms of the structural and functional characteristics of her brain. Although her disease caused progressive degeneration primarily in the frontal cortex, Seeley and colleagues (2008) found a surprising surplus of gray matter volume and a functional enhancement in activation in heteromodal associative (intraparietal sulcus and superior parietal lobule: IPS/SPL) and polymodal (superior temporal sulcus: STS) neocortex in the right hemisphere. These areas have been shown to be involved in visuomotor search and attentional control (Corbetta & Shulman, 2002) and sensory transcoding, which are necessary for sight-reading music (Sergent et al., 1992). Professional musicians also have greater brain volume in these regions (Gaser & Schlaug, 2003).
This in-depth case study led to a larger study of visual attention in SD patients, with the goal of shedding some light on the perceptual experience of patients who might develop visual artistic creativity in the setting of neurodegenerative disease. An exploratory study of visual attention in SD patients unearthed striking differences between areas of focus of SD patients, healthy controls, and other patient groups during freeform viewing of complex images. SD patients tended to avoid areas that grabbed the attention of other patients and healthy controls, such as the vanishing point in images that emphasized perspective, and instead seemed to be interested in dense textures and complex color patterns. Some representative examples may be seen in figure 6.4. Comparing data from nine healthy controls, nine AD patients, nine FTD patients, and seven SD patients, we found no significant differences in terms of the mean number of fixation points, the mean length of the fixation time, or the distance between fixation points (length of saccades), but patients did differ from controls and each other in terms of where in the image they tended to spend the most time looking (I. V. Viskontas & B. L. Miller, unpublished raw data).

Figure 6.4
Mean eye movement fixation density plots from nine healthy controls (Cont), nine bvFTD patients (FTD), nine AD patients (AD), and seven SD patients (SD).
Intrigued by these findings, we designed a study to investigate how our patient groups might differ on a standard visual search task. To that end, we adapted a task pioneered by Anne Treisman and colleagues (1977) in which participants are asked to search an array of items for a target (Viskontas et al., 2011). When the target and distracters have distinguishable features, the search is conducted in parallel, leading to the experience of the target “popping out” of the array, and increasing the number of distracters has little or no effect on response time or accuracy. When, however, the target and distracters share features, the search is conducted by serially investigating each item, and therefore the response time in the task correlates positively with the number of items in the array, and participants are more likely to make errors as the number of distracters increases (Treisman & Gelade, 1980). In addition to recording response time and accuracy, we also monitored participants’ eye movements and used voxel-based morphometry (VBM) to look for differences in brain volume that might account for differences in behavior.
Congruent with our observations that SD patients show preserved, if not enhanced, performance in tasks relying heavily on visual search such as completing jigsaw puzzles, playing solitaire, finding coins, and gardening, we found that SD patients were not only as accurate as healthy controls during both pop-out and serial search (both groups performed at ceiling), but they were significantly faster under the most difficult serial search condition. By contrast, AD patients showed significant impairments even under the easiest conditions and chance performance during serial search with an array size of thirty items. FTD patients were significantly worse than controls only in the most difficult serial search condition, and, like controls, their response times increased with more distracters in serial search. SD patients, in contrast, showed no increase in response time when the display size was doubled from fifteen to thirty items in the serial search condition, and found the target more quickly than all other groups even while looking at a smaller proportion of the image. That is, in the context of visual search tasks, SD patients may have a more efficient search strategy that allows them to ignore distracters and find the target more quickly than healthy people.
Performance on the serial search task correlated with gray matter volume in a dorsal frontoparietal network (including a region in the superior parietal lobe, the precuneus, the middle frontal gyrus, and higher visual regions in the occipital lobe) that is commonly associated with visual attention and which is spared in the setting of SD. The serial search task relies heavily on the binding of visual features into a coherent trace and the region of the parietal lobe that correlated with better performance in this task has also been shown to result in binding problems when it is damaged, as in Balint’s syndrome or hemispatial neglect (Chan et al., 2003). Furthermore, hyperactive binding, as seen in individuals with synesthesia, has also been correlated with neural activity and more gray matter in the parietal lobe (Weiss & Fink, 2009). There was no appreciable difference in gray matter in this region between SD patients and healthy controls, supporting the idea that the enhancement in visual search function seen in SD patients results from increased functional connectivity rather than structural differences. This hypothesis has recently gained more support as enhancements in dorsal parietal network connectivity have been reported in patients with FTD (Zhou et al., 2010). Taken together, the results of this comprehensive study of visual search in SD patients suggests that the paradoxical enhancement of visual artistry seen in certain patients might reflect a shift in the attentional space from language to vision, resulting from an enhancement in connectivity in the dorsal parietal network.
Motivation and Creativity
Despite their progressive neurodegeneration, SD and PNFA patients who showed enhanced creativity with disease progression share many features with historical figures with great creativity. For example, they show more interest and/or obsessions with their subjects, they neglect social and occupational responsibilities in favor of their art production, and they continue to produce work even in the absence of any encouragement or support from others (Miller et al., 1998). This observation leads to questions regarding the underlying motivating factors in these patients, and artists in general, to spend time creating art. Notably, patients with emergent new or preserved old artistic skills generally do not show extensive frontal lobe degeneration (Miller et al., 2000). This observation is in line with the wealth of research on creativity in neurologically intact individuals, which suggests that certain components of frontal lobe function are critical for the many organizational and motivational components of creativity (Chávez-Eakle et al., 2007).
The involvement of the PFC in creativity underscores the important role that motivation plays in generating and sustaining creative behavior. While creativity is described as a virtue in a wide variety of careers, the creative life is generally not financially rewarding. In fact, a common view among social psychologists is that monetizing creative output can destroy or disrupt the very process that leads to innovation (Amabile et al., 1986). The relationship between reward and creativity remains murky, as studies have found both negative and positive relationships between incentives and output (Eisenberger & Cameron, 1996). What’s clear, however, is that some tasks seem to be rewarding in and of themselves; as Harlow et al. (1950) first demonstrated, even monkeys will repeatedly solve mechanical puzzles even when the solution does not lead to any food or other extrinsic reward. Some activities are simply intrinsically rewarding, and many of these activities seem to have some component of creativity such as spontaneity, making new associations, or using one’s imagination. A meta-analysis conducted by Cameron and Pierce (1994) suggests that while verbal rewards increase the time that individuals spend on intrinsically rewarding tasks, tangible rewards such as money, gold stars, or awards tend to decrease the time spent on task once those rewards are removed. Furthermore, this effect is seen only when tangible rewards are expected, and disappears when the rewards are spontaneously given after the task is accomplished. One of the most widely accepted explanations for these effects is that adding tangible extrinsic rewards to the equation shifts the focus from the task itself and onto the narrow scope of the goal (Amabile et al., 1986).
Flow
Mihaly Csikszentmihalyi refers to the pleasurable state that motivates the engagement of these activities as “flow” (Csikszentmihalyi, 1991), and suggests that nine separable coexisting factors are necessary to create it: (1) the delineation of clear goals, wherein expectations and rules are discernible, goals are attainable, and goals align appropriately with one’s skill set and abilities; (2) a high degree of concentration on a limited number of items; (3) a loss of the feeling of self-consciousness, via the merging of action and awareness; (4) a distorted sense of subjective time; (5) direct and immediate feedback, which makes successes and failures apparent, so that behavior can be adjusted as needed; (6) a balance between ability level and challenge, such that the activity is neither too easy nor too difficult; (7) a sense of personal control over the situation or activity; (8) the activity is intrinsically rewarding, so that action feels effortless; and finally, (9) the activity is all-encompassing.
The dominance of the visual domain and the perseverative or obsessive behaviors characteristic of patients with SD can also be seen in the large proportion (~25 percent) of these patients for whom the completion of jigsaw puzzles becomes an important, or even the primary activity of daily living (Green & Patterson, 2009), likely because they are able to achieve “flow” by working on these puzzles. In a controlled study of jigsaw puzzle activity, Green and Patterson (2009) found that SD patients have preserved jigsaw puzzle completion skills, sometimes even above and beyond the performance of age-matched controls. In particular, these patients showed better performance than their healthy counterparts in completing “reality-disrupted” puzzles, in which expectations based on knowledge of the real world can interfere with puzzle completion, and on “grain” jigsaw puzzles, where conceptual knowledge does not benefit performance. Most encouraging, from a clinical standpoint, SD patients, who often exhibit flat affect and demeanor, have been observed to display signs of pleasure and pride during the completion of jigsaw puzzles, even if the activity was not a primary one for that particular person, suggesting that this task might be a good candidate for enabling the pleasurable state of flow which might drive the quest to create in some patients with SD.
The Role of Inhibition in Creativity
Besides having the necessary skills in a given field, creative individuals often lack inhibition in both behavior and cognition (Martindale & Hines, 1975; Martindale, 1999), a trait seen in many patients who show artistic expression in the setting of neurodegenerative disease. Anecdotally, creative people describe themselves as lacking self-control, and the creative process is sometimes described as effortless and without deliberation (Csikszentmihalyi, 1996). In line with the network view of neurodegenerative disease and higher cognitive function, Miller and colleagues (2000) have suggested that the mechanism by which creativity is unleashed in the setting of left anterior temporal lobe degeneration might involve releasing right dorsal parietal network from inhibition. The psychiatric disorders that have been also associated with creativity, such as bipolar disorder, usually include impulsive behaviors in their symptomatology (Martindale, 1971). It follows, then, that decreased activation in the frontal lobes, particularly on the left side, might be the source of disinhibition that has been associated with creativity. Support for this idea can be found in a doctoral dissertation by Hudspeth (1985), who found that more creative people show higher amplitude frontal lobe theta wave activity, which presumably indicates lower frontal lobe function. These results are consistent with findings reported by Carlsson et al. (2000), who found that decreases in regional blood flow (rCBF) in both the left and right superior frontal lobes correlated with superior performance on the Alternate Uses Task. Taken together, these findings highlight the complexity of the creative process and demonstrate that whereas the frontal lobes are involved in creative cognition and the organization of deliberate creative behaviors as described in the model presented earlier in this chapter, many of the spontaneous aspects of creative output seem to be supported by regions outside of the frontal cortex. One might even go a step further and suggest that under some conditions the frontal lobes need to be “turned off” in order to facilitate the emergence of these spontaneous behaviors, or that these behaviors emerge during the periodic down-regulation of frontal lobe function in healthy individuals (Dietrich, 2004).
The hypothesis that deactivation of frontal cortex might enhance creativity was tested by Snyder and colleagues (2003), who used repetitive transcranial magnetic stimulation (rTMS), a noninvasive method of transiently deactivating parts of the cortex in awake humans. Indeed, rTMS of the left frontotemporal lobe enhanced the creative components of certain skills, such as drawing (though not necessarily drawing ability per se, but rather the ability to capture perspective, kinetics, and certain highlighted details) and the ability to detect commonly overlooked duplicate words while proofreading. These proofreading skills rely on similar perceptual and attentional processes that are enhanced in SD patients. Caution must be used to interpret these results, however, as the effects were seen only in a subset of participants (4/11 in drawing, 2/11 in proofreading). Nevertheless, the results are consistent with the general notion that deliberate language-based focus can inhibit spontaneous creative thinking and that the absence of these constraints may enable the dynamic processes which lead to the novel recombination of ideas (Bristol & Viskontas, 2006).
Summary
Progressive neurodegenerative disease that disrupts the interactions between the frontal and the temporal, parietal, and occipital lobes, or between the dominant and nondominant hemispheres, has been shown to affect creativity in a myriad of ways. Diminished language function via neurodegenerative diseases that target the left frontal or left anterior temporal lobes sometimes leads to the emergence of previously unrecognized visual and musical creativity, possibly by facilitating function in posterior brain regions. Down-regulation of frontal function may enable spontaneous creative insights, but as patients with frontal lobe dysfunction demonstrate, the frontal lobes are necessary for many components of creativity, including organization, monitoring, and other executive functions. The importance of studying the paradoxical facilitation of behaviors that can help patients achieve “flow” is underscored by the observation that patients who engage in creative activities display many signs of improved quality of life. It is important to note, however, that while the art created by patients with neurodegenerative disease might be hailed as more creative by the artistic community, the brain networks underlying the drive to make these choices may not necessarily be the same as those upon which healthy artists rely.
References
Amabile, T. M., Hennessey, B. A., & Grossman, B. S. (1986). Social influences on creativity: The effects of contracted-for reward. Journal of Personality and Social Psychology, 50, 14–23.
Amaducci, L., Grassi, E., & Boller, F. (2002). Maurice Ravel and right-hemisphere musical creativity: Influence of disease on his last musical works? European Journal of Neurology, 9, 75–82.
Bogen, J. E., & Bogen, G. M. (1988). Creativity and the corpus callosum. Psychiatric Clinics of North America, 11, 293–301.
Boxer, A. L., & Miller, B. L. (2005). Clinical features of frontotemporal dementia. Alzheimer Disease and Associated Disorders, 19, S3–S6.
Bristol, A. S., & Viskontas, I. V. (2006). Dynamic processes within associative memory stores: Piecing together the neural basis of creative cognition. In J. C. Kaufman & J. Baer (Eds.), Creativity, knowledge, and reason (pp. 60–80). Cambridge: Cambridge University Press.
Cameron, J., & Pierce, W. D. (1994). Reinforcement, reward, and intrinsic motivation: A meta-analysis. Review of Educational Research, 20, 51–61.
Carlsson, I., Wendt, P. E., & Risberg, J. (2000). On the neurobiology of creativity: Differences in frontal activity between high and low creative subjects. Neuropsychologia, 38, 873–885.
Chan, R. C., Robertson, I. H., & Crawford, J. R. (2003). An application of individual subtest scores calculation in the Cantonese version of the Test of Everyday Attention. Psychological Reports, 93, 1275–1282.
Chávez-Eakle, R. A., Graff-Guerrero, A., García-Reyna, J. C., Vaugier, V., & Cruz-Fuentes, C. (2007). Cerebral blood flow associated with creative performance: A comparative study. NeuroImage, 38, 519–528.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews: Neuroscience, 3, 201–215.
Courtney, S.M., Petit, L., Haxby, J.V., & Ungerleider, L.G. (1998). The role of prefrontal cortex in working memory: Examining the contents of consciousness. Philosophical Transactions of the Royal Society, Series B: Biological Sciences, 353, 1819–1828.
Csikszentmihalyi, M. (1991). Flow: The psychology of optimal experience. New York: Harper & Row.
Csikszentmihalyi, M. (1996). Creativity. New York: Harper Collins.
Damasio, H., Grabowski, T., Frank, R., Galaburda, A. M., & Damasio, A. R. (1994). The return of Phineas Gage: Clues about the brain from the skull of a famous patient. Science, 264, 1102–1105.
Dietrich, A. (2004). The cognitive neuroscience of creativity. Psychonomic Bulletin & Review, 11, 1011–1026.
Eisenberger, R., & Cameron, J. (1996). Detrimental effects of reward: Reality or myth? American Psychologist, 51, 1153–1166.
Elbert, T., & Rockstroh, B. (2004). Reorganization of human cerebral cortex: The range of changes following use and injury. Neuroscientist, 10, 129–141.
Ericsson, K. A. (2008). Deliberate practice and acquisition of expert performance: A general overview. Academic Emergency Medicine, 15, 988–994.
Flaherty, A. W. (2005). Frontotemporal and dopaminergic control of idea generation and creative drive. Journal of Comparative Neurology, 493, 147–153.
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen, D. C., & Raichle, M. E. (2005). The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proceedings of the National Academy of Sciences of the United States of America, 102, 9673–9678.
Gaser, C., & Schlaug, G. (2003). Brain structures differ between musicians and non-musicians. Journal of Neuroscience, 23, 9240–9245.
Gorno-Tempini, M. L., Dronkers, N. F., Rankin, K. P., Ogar, J. M., Phengrasamy, L., Rosen, H. J., et al. (2004). Cognition and anatomy in three variants of primary progressive aphasia. Annals of Neurology, 55, 335–346.
Green, H. A., & Patterson, K. (2009). Jigsaws: A preserved ability in semantic dementia. Neuropsychologia, 47, 569–576.
Harlow, H. F., Harlow, M. K., & Meyer, D. R. (1950). Learning motivated by a manipulation drive. Journal of Experimental Psychology, 40, 228–234.
Hudspeth, S. (1985). The neurological correlates of creative thought. Unpublished PhD dissertation. Los Angeles, CA: University of Southern California.
Keenan, J. P., Wheeler, M. A., Gallup, G. G., Jr., & Pascual-Leone, A. (2000). Self-recognition and the right prefrontal cortex. Trends in Cognitive Sciences, 4, 338–344.
Martindale, C. (1971). Degeneration, disinhibition, and genius. Journal of the History of the Behavioral Sciences, 7, 177–182.
Martindale, C. (1999). Biological bases of creativity. In R. J. Sternberg (Ed.), Handbook of creativity (pp. 137–152). Cambridge: Cambridge University Press.
Martindale, C., & Hines, D. (1975). Creativity and cortical activation during creative intellectual and EEG feedback tasks. Biological Psychology, 3, 91–100.
Miller, B. L., Boone, K., Cummings, J. L., Read, S. L., & Mishkin, F. (2000). Functional correlates of musical and visual ability in frontotemporal dementia. British Journal of Psychiatry, 176, 458–463.
Miller, B. L., Cummings, J., Mishkin, F., Boone, K., Prince, F., Ponton, M., et al. (1998). Emergence of artistic talent in frontotemporal dementia. Neurology, 51, 978–982.
Orenstein, A. (1991). The ballets of Maurice Ravel: Creation and interpretation. Burlington: Ashgate.
Rankin, K. P., Liu, A. A., Howard, S., Slama, H., Hou, C. E., Shuster, K., et al. (2007). A case-controlled study of altered visual art production in Alzheimer’s and FTLD. Cognitive and Behavioral Neurology, 20, 48–61.
Sacks, O. W. (2007). Musicophilia: Tales of music and the brain. New York: Alfred A. Knopf.
Seeley, W. W., Matthews, B. R., Crawford, R. K., Gorno-Tempini, M. L., Foti, D., Mackenzie, I. R., et al. (2008). Unravelling Bolero: Progressive aphasia, transmodal creativity, and the right posterior neocortex. Brain, 131, 39–49.
Seeley, W. W., Crawford, R. K., Zhou, J., Miller, B. L., & Greicius, M. D. (2009). Neurodegenerative diseases target large-scale human brain networks. Neuron, 62, 42–52.
Sergent, J., Zuck, E., Terriah, S., & MacDonald, B. (1992). Distributed neural network underlying musical sight-reading and keyboard performance. Science, 257, 106–109.
Simonton, D. K. (1994). Greatness: Who makes history and why. New York: Guilford Press.
Snyder, A. W., Mulcahy, E., Taylor, J. L., Mitchell, D. J., Sachdev, P., & Gandevia, S. C. (2003). Savant-like skills exposed in normal people by suppressing the left fronto-temporal lobe. Journal of Integrative Neuroscience, 2, 149–158.
Treisman, A., Sykes, M., & Gelade, G. (1977). Selective attention and stimulus integration. In S. Dornic (Ed.), Attention and performance VI. Hillsdale, NJ: Erlbaum.
Treisman, A. M., & Gelade, G. (1980). A feature integration theory of attention. Cognitive Psychology, 12, 97–136.
Viskontas, I. V., Boxer, A. L., Fesenko, J., Matlin, A., Heuer, H. W., Mirsky, J., et al. (2011). Visual search patterns in semantic dementia show paradoxical facilitation of binding processes. Neuropsychologia, 49, 468–478.
Weiss, P. H., & Fink, G. R. (2009). Grapheme-colour synaesthetes show increased grey matter volumes of parietal and fusiform cortex. Brain, 132, 65–70.
Zhou, J., Greicius, M. D., Gennatas, E. D., Growdon, M. E., Jang, J. Y., Rabinovici, G. D., et al. (2010). Divergent network connectivity changes in behavioural variant frontotemporal dementia and Alzheimer’s disease. Brain, 133, 1352–1367.