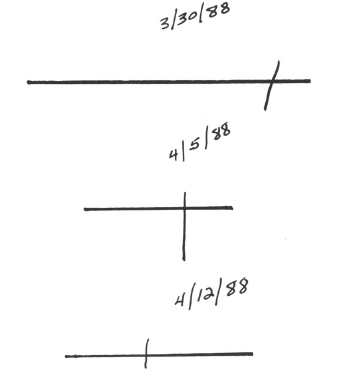
Fig. 8.1. Changes in line bisection over a two week period as tested at bedside.
Attentional Search in Unilateral Visual Neglect
Departments of Neurology and Psychiatry, University of California, Davis, and Veterans Administration Medical Center, Martinez, California, USA
University Hospital, Department of Neurology, Zurich, Switzerland
Introduction
Patients with severe visual neglect typically come to attention on a neurology ward because of their altered visual scanning behaviour. Eye deviations to the ipsilesional (i.e. intact) side of space and difficulty scanning towards the contralesional (i.e. neglected) side can be seen overtly, especially in the acute stages. Although the frequency of right and left neglect is controversial due to the confounding deficit of aphasia with left hemisphere damage (see Battersby, Bender, Pollock, & Kahn, 1956; De Renzi, 1982; Ogden, 1987; Zarit & Kahn, 1974), the greater severity of neglect with right hemisphere damage at least in the acute stages is well accepted (see De Renzi, 1982; Mesulam, 1981, 1983, 1985; Ogden, 1987; Roy et al., 1987). Acutely, patients with right hemisphere damage are more likely to deviate eyes, head and body towards their intact side than patients with left hemisphere damage. The overt manifestation of visual neglect clearly points to altered search of space. The major question we will address in this chapter is what cognitive mechanisms contribute to the overt scanning deficit?
Before proceeding, it is wise to keep in mind the vast variability inherent in neurological syndromes during the acute stage. Clinical neglect generally requires relatively large lesions to be evident. It usually lasts for only a few weeks or months, and the same patient tested on different days may have quite different profiles (note the dates and performance in Fig. 8.1 of a patient tested in a standard line bisection task). Even patients tested within the same session can show such variability (Marshall & Halligan, 1989). Chronic cases are relatively rare. Neglect typically resolves into double simultaneous extinction where items on the contralateral side to the lesion are ignored only when two items are presented simultaneously side by side. Neglect and extinction probably lie on a continuum with extinction being a less severe form of neglect (see Robertson, 1992). However, there is some controversy over this claim because neglect can be found without extinction. However, the tests for the two have not been equated in any way when defining these conditions clinically, so it is unclear what this dissociation means. It is also important to keep in mind that some explanations of neglect are based on patients with extinction and others on patients with neglect. Some are even based on patients who have no clinical evidence of neglect or extinction at the time of testing, but do have lesions in cortical areas often associated with neglect (most prominently the parietal lobe; but see Heilman, Watson, & Valenstein, 1985; Mesulam, 1985; Rizolatti & Camarda, 1987).
Fig. 8.1. Changes in line bisection over a two week period as tested at bedside.
Despite the difficulties in standardising neglect and testing patients with it, a very interesting picture is beginning to emerge from recent studies largely motivated by theories derived from cognitive psychology. Much of the focus has been on the role of attention vs spatial representation in visual spatial neglect. Several investigators argue that neglect is due to an attentional deficit (although the argument comes in different forms; Eglin, Robertson, & Knight, 1989; Farah, 1990; Heilman et al., 1985; Kinsbourne, 1987; Ladavas, this volume; Reuter-Lorenz, Kinsbourne, & Moscovitch, 1990; Robertson, 1992), while others argue for a deficit in spatial representation (Bisiach & Berti, 1987; De Renzi, 1982; Morrow, 1987). According to the attentional theories, attentional mechanisms are affected directly. According to theories of altered spatial representation, the observed attentional deficits are affected indirectly. In both cases, the overt manifestation of the deficit will be altered visual search.
Attention and Visual Neglect
When relying on vision, all animals need some kind of mechanism to tell them where to look next. A system where the eyes would move automatically to whatever is most (or perhaps least) salient in the peripheral visual field would be rather maladaptive. For instance, without control over spatial attention, it would be hopeless to try to keep one’s eyes on this sentence until it is finished before moving them to the person who just entered the room. Some mechanism is required that monitors areas of the visual field for potentially important information and other mechanisms to determine when and where the eyes move next or even if they move at all.
The most thorough investigations of the elementary operations of such a system have been reported by Posner and his colleagues (Posner & Petersen, 1990; Posner, Walker, Friedrich, & Rafal, 1984; Posner, Inhoff, Freidrich, & Cohen, 1987a; Posner, Walker, Friedrich, & Rafal, 1987b; Rafal et al., 1988; Rafal & Posner, 1987). In a simple visual task, they were able to break down covert spatial attention into move, engage and disengage operations, each associated with different neural structures in an overall network that guides visual spatial attention (Posner & Cohen, 1984). For the present purpose, the disengage operation is the most important because it is said to be altered in patients with neglect due to middle cerebral artery infarct (Morrow & Ratcliff, 1987), and in patients with extinction and/or more focal parietal lobe damage (Posner et al., 1984, 1987; Rafal, pers. comm.). The basic procedure is as follows. Three boxes appear horizontally across the midline on a computer screen, and the subjects fixate on the central box. A trial begins with one of the two peripheral boxes brightening for a short period of time. This is the cue. After a variable interval (usually 50–1000 msec), an asterisk appears in either peripheral box. The subjects’ task is to keep their eyes fixated on the central box and to press a button as soon as the asterisk is detected. Reaction time (RT) to the target in the cued location becomes faster as the interval between cue and target lengthens with an increasing delay in RT when the target appears in the uncued location (invalid location RT minus valid location RT). This difference between valid and invalid RT theoretically reflects the time to disengage covert attention from the cued location, move and engage on the target in the uncued location. Patients with parietal lobe damage detect the asterisk nearly as well in the contralesional and ipsilesional sides of space when it occurs in the cued location (nearly equal ability to move and engage attention to cued locations). However, the delay in responding to the asterisk is increased substantially when it occurs in the uncued location in the neglected field. Patients with parietal lobe damage show an abnormal contralesional delay. When attention must be disengaged from a location on the intact side, there is abnormal delay, and, as with clinical neglect, the delay is worse for right hemisphere groups than for left. Therefore, it is possible that the visual search problems observed in patients with neglect stem from a difficulty in disengaging attention from the ipsilesional side of space once attention has been engaged on an object or location on that side.
These findings are consistent with electrophysiological evidence in monkey, showing that the increased response of many visual cells in parietal lobe (area 7) only occur when the stimulus is relevant to the animal (e.g. predicts food; Bushnell, Goldberg, & Robinson, 1981; Lynch, 1980; Mountcastle, 1978). During free visual search, the cells do not increase firing rate, nor do they do so when the monkey is fixated and the stimulus is not predictive. These data led investigators to conclude that a subset of cells in area 7 are responsible for spatial attention to the object. Posner and co-workers’ data suggest a refinement of this conclusion. Rather than the parietal lobe being more involved in spatial attention as a whole, it appears to be involved in the command to disengage attention from its current location. It would make sense that this command would only be sent if a peripheral stimulus were potentially important and would not be sent if attention were not fixated because no disengagement would be required. Although animal studies have not pursued the disengage operation, Posner and co-workers’ data suggest that findings from humans could be fruitfully related to investigations into more specific elementary operations in animals.
Recent findings using a very different task (Eglin et al., 1989) are also consistent with the idea that attentional disturbances in patients with neglect reflect a disengage problem in a task that requires visual search. Using feature and conjunction search tasks developed by Treisman and her colleagues (Treisman & Gelade, 1980; Treisman & Gormican, 1988), we varied the side of a predesignated target among a variable number of distractors. Our findings show that patients with neglect have difficulty disengaging from stimulation on the entire half of space ipsilateral to the lesion, but again have no difficulty if target and distractors only appear on the right or left side of the page.
In feature and conjunction search tasks, a target is displayed among a varying number of distractors. The target location varies from trial to trial, and RT to detect its presence or absence is measured. When the target is defined by a unique visual feature (e.g. colour, orientation), the number of distractors make little difference. The target “pops out” from its background. Conversely, when the target is defined by a combination of features (a conjunction), RTs typically increase linearly with the number of distractors in the display. Feature integration theory was developed to accommodate such results (Treisman & Gelade, 1980). It proposes two stages of analysis: an initial stage during which features are detected and a second attentional stage during which conjunctions of features are identified. Feature and conjunction search tasks can be used to determine whether or not a deficit in disengagement of attention from an ipsilateral object is specific to the right or left side of a display and whether it only occurs when attention is involved. Riddoch and Humphreys (1987) addressed these questions in three patients with left neglect due to right hemisphere damage. These subjects were poor at detecting both feature and conjunction targets on the left side. However, error rates were so high in the neglected field that search rates calculated from RT were problematic.
We used a localisation task rather than a presence-absence judgement and timed subjects’ ability to point to a target that they knew would be present on every trial (Eglin et al., 1989). Under these conditions, even patients with relatively severe neglect continued to search into the neglected space because they knew that a target must be present. We tested seven patients who were included on a behavioural criterion of neglect: None of them crossed out any lines past the midline of a line cancellation test. RT was measured from stimulus presentation until a pointing response to the target was given. The target and distractor locations varied and were crossed with the right or left side of the display. The number of distractors varied orthogonally with side of display (see Fig. 8.2 for examples).
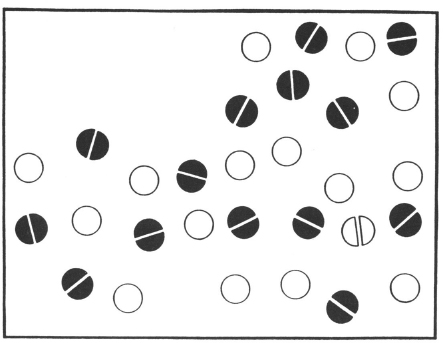
Fig. 8.2. Example of stimulus used in conjunction search task with target plus 19 distractors on the right side and 10 distractors on the left. Open circles represent red dots and filled circles represent blue dots. Reprinted with permission from Eglin, Robertson, & Knight (1989).
The most revealing findings were produced by the conjunction search task where serial search was required (see Fig. 8.3). The major findings are as follows:
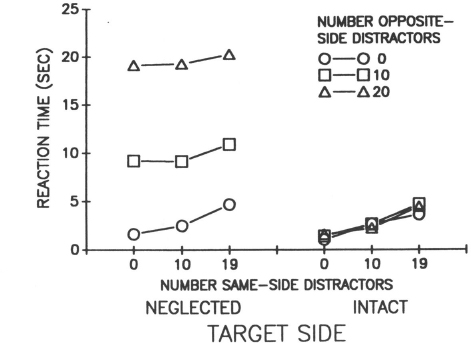
Fig. 8.3. Reaction time to locate a conjunction target on the intact and neglected side of a display as a function of the number of same-side and opposite-side distractors. Reprinted with permission from Eglin, Robertson, & Knight (1989).
The contralateral delay is represented in a different way in Fig. 8.4, where RT for each vertical quartile of the displays from the most intact to the most neglected quartile sections is plotted collapsed over all other conditions. There was a gradual increase from the most intact to the most neglected side (see also Posner, Inhoff, Freidrich, & Cohen, 1987b), but there was also an abrupt change in the function in the centre. Crossing the midpoint posed a special problem for these patients, but only when stimuli occurred on both sides of the display. In a follow-up study with stable patients with unilateral posterior or anterior lesions without clinical signs of neglect, we did not find this midline problem. However, search still began at the most ipsilateral point to the side of the lesion with RT increasing gradually as the target appeared more towards the neglected side (Eglin, Robertson, & Knight, 1991). For patients with severe neglect, attention was not distributed over space like a single gradient smoothly crossing the midline. The midline of the body defines a spatial frame that demarcates left and right in viewer-centred co-ordinates. The dominance of viewer-centred co-ordinates dissipates over time in patients with neglect, and may be part of the anosognosia that so often co-occurs with syndrome.
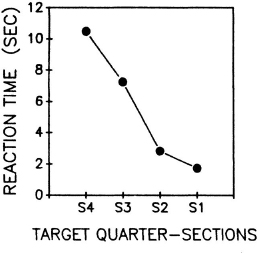
Fig. 8.4. Reaction time to locate a conjunction target in the four vertical quartiles of the page collapsed over other conditions. Reprinted with permission from Eglin, Robertson, & Knight (1989).
It is intriguing that Luck, Hillyard, Mangun and Gazzaniga (1989) have reported evidence in patients with full commissurotomy which suggests that each hemisphere independently guides search on the contralateral side of space. This may account for the fact that distractors on the neglected side have no effect until attention crosses into the contralesional side in patients with neglect.
In sum, these findings together suggest at least three different contributions to attentional search deficits in patients with unilateral visual neglect. The first is a directional scanning bias starting at the most ipsilesional stimulus on the intact side. The second is a slowed attentional search rate regardless of field (perhaps a slowed disengage problem from point to point everywhere in the visual field). This could contribute to neglect by increasing the probability of returning to parts of the display that already had been examined. Possibly, inhibitory tagging could be disrupted (Klein, 1988). The third is a difficulty in disengaging from the intact half of the stimulus display to move into the neglected half.
Our findings have been replicated with other displays that do not rely on colour or even on conjoining features (Eglin, Robertson, Knight, & Brugger, submitted). Examples of such stimuli are shown in Fig. 8.5, where the Q pops out from the Os, but to find an O in a field of Qs requires a serial search. The absence of a feature in the target requires a serial search, while the presence of a feature does not (Treisman & Gormican, 1988; Treisman & Souther, 1985). Finally, the contralateral delay can be reduced if the stimuli are grouped by one of the features defining the target (e.g. by colour or by shape) because attention can be allocated to a homogeneous group of items as a whole. Even grouping on the contralateral side can affect the magnitude of neglect (Grabowecky, Robertson, & Treisman, in press) even though a heterogeneous field has no effect on search whatsoever. These findings together show that the number of individual items on the ipsilesional side in a serial search task does not suffice to produce the contralateral delay. Rather, only those items or groups of items that are serially scanned contribute to the delay in patients with neglect (Eglin et al., submitted). The latter result also suggests intact grouping processes in patients with neglect.

Fig. 8.5. Example of stimuli. The top display produces functions indicative of serial search and the bottom does not.
Attention, Spatial Representation and Neglect
There are clear indications from the foregoing discussion as well as clinical observation that spatial attention is altered in patients with unilateral visual neglect. This does not necessarily mean that attentional mechanisms are affected directly. Attention must be engaged on something (whether an object or a location), and if certain parts of space are weakly represented, then attention will be indirectly affected.
The most prominent competing theory to attentional accounts suggests just this. Bisiach and his colleagues (Bisiach & Berti, 1987; Bisiach, Capitani, Luzzatti, & Perani, 1981; Bisiach & Luzzatti, 1978; Bisiach, Luzzatti, & Perani, 1979) propose that representational space has been altered in unilateral neglect. Patients with left neglect will neglect the left side of a stimulus even when it is visually imagined (Bisiach & Luzzatti, 1978; Bisiach et al., 1981), or when a pattern is presented sequentially by drifting it behind a cut-out slit in central vision (Bisiach et al., 1979). These investigators also found that when patients did recall the neglected side of an imagined scene, they often misplaced the object into the intact side. There was “visual allaesthesia”. This observation is consistent with clinical evidence. When drawing in the clock numbers of a clock face, patients with neglect typically do not draw in 1 through 6 and then stop. They are much more likely to place 7 through 11 on or towards the intact side. As shown in Fig. 8.6., the left side seems compressed rather than absent. This does not appear only in drawings. Mijovic (1990) demonstrated that patients with neglect began to move to locations on a covered clock face in a direction that reflected altered spatial representation similar to the drawings. The subjects were first shown a clock face and then a cover with a small hole in the centre was placed on top of it. Thus, all the subjects saw during the test was a cover with a white square cut out. When asked to move the open square to a number on the clock face, initial movement was towards a position that reflected an altered spatial representation consistent with the pattern seen in patient drawings. For instance, when asked to move to 9 o’clock, the subjects might move in a direction towards 2 or 3. As in patient drawings, the left-sided numbers were displaced towards the right within the representation of the clock face. In other studies, Mijovic was able to rule out a directional motor movement bias.
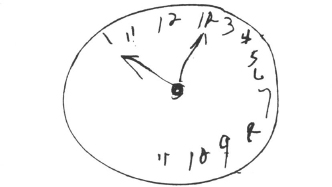
Fig. 8.6. Example of a patient’s drawing of clock numbers.
One problem with the representational account is that there are other cases in which patients will draw clock numbers towards the intact side with their eyes open but draw them appropriately when told to close their eyes (Mesulam, 1985). This suggests there may be some cases of neglect that are due to altered spatial representation and others that are not. Also, Mijovic’s representational account has difficulty explaining the lack of effect of contralesional distractors on searching for a target on the intact side in the study by Eglin et al. (1989). Visual allaesthesia should increase the number of distractors perceptually represented on the intact side. If we assume that the number of displaced items increases the more items are present on the neglected side, the slopes on the intact side should increase as the number of distractors on the neglected side increases. This would increase the slopes of the search functions on the intact side relative to the neglected side of space, which did not occur. Neither do the data from Eglin et al. account for the visual allaesthesia observed in clock drawings and the data from Mijovic’s study. Visual allaesthesia and contralateral delay likely reflect different components of the neglect syndrome.
Neglect and Spatial Transformations
If the major deficit in neglect is altered spatial representation, it is somewhat surprising that spatial transformations that would seem to require a rather high degree of abstract spatial representation can be made by patients with visual neglect. For instance, Driver and Halligan (1991), Farah et al. (1990) Làdavas (1987; 1990), Calvanio, Petrone and Levine (1987) and Posner et al. (1987) have shown that neglect or extinction are found both on one side of space defined by the body midline, the stimulus display, or by the orthogonal upright environment. When patients with right hemisphere damage and neglect are rotated 90°, viewer-centred and environmentally centred reference frames are orthogonal, yet neglect occurs within both frames. Driver and Halligan (1991) have also shown that neglect can be relative to object-centred frames as well. Same-different judgements for patterns rotated 45° showed neglect for the left side of the objects and did so even when presented entirely within the intact field.
Patients with unilateral left neglect are able to represent differently oriented frames of reference, but then they neglect the left side of each frame. In other words, spatial representations and the deficits within them are stable across orientation and location transformations. This stability does not preclude the possibility that spatial representation is altered consistently on one side across reference frame transformations, but it does make the explanations more complicated and demonstrates that patients with neglect can utilise abstract representations of space in similar ways as normals.
Consistently, there is evidence in normals that directional scanning biases are relative to the orientation of a reference frame. In a series of studies, Robertson and Lamb have shown that a rightward sampling bias to upright patterns continues as a rightward bias in disoriented patterns (Robertson & Lamb, 1988, 1989; Robertson, 1990). When a group of four letters was presented in the left or right visual field and subjects judged whether the letters were normal or reflected (see Fig. 8.7a), a right visual field advantage was observed.1 When the letters were rotated around the centre fixation point into the upper or lower visual field as in Fig. 8.7b, the advantage was still for the right field relative to the orientation of the letters. When the tops of the letters were +90° from upright as in Fig. 8.7b, RT advantage was in the lower visual field, and when they were −90° from upright in the opposite direction as in Fig. 8.7c, RT advantage was in the upper visual field. In both cases, the advantage was for patterns in the relative right field in a reference frame rotated through fixation to correspond to the orientation of the letters (see Fig. 8.8). This was the case whether the patterns were normal or reflected ruling out sensory interpretations. As in patients with neglect, scanning biases were consistent across spatial transformation and were rightward in every case.
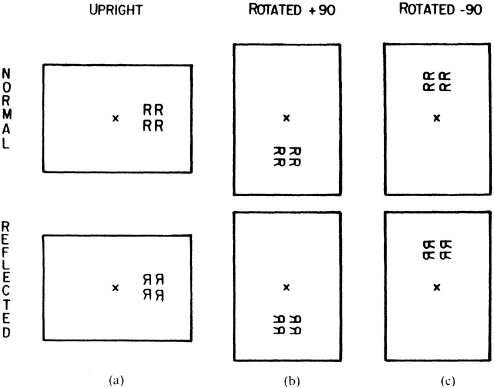
Fig. 8.7. Example of stimuli used in studies of reference frame effects. All patterns are shown in the right relative field. Reprinted with permission from Robertson & Lamb (1988).
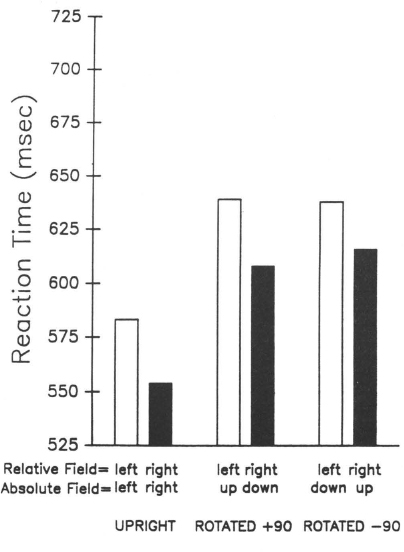
Fig. 8.8. Reaction time as a function of display field (both relative and absolute) for the three orientation conditions. Reprinted with permission from Robertson & Lamb (1988).
Other investigators using very different paradigms have also found a rightward advantage in normals (Efron, 1990; Egly & Homa, 1984; Reuter-Lorenz et al., 1990; Werth & Poppel, 1988). Efron and his colleagues (Efron, Yund, & Nichols, 1987, 1990a, 1990b; Yund, Efron, & Nichols, 1990a, 1990b) have repeatedly reported a rightward advantage when subjects must detect a non-linguistic target among a briefly presented display of other non-linguistic targets. Overall target detection was better in the right upper quadrant. This occurred whether the subjects were literate or illiterate, demonstrating the independence of a rightward sampling bias from language processes (Efron, 1990). Obviously, not all tasks will produce a rightward bias in normals. As many visual field studies have shown, the rightward bias can be changed by stimulus and/or task demands (Hellige, 1983), but all else being equal, normals are more likely to sample the right side of space before the left. A recent set of studies reported by Reuter-Lorenz et al. (1990) showed that when a left and right bias were pitted against each other, a rightward bias consistently prevailed.
The rightward biases in normals in upright conditions are consistent with a model of hemisphere laterality and attentional allocation developed by Kinsbourne (1975). His model was designed to account for rightward biases in both normals and patients with left neglect (Kinsbourne, 1977; 1987; see also Chapter 3, this volume). According to his model, attention is directed to locations through a balance in opponent activation between the two halves of the brain (see Kinsbourne, 1987). All else being equal, the left hemisphere is more highly activated than the right hemisphere, causing an overall bias towards the right side of space. In addition, there is mutual inhibition between the hemispheres (actually between tectal regions influenced by hemispheric activation) and when right hemisphere damage occurs, the normal inhibitory effect of the stronger left hemisphere bias is disrupted and causes more severe neglect than with left hemisphere damage.
In its present form, this model can account for the rightward bias in many studies in normals and in the upright condition of the Robertson and Lamb experiments, but the model has difficulty in accounting for the rightward bias when patterns are rotated around the picture plane or when subjects with neglect are rotated 90°. However, it is possible that the two hemispheres represent the two halves of space the same, whether the stimuli are spatially defined by sensory channels or transformed via internal mechanisms. This account is consistent with much of the work in imagery and suggests that the subjects may have rotated the letters in the Robertson and Lamb studies around fixation into a viewer-centred frame with the patterns then being channelled through the right or left hemispheres as if they were upright. This issue is critical because Kinsbourne’s idea that the vector of attention is determined by opponent processes between the two hemispheres could be extended to imagery tasks if we could show that an image of the pattern was rotated to upright before attentional scanning began. Under such conditions, the image in its imagined space would be analogous to a pattern presented in either the left or right visual field. However, image rotation received little support in studies we conducted to examine this issue (Robertson, 1990).
In one experiment, subjects judged the reflection of rotated letters as in Fig. 8.9. In this case, the patterns were always presented in the left or right visual field. If the subjects rotated letters around fixation to perform the task, then the image of the patterns would be represented in the upper or lower visual field. According to a hemispheric account, no asymmetry should be found. Yet when letters were locally rotated +90°, a right visual field advantage was found, and when they were rotated -90°, a left visual field advantage was found (Robertson & Lamb, 1989). The fact that rotated letters produced longer RTs than upright letters was also consistent with a mental rotation of some kind. There were four possible rotations that subjects may have used: local rotation of an image, local rotation of a reference frame, global rotation of an image or global rotation of a reference frame. However, in any of these cases, the findings were inconsistent with Kinsbourne’s opponent hemisphere activation model.
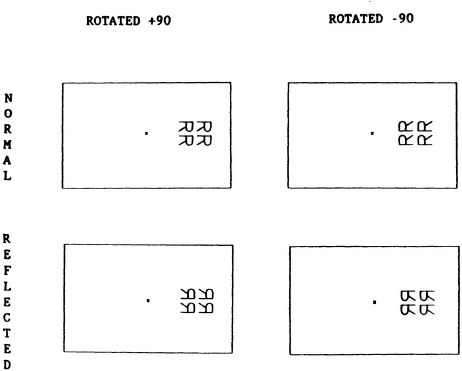
Fig. 8.9. Example of stimuli with a local rotation within the left or right visual field. From Robertson & Lamb (1989).
A local rotation around the centre of the four-lettered pattern would produce an image of the pattern in the field of presentation. In this case, the advantage should not vary as a function of orientation because, whether the letters were oriented plus or minus 90°, a local rotation would result in image representation in the same visual field as the pattern was presented. Although the magnitude of the field advantage was not the same for + 90° and −90°, the direction was clearly opposite, as shown in Fig. 8.10. The same logic holds for local rotation of a reference frame. On the other hand, a global rotation of an image around fixation would result in image representation in the upper or lower visual field. Although a field
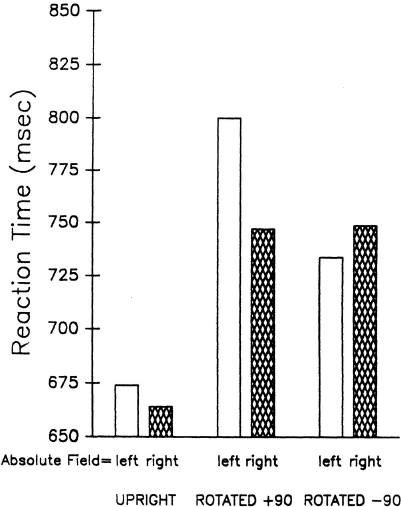
Fig. 8.10. Reaction time for left and right visual field over the three orientation conditions. From Robertson & Lamb (1991).
advantage was found in this experiment commensurate with an upper relative field advantage, a hemispheric explanation is not tenable, since the representation would now be along the up/down axis of the head through the midline. Finally, an upright reference frame may have been rotated into the orientation of the letters. Again, the field advantage in this case would be for the relative upper field within represented space, the difference being that the fields are defined within a rotated space from upright rather than viewer-centred as with image rotation. In its present form, the opponent activation model of differential hemisphere activation cannot account for the consistent field advantage over transformations within this type of rotation either. With a frame rotation, the “image” would not be moved from the right or left visual field, yet the visual field advantage changed to favour the upper relative field.
These results may be relevant for attentional and/or representational explanations of neglect, even though at first glance they do not seem to fit well with the functional neuroanatomy proposed to underly attentional systems and neglect (e.g. Heilman et al., 1985; Kinsbourne, 1975, 1977, 1987; Mesulam, 1985). There are cases of altitudinal neglect that have been reported consistent with a directional bias along the vertical axis (Butter, Evans, Kirsch, & Kewman, 1989; Rapcsak, Cimino, & Heilman, 1988), and patients with neglect do show directional biases across spatial transformations. Thus, the data we have collected could reflect a right hemisphere dominance in allocating attention along the left/right dimension and up/ down dimension. The issue of frame vs image rotation then becomes critical in interpreting the effects.
In order to distinguish between the two, we used an orientation priming task similar to the one used by Robertson, Palmer and Gomez (1987) to study frame effects in mental rotation. In this task, a group of letters was oriented plus or minus 90°, and the subject responded whether the letters were normal or reflected as if they were upright. We will call this pattern the prime. A second group of letters then appeared (the target), again either normal or reflected, and the subject made a second reflection judgement. By the pattern of RT to the target, it could be determined whether frame or image rotation occurred when judging the reflection of the prime. To study hemispheric effects, misoriented primes were presented in the lower or upper visual field followed by an upright target in the left or right visual field (see Fig. 8.11a). If the subjects rotated the prime to upright around central fixation to make their first reflection judgement, then the image representation would be as shown in Fig. 8.11b. This should produce faster RTs for a subsequent target presented in a consistent location (Fig. 8.11c) where the image was expected to be compared to an inconsistent location (Fig. 8.11d). If a frame rotation occurred in order to judge the reflection of the prime, then a subsequent upright pattern should not benefit. The data supported frame rotation rather than image rotation. An overall right visual field advantage occurred for the upright target consistent with the previous results, but the prime did not change RT to the target in accordance with image interpretations. A rotation of an image into the right visual field would have facilitated performance for a subsequent pattern presented in the right visual field, and a rotation of an image into the left visual field would have facilitated performance for a subsequent pattern presented in the left visual field. This did not occur. It appears instead that subjects rotated a frame to make their judgements. In other words, together the data suggest that attention is allocated relative to the frame of reference used to analyse the display. We have not yet tested patients with neglect using this procedure, but we do know that neglect can be found within different frames of reference (Calvanio et al., 1987; Driver & Halligan, 1991; Farah et al., 1990; Grabowecky et al., in press). The foregoing discussion suggests that the rightward bias found in patients with left neglect may be an exaggerated form of the rightward bias found in normals. It also suggests that mechanisms that select and transform reference frames are intact in these patients with attention being allocated rightward within the frame. It is difficult to know how an opponent process between the two hemispheres could account for an upper or lower visual field bias that is relative to the frame, although Kinsbourne’s idea of a vector of attention that is rightward biased has been well supported by the evidence. However, the rightward bias is not relative to the decussation of the visual pathways. Rather, it is relative to the frame of reference that defines the field.
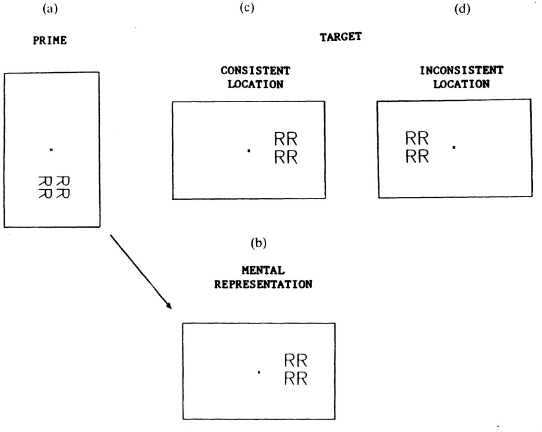
Fig. 8.11. Example of prime (a) and target (c or d) location and orientation, (b) shows the expected mental representation if a global rotation of an image occurred when responding to the prime.
Conclusion
It seems fairly clear that the directional scanning biases observed in patients with unilateral visual neglect are due to attentional mechanisms that are either directly or indirectly affected by neural damage. Visual neglect is not limited to the hemisphere of visual input. It has been found relative to viewer-centred, environment-centred and object-centred co-ordinates. It is not information within a visual field but information within the stimulus display in representational space that is neglected. As discussed in the first section, attention can be moved into neglected space as easily as into intact space when the intact field is devoid of task-relevant objects.
As discussed in the second section, the evidence from normals and patient populations together suggests a continuum of attentional bias towards the right of representational space. When all else is equal, information to the right of an adopted reference frame is sampled first. When right hemisphere damage occurs, this tendency is magnified. The case of neglect may be the end-point where everything on one side of the frame is ignored. Because this occurs over spatial transformations, it suggests a link between attentional biases and the reference frame one uses to analyse the scene. It also seems important that various attentional manipulations that can change the basic rightward bias in normals can increase or decrease the amount of neglect observed as well. For instance, normals can be cued to sample the left side of space first as can patients with neglect (Riddoch & Humphreys, 1983; 1987).
Notice that this account of neglect does not solve the issue of whether neglect is due to a direct effect on an attentional mechanism or to altered spatial representation. It is possible that, all else being equal, the right side of space (whether viewer-centred, object-centred or environment-centred) is more strongly represented than the left even in normals, and that it is this inequality that is magnified in patients with unilateral neglect.
Finally, it seems to us that an equally plausible account of an indirect effect on attention as altered spatial representation would be that some lateralised functions attract and/or sustain attention more than others in normals and in patients. With brain injury to a given hemisphere, certain types of information would be missing or weakly represented. For instance, patterns with global forms created from several local forms are responded to in opposite order in patient groups with left or right temporal-parietal (T-P) lesions. Patients with right T-P lesions favour local forms over global, while those with left T-P lesions favour global forms over local (Lamb, Robertson, & Knight, 1990; Robertson & Lamb, 1991; Robertson, Lamb, & Knight, 1988). This occurs whether the pattern is presented in the contralateral or ipsilateral field. Since local elements are typically more frequent than global ones in any visual scene, numerous local elements may be scanned on the ipsilateral side (utilising attention) and produce more severe signs of neglect in patients with right posterior damage, whereas for patients with left posterior damage and a global bias, relatively few global elements will compete for attention. The contralesional deficit would therefore be more severe with right than with left brain damage for hierarchically organised visual scenes. Although this idea is speculative, it does illustrate the fact that there are other possible indirect contributions to attention that could be pursued in addition to a spatial representation one. The current controversy need not be one of attention vs spatial representation, but rather the controversy may best be framed as direct effects vs indirect effects on attention and indeed even as direct combined with indirect effects that may produce different profiles of neglect. Whatever the case, the role of spatial transformations will have to play a large part in any theory of either normal or defective scanning biases.
Acknowledgements
Preparation of this manuscript was partially supported by VA Medical Research funds and by ADAMHA Grant No. AA06637 to L.C.R.
References
Battersby, W.S., Bender, M.B., Pollock, M., & Kahn, R.L. (1956). Unilateral “spatial agnosia” (inattention). Brain, 79, 68–93.
Bisiach, E. & Berti, A. (1987). Dyschiria: An attempt at its systemic explanation. In M. Jeannerod (Ed.), Neurophysiological and neuropsychological aspects of spatial neglect. Amsterdam: Elsevier.
Bisiach, E., Capitani, E., Luzzatti, C., & Perani, D. (1981). Brain and conscious representation of outside reality. Neuropsychologia, 19, 543–551.
Bisiach, E. & Luzzatti, C. (1978). Unilateral neglect of representational space. Cortex, 14, 129–133.
Bisiach, E., Luzzatti, C., & Perani, D. (1979). Unilateral neglect, representational schema and consciousness. Brain, 102, 609–618.
Bushnell, M.C., Goldberg, M.E., & Robinson, D.L. (1981). Behavioral enhancement of visual responses in monkey cerebral cortex I. Modulation in posterior parietal cortex related to selective visual attention. Journal of Neurophysiology, 46, 755–772.
Butter, C.M., Evans, J., Kirsch, N., & Kewman, D. (1989). Altitudinal neglect following traumatic brain injury: A case report. Cortex, 25, 135–146.
Calvanio, R., Petrone, P.M., & Levine, D.N. (1987). Left visual spatial neglect is both environmental-centered and body-centered. Neurology, 37, 1179–1183.
De Renzi, E. (1982). Disorders of space exploration and cognition. New York: John Wiley.
Driver, J. & Halligan, P.W. (1991). Can visual neglect operate in object-centered coordinates? An affirmative single-case study. Cognitive Neuropsychology, 8, 475–496.
Efron, R. (1990). Decline and fall of hemispheric specialization. Hillsdale, NJ: Lawrence Erlbaum Associates Inc.
Efron, R., Yund, E.W., & Nichols, D.R. (1987). Scanning the visual field without eye movements: A sex difference. Neuropsychologia, 25, 637–644.
Efron, R., Yund, E.W., & Nichols, D.R. (1990a). Detectability as a function of target location: Effects of spatial configuration. Brain and Cognition, 12, 102–116.
Efron, R., Yund, E.W., & Nichols, D.R. (1990b). Visual detectability gradients: The effect of distractors in contralateral field. Brain and Cognition, 12, 128–143.
Eglin, M., Robertson, L.C., & Knight, R.T. (1989). Visual search performance in the neglect syndrome. Journal of Cognitive Neuroscience, 1, 372–385.
Eglin, M., Robertson, L.C., & Knight, R.T. (1991). Cortical substrates supporting visual search in humans. Cerebral Cortex, 1, 262–272.
Eglin, M., Robertson, L.C., Knight, R.T., & Brugger, T. (submitted). Search for simple visual features in patients with visual neglect.
Egly, R. & Homa. D. (1984). Sensitization in the visual field. Journal of Experimental Psychology, 10, 778–793.
Farah, M J. (1990). Visual agnosia: Disorders of object recognition and what they tell us about normal vision. New York: Academic Press.
Farah, M.J., Brunn, J.L., Wong, A.B., Wallace, M.A., & Carpenter, P.A. (1990). Frames of reference for allocating attention to space: Evidence from the neglect syndrome. Neuropsychologia, 28, 335–347.
Grabowecky, M., Robertson, L.C., & Treisman, A. (in press). Preattentive processes guide visual search: Evidence from patients with unilateral visual neglect. Journal of Cognitive Neuroscience.
Heilman, K.M., Watson, R.T., & Valenstein, E. (1985). Neglect and related disorders. In K.M. Heilman & E. Valenstein (Eds), Clinical neuropsychology. New York: Oxford University Press.
Hellige, J.B. (1983). Hemisphere x task interaction and the study of laterally. In J.B. Hellige (Ed.), Cerebral hemisphere asymmetry: Method, theory and application. New York: Praeger.
Kinsbourne, M. (1975). The mechanisms of hemispheric control of the lateral gradient of attention. In P.M.A. Rabbitt & S. Dornic (Eds), Attention and performance v. London: Academic Press.
Kinsbourne, M. (1977). Hemi-neglect and hemispheric rivalry. In E.A. Weinstein & R.P. Friedland (Eds), Hemi-inattention and hemispheric specialization. New York: Raven Press.
Kinsbourne, M. (1987). Mechanisms of unilateral neglect. In M. Jeannerod (Ed.), Neurophysiological and neuropsychological aspects of spatial neglect. Amsterdam: Elsevier.
Klein, R. (1988). Inhibitory tagging system facilitates visual search. Nature, 334, 430–431.
Làdavas, E. (1987). Is the hemispatial deficit produced by right parietal lobe damage associated with retinal or gravitational coordinates? Brain, 110, 167–180.
Làdavas, E. (1990). Selective spatial attention in patients with visual extinction. Brain, 113, 1527–1530.
Làdavas, E. (1993, this volume). Spatial dimensions of automatic and voluntary orienting components of attention. In I.H. Robertson & J.C. Marshall (Eds), Unilateral neglect: Clinical and experimental studies. Hove: Lawrence Erlbaum Associates Ltd.
Lamb, M.R., Robertson, L.C., & Knight, R.T. (1990). Component mechanisms underlying the processing of hierachically organized patterns; Inferences from patients with unilateral cortical lesions. Journal of Experimental Psychology: Memory, Learning and Cognition, 16, 471–483.
Luck, S.J., Hillyard, S.A., Mangun, G.R., & Gazzaniga, M.S. (1989). Independent hemispheric attentional systems mediate visual search in split-brain patients. Nature, 342, 543–545.
Lynch, J.C. (1980). The functional organization of posterior parietal association cortex. Behavior and Brain Science, 3, 485–534.
Marshall, J.C. & Halligan, P.W. (1989). When right goes left: An investigation of line bisection in a case of visual neglect. Cortex, 25, 503–515.
Mesulam, M.M. (1981). A cortical network for directed attention and unilateral neglect. Annals of Neurology. 10, 309–325.
Mesulam, M.M. (1983). The functional anatomy and hemispheric specialization for directed attention: The role of the parietal lobe and its connectivity. Trends in Neurosciences, 6, 384— 387.
Mesulam, M.M. (1985). Attention, confusional states and neglect. In M.M. Mesulam (Ed.), Principles of behavioral neurology. Philadelphia, PA: F.A. Davis.
Mijovic, D. (1990). The mechanisms of visual neglect. Unpublished doctoral dissertation, Boston University.
Morrow, L.A. (1987). Cerebral lesions and internal spatial representation. In P. Ellen & C. Thinus-Blanc (Eds), Cognitive processes and spatial orientation in animal and man. Vol. II. Boston, MA: Martinus Nijhoff.
Morrow, L.A. & Ratcliff, G. (1987). Attentional mechanisms in clinical neglect. Journal of Clinical and Experimental Neuropsychology, 9, 74–75.
Mountcastle, V.B. (1978). Brain mechanisms for directed attention. Journal of the Royal Society of Medicine, 71, 14–28.
Ogden, J.A. (1987). The “neglected” left hemisphere and its contribution to visuospatial neglect. In M. Jeannerod (Ed.), Neurophysiological and neuropsychological aspects of spatial neglect. Amsterdam: Elsevier.
Posner, M.I. & Cohen, Y. (1984). Components of visual orienting. In H. Bouma & D. Bowhais (Eds), Attention and performance X. Hillsdale, NJ: Lawrence Erlbaum Associates Inc.
Posner, M.I., Inhoff, A.W., Friedrich, F.J., & Cohen, A. (1987a). Isolating attentional systems: A cognitive-anatomical analysis. Psychobiology, 15, 107–121.
Posner, M.I. & Petersen, S.E. (1990). The attention system in the human brain. Annual Review of Neuroscience, 13, 25–42.
Posner, M.I., Walker, J.A., Friedrich, F.A. & Rafal, R.D. (1984). Effects of parietal injury on covert orienting of attention. Journal of Neuroscience, 4, 1863–1874.
Posner, M.I., Walker, J.A., Friedrich, F.A., & Rafal, R.D. (1987b). How do the parietal lobes direct covert attention? Neuropsychologia, 25, 235–245.
Rafal, R.D. & Posner, M.I. (1987). Deficits in human visual spatial attention following thalamic lesions. Proceedings of the National Academy of Sciences, 84, 7349–7353.
Rafal, R.D., Posner, M.I., Friedman, J.H., Inhoff, A.W., & Bernstein, E. (1988). Orienting of visual attention in progressive supranuclear palsy. Brain, 111, 267–280.
Rapcsak, S.Z., Cimino, C.R., & Heilman, K.M. (1988). Altitudinal neglect. Neurology, 38, 277–281.
Reuter-Lorenz, P.A., Kinsbourne, M., & Moscovitch, M. (1990). Hemispheric control of spatial attention. Brain and Cognition, 12. 240–266.
Riddoch, M.J. & Humphreys, G.W. (1983). The effect of cueing on unilateral neglect. Neuropsychologia, 21, 589–599.
Riddoch, M.J. & Humphreys, G.W. (1987). Perceptual and action systems in unilateral visual neglect. In M. Jeannerod (Ed.), Neurophysiological and neuropsychological aspects of spatial neglect. Amsterdam: Elsevier.
Rizzolatti, G. & Camarda, R. (1987). Neural circuits for spatial attention and unilateral neglect. In M. Jeannerod (Ed), Neurophysiological and neuropsychological aspects of spatial neglect. Amsterdam: Elsevier.
Robertson, L.C. (1990). Visual field asymmetries in analyzing rotated letters. Paper presented at the International Neuropsychoiogical Society Meeting, Innsbruck, Austria (July).
Robertson, L.C. (1992). The role of perceptual organization and search in attentional disorders. In D.I. Margolin (Ed.), Cognitive neuropsychology in clinical practice. New York: Oxford University Press.
Robertson, L.C. & Lamb, M.R. (1988). The role of perceptual reference frames in visual field asymmetries. Neuropsychologia, 26, 145–152.
Robertson, L.C. & Lamb, M.R. (1989). Judging the reflection of misoriented patterns in the right and left visual fields. Neuropsychologia. 27, 1081–1089.
Robertson, L.C. & Lamb, M.R. (1991). Neuropsychoiogical contributions to part-whole organization. Cognitive Psychology, 23, 299–330.
Robertson, L.C, Lamb, M.R., & Knight, R.T. (1988). Effects of lesions of temporal-parietal junction on perceptual and attentional processing in humans. Journal of Neuroscience, 8, 3757–3769.
Robertson, L.C., Palmer, S.E., & Gomez, L.M. (1987). Reference frames in mental rotation. Journal of Experimental Psychology: Learning, Memory and Cognition, 13, 368–379.
Roy, E.A., Reuter-Lorenz, P., Roy, L.G., Copland, S., & Moscovitch, M. (1987). Unilateral attention deficits and hemisphere asymmetries in the control of attention. In M. Jeannerod (Ed.), Neurophysiological and neuropsychological aspects of neglect. Amsterdam: Elsevier.
Treisman, A. & Gelade, G. (1980). A feature-integration theory of attention. Cognitive Psychology, 12, 97–136.
Treisman, A. & Gormican, S. (1988). Feature analysis in early vision: Evidence from search asymmetries. Psychological Review, 95, 15–48.
Treisman, A. & Souther, J. (1985). Search asymmetry: A diagnostic for preattentive processing of separable features. Journal of Experimental Psychology: General, 114, 285–310.
Werth, R. & Poppel, E. (1988). Compression and lateral shift of mental coordinate systems in a line bisection task. Neuropsychologia, 26, 741–745.
Yund, E.W., Efron, R., & Nichols, D.R. (1990a). Detectability gradients as a function of target location. Brain and Cognition, 12, 1–16.
Yund, E.W., Efron, R., & Nichols, D.R. (1990b). Target detection in one visual field in the presence or absence of stimuli in the contralateral field by right and left handed subjects. Brain and Cognition, 12, 117–127.
Zarit, S.H. & Kahn, R.L. (1974). Impairment and adaptation in chronic disabilities: Spatial inattention. Journal of Nervous and Mental Disease, 159, 63–72.
1 The stimulus configuration and task were chosen because a previous study found frame effects with these patterns in a study of frame rotation in mental rotation tasks (Robertson et al., 1987). Obtaining a significant right visual field advantage using letters in a reflection task is by no means guaranteed even in the upright condition. In seven different experiments using such stimuli, a mean right visual field advantage was found on every occasion (8–29 msec), but the advantage only reached significant levels on four of those occasions (with an N between 12 and 16). Obviously, asymmetries and their relation to frames can only be tested if a tield advantage is first observed in the upright condition. It should also be noted that the study of frame effects is not limited to right visual field advantage. In order to make a frame interpretation, it is sufficient to have a consistent field advantage (either right or left) over spatial transformations.