3 Neuroanatomy of the Ferret Brain with Focus on the Cerebral Cortex
Christopher D. Kroenke, Brian D. Mills, Jaime F. Olavarria, and Jeffrey J. Neil
Introduction
Ferrets (Mustela putorius furo) are widely used in developmental and system neuroscience studies because they offer a combination of useful characteristics [1]. In addition to having relatively large litters, the ferret gestational term is short relative to the rate of central nervous system development, such that at birth, the brain is at a similar developmental stage to mid-gestational brains of primate species. This permits experimental manipulations that would require in utero interventions in other species. Moreover, although ferrets are relatively small in body size, their brain presents many of the characteristics observed in phylogenetically advanced vertebrates such as humans and several nonhuman primates. For instance, the cerebral cortex of adult ferrets is folded into several gyri and sulci, which makes them an attractive model for studying factors influencing the development and organization of the cortical mantle.
The gross anatomy of the ferret brain has been described in detail previously [1–3]. Since the publication of this earlier work, significant progress in characterizing the development and organization of the ferret cerebral cortex has been made. Therefore, emphasis is placed here on reviewing this recent work.
Magnetic resonance imaging (MRI) is becoming an increasingly useful tool for neuroanatomical investigations due to its ability to characterize the three-dimensional structure of the brain, and it has recently been utilized in studies of ferrets [4–7]. An advantage of this technique is that it can noninvasively and nondestructively provide images that enable segmentation and three-dimensional surface rendering of the brain to reveal structures and their relationships to one another. Herein we utilize T2-weighted and diffusion tensor imaging (DTI) MRI data acquired from an adult female postmortem brain to illustrate the three-dimensional organization of cerebral cortex relative to subcortical structures. The brain was prepared by intracardial perfusion fixation with 4% paraformaldehyde as described previously [5] and transferred to phosphate-buffered saline (PBS) following 24 hours of postfixation. Both MRI data sets were acquired with the brain immersed in PBS using a small-animal 11.7-T MRI system (Bruker Biospin, Rheinstetten, Germany) interfaced with a 9-cm inner diameter magnetic field gradient coil and a 2.5-cm inner diameter, single-turn solenoidal transmit/receive radiofrequency coil. The T2-weighted image resolution consisted of 0.15-mm-sided cubic voxels, obtained from a multi-slice spin-echo pulse sequence in which the echo time was 22 ms and the recycle delay was 8 seconds. The DTI data were acquired with image resolution of 0.3-mm-sided voxels and diffusion sensitization settings consisting of 2 b = 0 and 25 b = 2.5 ms/µm2 acquisitions (27 total) as previously described [8]. The fields of view for both images were sufficiently large to cover the entire brain. The data represented in Fig. 3.5, as well as the T2-weighted image, have been made publicly available through the SumsDB web site (http://sumsdb.wustl.edu:8081/sums/index.jsp).
Overview
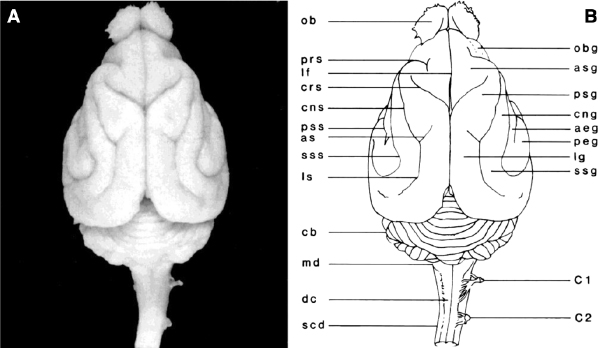
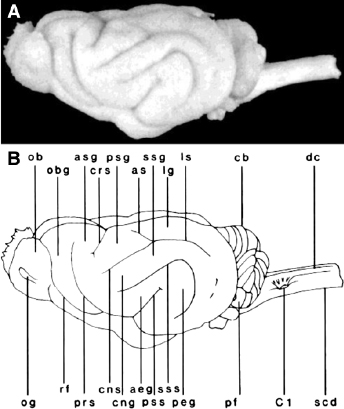
The gyral/sulcal cortical folding pattern observed in the ferret is highly conserved between animals, though some common minor variant patterns have been identified with respect to the caudal part of the lateral sulcus [9]. Overall, the pattern of sulci and gyri in the ferret brain resembles that of other carnivores (Fig. 3.1, Fig. 3.2, Fig. 3.3, and Fig. 3.4), with particular similarity being noted in cortical folding and functional organization between ferrets and cats [10,11]. The caudal half of the cerebral cortex contains a series of primarily rostral-to-caudal-oriented sulci and gyri. The lateral gyrus is the most dorsal and medial of these structures (Fig. 3.1 and Fig. 3.3). Extending laterally, the lateral sulcus separates the lateral and suprasylvian gyri. These gyri contain somatosensory areas and visual areas with proposed homology to the parietal processing stream in primates [12]. The suprasylvian sulcus and ectosylvian gyrus are lateral to the suprasylvian gyrus. The ectosylvian gyrus is curved, with the anterior ectosylvian gyrus located rostral and medial to the posterior ectosylvian gyrus, with the pseudosylvian sulcus separating them. Several auditory cortical areas reside on the ectosylvian gyrus, in proximity to the pseudosylvian sulcus. This arrangement resembles that of primate auditory areas, which are proximal to the primate's sylvian fissure. The rhinal fissure, located lateral and ventral to the ectosylvian gyrus (Fig. 3.3), separates isocortex from allocortex [3].
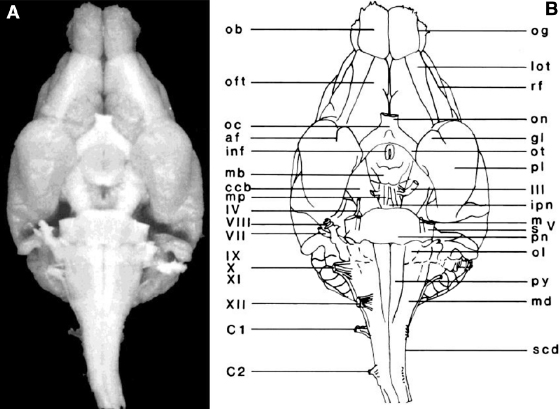
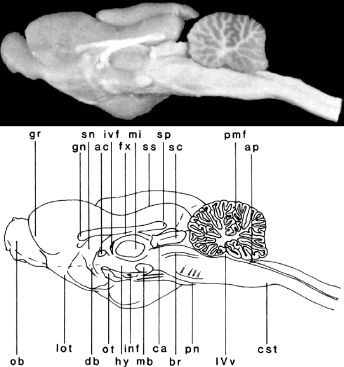
Fig. 3.1. A photograph (A) and line drawing (B) of a dorsal view of the ferret brain. See Table 3.1 for abbreviations.Fig. 3.2. A photograph (A) and line drawing (B) of a ventral view of the ferret brain. See Table 3.1 for abbreviations.Fig. 3.3. A photograph (A) and line drawing (B) of a lateral view of the ferret brain. See Table 3.1 for abbreviations.Fig. 3.4. A photograph (A) and line drawing (B) of a medial view of the ferret brain after sagittal bisection. See Table 3.1 for abbreviations.
The rostral end of the lateral gyrus is bounded by the ansinate sulcus, which forks medially from the lateral gyrus (Fig. 3.1 and Fig. 3.3). Extending rostrally from the lateral gyrus, the posterior and anterior sigmoid gyri are oriented in a lateral-to-medial direction, and they are separated by the cruciate sulcus (Fig. 3.1 and Fig. 3.3). The border between primary somatosensory and primary motor cortices is located on the sigmoid gyrus [13]. Rostral to the anterior sigmoid gyrus are the presylvian sulcus, the orbital gyrus, olfactory sulcus, and gyrus rectus (Fig. 3.5), which contain prefrontal cortical areas.
Fig. 3.5. Functional cerebral cortical areas identified in the ferret cerebral cortex. See text for details. Abbreviations are given in Table 3.1. Area AMLS in the medial bank of the suprasylvian sulcus is not visible but is located medial to ALLS [30].
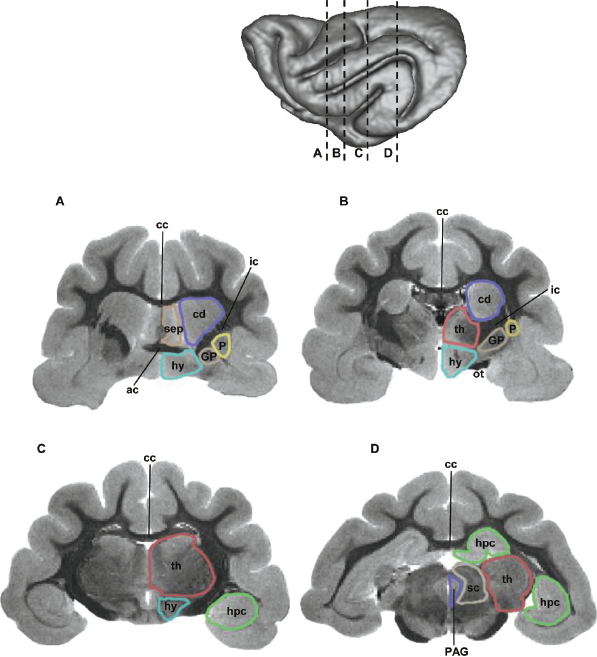
The arrangement of subcortical structures relative to the cerebral cortex is illustrated in Fig. 3.6, in which white matter appears darker than gray matter. In the coronal slice shown in Fig. 3.6A, a well-defined internal capsule separates the caudate from the lentiform nucleus, which consists of the putamen and globus pallidus. More caudal coronal planes, which intersect the thalamus, are shown in Fig. 3.6B,C. The rostral and ventral hippocampus, proximal to the amygdala, are shown in Fig. 3.6C, and both the dorsal and ventral aspects of the hippocampus are observed in the caudal coronal plane shown in Fig. 3.6D, which additionally intersects the superior and inferior colliculi. An axial image intersecting the majority of the subcortical structures outlined in Fig. 3.6 is shown in Fig. 3.7A.
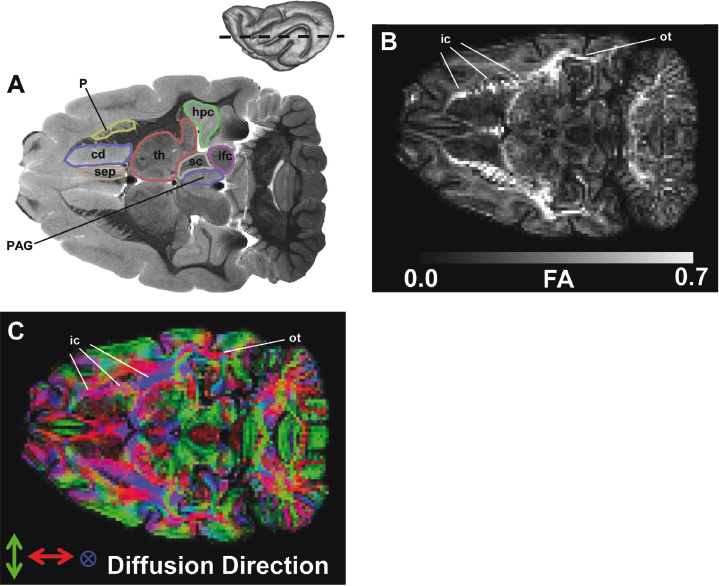
Fig. 3.6. Coronal, T2-weighted images of ferret brain. The locations of each slice are indicated in the inset. See Table 3.1 for abbreviations.Fig. 3.7. A T2-weighted image (A), fractional anisotropy (FA) map (B), and diffusion direction map (C) of an axial plane through the ferret brain. The location of the slice is indicated in the inset. The FA values range from 0 to 0.7 as indicated by the color scale in B. The color codes in the diffusion direction map are as follows: green encodes left/right, red encodes the rostral/caudal, and blue encodes the dorsal/ventral components of the vector aligned along the least restricted direction of diffusion. The three line segments indicate the rostral, middle, and caudal regions of the internal capsule (ic), as described in the text. See Table 3.1 for abbreviations.
Several cerebral white matter structures, including the internal capsule, anterior commissure, corpus callosum, and optic tract are apparent in the T2-weighted MRI data (Fig. 3.6 and Fig. 3.7). DTI is an additional neuroimaging technique commonly used to study the structure and integrity of white matter. Within myelinated areas, water displacements are larger parallel to myelinated fibers than perpendicular to them. This phenomenon is termed diffusion anisotropy, and fractional anisotropy (FA) is a quantitative index that expresses the amount of anisotropy in water diffusion. FA values range from 0 to 1, with FA = 0 indicating no anisotropy, and FA = 1 indicating extreme anisotropy. In Fig. 3.7B, an FA map of the axial plane corresponding to Fig. 3.7A is shown. Within white matter structures such as the internal capsule and optic tract, FA is large (white regions), whereas within neighboring gray matter structures, FA is lower. Additional information available through DTI data is the orientation of white matter tracts. In Fig. 3.7C, the least restricted direction of diffusion is indicated using a conventional diffusion direction map [14]. The intensity of red, green, and blue colors indicates the rostral/caudal, left/right, and dorsal/ventral components of the vector describing the direction of least restricted diffusion. As indicated in the diffusion direction map, the caudal optic tract contains fibers that are oriented in a rostral/caudal direction (red), whereas the fibers of the internal capsule are heterogeneously oriented, with those of the anterior limb aligned in a rostral/caudal direction (red), those of the posterior limb oriented in a dorsal/ventral direction (blue), and those of the genu being obliquely oriented. The appearance of clearly resolved white matter structures with varying orientations in DTI resembles patterns observed in primate brains and is in contrast to the structure of relatively simpler rodent brains.
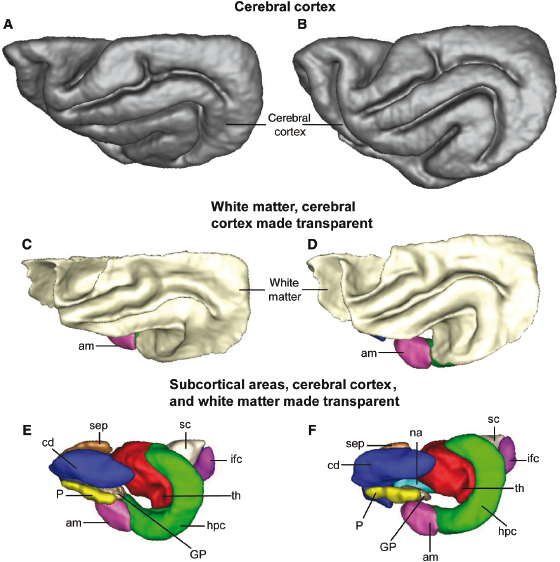
An overview of the spatial relationships between the cerebral cortex, cerebral white matter, and the subcortical gray matter structures outlined in Fig. 3.6 and Fig. 3.7 is given in Fig. 3.8 (dorsal views are Fig. 3.8A,C,E and lateral views are Fig. 3.8B,D,F). Each of the surfaces in Fig. 3.8 was generated by manually labeling T2-weighted image voxels that reside within each of the nuclei and creating contours of each of the labeled structures. In the top row (Fig. 3.8A,B), the cerebral cortex is rendered opaque, and therefore, underlying structures are not observable. In the second row (Fig. 3.8C,D), the cerebral cortex is rendered transparent, which reveals the underlying white matter, the amygdala, and part of the rostral hippocampus. In the third row (Fig. 3.8E,F), both the cerebral cortex and white matter surfaces are rendered transparent to expose the remaining labeled subcortical structures. The color coding is preserved among Fig. 3.6, Fig. 3.7, and Fig. 3.8.
Fig. 3.8. Three-dimensional surface contours of the cerebral cortex (A,B), white matter (C,D), and subcortical structures traced in Fig. 3.5 and Fig. 3.6 (E,F). Dorsal views are shown in panels A, C, and E and lateral views in panels B, D, and F. The images in panels C and D were obtained by rendering the cerebral cortical surface transparent. The images in panels E and F were obtained by rendering the cerebral cortex and white matter transparent.
Development of the Cerebral Cortex
As has been recently documented in the ferret, and in common with other species, pyramidal neurons of the cerebral cortex are generated from asymmetric mitotic events in ventricular [15,16] and subventricular [17] zones of the telencephalon, followed by radial migration to the cortical plate. Birthdating studies examining somatosensory and visual cortices have shown that both areas of the neocortex are generated in an inside-out manner, with deep cortical layers (layers V and VI) being born 12–15 days prior to neurons destined for more superficial layers II/III [1,15,16]. Furthermore, neurons within a given lamina of the rostrally located somatosensory cortex are born on average between 0.5 and 10 days prior to the corresponding lamina in the caudal visual cortex [15]. This regional difference in pyramidal neuron age is consistent with the rostral/lateral to caudal/dorsal transverse neurogenic gradient (TNG) described in neocortical development of vertebrate species ranging from mice [18] to human [19] and described for ferret in detail by McSherry [20].
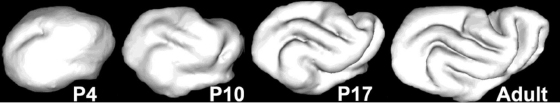
Subsequent to the genesis and migration of pyramidal neurons to the cortical plate, these cells undergo morphological differentiation [21] and form synaptophysin-positive synapses [22] over the period from approximately postnatal day 7 (P7) to P35. A rostral-to-caudal gradient parallel to the TNG has been reported in synaptogenesis [22] and morphological differentiation, as revealed through a gradient in water diffusion anisotropy within the cerebral cortex [5]. The gradient is presumably a consequence of the regional gradient in neuron age that results from the TNG. Concomitant with morphological differentiation of the cerebral cortex, the surface area of the cortical sheet expands rapidly over the period from P7 to P35 [23], and this period is also the time during which gyral and sulcal folds appear [24–26] (Fig. 3.9). In parallel with the rostral/lateral to caudal/medial gradient in morphological differentiation, similar gradients in cortical surface area expansion and folding have been described [6,23–26].
Fig. 3.9. Three-dimensional surface contours of cerebral cortex in postnatal day 4, 10, 17, and adult ferrets. (Barnette AR, Neil JJ, Kroenke CD, Griffith JL, Epstein AA, et al. (2009) Characterization of brain development in the ferret via MRI. Pediatr Res 66: 80–84.)
Following the initial primary expansion and folding of the cerebral cortex, subsequent changes in the brain external structure beyond P35 are less dramatic. However, white matter fiber tracts become myelinated over a period ranging from approximately P35 to P65, as has been revealed in T2-weighted MRI data, in which the white matter signal intensity decreases in association with myelination [4,6]. Concurrently, gyri become more tightly packed, and the brain takes on a more elongated shape, stretching in the rostral/caudal direction and narrowing in the medial/lateral direction [6].
Visual System
A number of recent studies using electrophysiological, anatomical tracer, and cytoarchitectural methods have led to the identification of several cortical areas responsive to visual stimuli [12,27–32]. Recent immunohistochemical studies have recognized 22 distinct visual areas, many of which are homologous to cortical areas in corresponding relative locations in the cat [10]. Consistent with this, a similarity between ferrets and cats has also been described with regard to the overall pattern of callosal connections in visual cortex [33]. The distribution of callosal connections has been associated with the subdivision of visual areas in many species, and several features of the callosal pattern in visual cortex of the cat (Olavarria and Van Sluyters [34]) resemble features of the callosal pattern in ferrets [11]. The primary or striate visual cortex (area 17) spans the occipital ridge of the ferret brain across its ventral/lateral to dorsomedial extent [35]. Along the lateral cortical surface, three cytoarchitecturally distinct representations of the visual field have been identified at progressively more rostrally located sites. These have been named areas 18, 19, and 21 from caudal to rostral, respectively, in recognition of their similarity in organization to the cat visual cortex [27,28]. Area 18 occupies the lateral gyrus and shares the rostral border of area 17. Area 19 shares its caudal border with area 18 on the lateral gyrus and its rostral border with area 21 on the suprasylvian gyrus, spanning the most lateral aspect of the intervening lateral sulcus. Area 21 extends further rostrally into the suprasylvian sulcus. Both areas 19 and 21 are less extensive than areas 17 and 18 in the medial/dorsal to lateral/ventral direction.
Retinotopic organization and connectivity of visual areas in the anteromedial lateral suprasylvian (AMLS) [31] and posteromedial suprasylvian sulcus (PMLS) [29] visual areas have been characterized in the ferret. Subsequent neurofilament and chondroitin sulfate proteoglycan (Cat-301) immunohistochemistry has led to the proposed parcellation of the suprasylvian sulcus into seven distinct visual areas (AMLS, ALLS, PMLS, PLLS, VLS, DLS, and PS) [10]. Support for this parcellation comes from the similarities drawn between the ferret and cat visual cortex organization [10], but further physiological and hodological studies are required to confirm the existence of a number of the proposed visual areas.
Analogous to primate species, dorsal and ventral visual processing streams have been proposed to exist in the ferret, in which the rostral and caudal posterior parietal cortical areas, PPr and PPc, respectively, are part of the dorsal/“where” stream [12]; and temporal cortical areas 20a, 20b, and PS are included in the ventral/“what” stream [26]. The posterior parietal cortical areas reside on the lateral and suprasylvian gyri and are bounded laterally by AMLS. The rostral boundary of PPr is shared with somatosensory area SIII. Areas 20a, 20b, and PS are the most ventral/lateral neocortical areas characterized in the ferret. The caudal border of 20a is shared with the rostral/lateral border of area 18, and the medial border of area 20a is shared with the lateral border of area 21. Areas 20b and PS are located progressively rostrally from area 20a. The anterior ectosylvian visual area (AEV) is located in the rostral bank of the pseudosylvian sulcus [35], isolated from other visual areas. This region contains neurons that are responsive to visual as well as other sensory stimuli. Based on these properties, it is considered to be homologous to AEV in the cat [35].
Auditory System
Electrophysiology [36], 2-deoxyglucose measurements of glucose metabolic rate [37], and optical imaging measures of functional activation [38,39] have been used to localize cerebral cortical areas subserving auditory functions in the ectosylvian gyrus. The two primary auditory areas, A1 and AAF, are located on the dorsal-most middle ectosylvian gyrus (MEG). Ventral and rostral to these two areas are nonprimary auditory areas AVF and ADF, and ventral and caudal to the primary auditory cortex are areas PPF and PSF. AAF and A1 exhibit clear tonotopic organization in which neurons responsive to high frequency tones are located dorsal to neurons responsive to lower frequencies [36,38,39]. Although neurons of the posterior, nonprimary areas PPF and PSF are less frequency-selective than those of primary auditory cortex, tonotopic organization is observed in a high-low-high frequency pattern across the ectosylvian gyrus, and the frequency reversal serves as a basis for distinguishing PPF and PSF [36]. Areas ADF and AVF do not exhibit tonotopic organization, and many neurons in these areas, particularly in AVF, are unresponsive to pure tones but do respond to other audio stimuli such as broadband noise [36]. Due to the proximity of AVF to multisensory areas [40] as well as AEV [35], the distinction between AVF and cortical areas characterized using different modalities and stimuli of other sensory systems remains somewhat unclear.
Somatosensory System
As with other sensory systems, several similarities in the organization of somatosensory cortex have been noted between the ferret and the cat. Relative to the ferret, the cat somatosensory cortex has been more completely characterized, with five somatosensory cortical areas (SI, SII, SIII, SIV, and SV) having been identified. Electrophysiological and cytoarchitectural studies conducted in the ferret identified neurons rostral to the ansate sulcus that are responsive to cutaneous and deep stimulation and are arranged in a somatotopic map. In this map, medial neurons of the posterior sigmoid gyrus respond to forepaw stimulation [41], and more lateral neurons in the suprasylvian gyrus are face-responsive [42,43]. This region comprises the ferret primary somatosensory cortex (SI). Caudal to this area, a region containing somatosensory-responsive neurons with larger receptive fields was identified, and subsequent electrophysiological and anatomical tracer experiments have revealed a second somatosensory cortical representation that extends from SI rostrally to the rostral-most parietal visual area PPr caudally [13]. Based on similarities in receptive field size and connectivity patterns to other cortical and subcortical structures, it has been proposed that this area is homologous to cat somatosensory area SIII. It has further been suggested that area SII resides on the ectosylvian gyrus, rostral to auditory and multisensory areas AVF and AEV, respectively [13,35]. However, we are unaware of published studies delineating the borders of SII.
Multisensory Cortical Areas
It is increasingly recognized that neurons responsive to sensory stimuli other than the primary modality are present in most sensory cortical areas, including “low-level” (e.g., primary) sensory cortices. This phenomenon was explicitly studied in the ferret auditory cortex [44]. However, some nonprimary sensory cortical areas also have anatomical and physiological characteristics that suggest their involvement in the integration of sensory information across multiple sensory modalities. Numerous regions in the ferret cerebral cortex have been described as having properties of multisensory cortex. As mentioned previously, area AES on the caudal bank of the rostral ectosylvian gyrus contains neurons that respond to visual as well as auditory and somatosensory stimuli [35]. Anatomical tracer experiments have further demonstrated that primary somatosensory and visual cortices project to a common area in the pseudosylvian sulcus [40]. Similarly, it has been shown that the medial bank of the rostral suprasylvian sulcus (MRSS) is responsive to somatosensory as well as auditory stimulation [45]. Finally, the integration of somatosensory and visual stimuli has recently been characterized by electrophysiological and anatomical methods within PPr area [46], an area initially described as a component of the parietal visual processing stream [12,26].
Motor/Premotor Cortical Areas
Based on expected similarities to other carnivores, Lockard has assigned the primary motor cortex to be the area near the cruciate sulcus but caudal to the presylvian sulcus [3]. Although we are unaware of work that specifically delineates the borders of this cortical area in the ferret, others have also indicated their expectation that primary motor cortex resides in this area [13,47], and Foxworthy and Meredith label a region extending from the posterior sigmoid gyrus to the ventral/lateral extent of the rostral coronal gyrus as area M1 [13]. Rostral to the presumed primary motor cortex, a region spanning both banks of the presylvian and olfactory sulci from their dorsal/medial to ventral/lateral extents has been defined as the ferret prefrontal cortex based on its reciprocal connections with the mediodorsal nucleus of the thalamus [47].
Summary
As a model for evaluating brain development and structure, the ferret offers unique advantages. Compared with rodent species, white matter in the ferret occupies a larger volume fraction of the intracranial volume, which lends itself to evaluation with DTI methods. In addition, much of the development of the cortical mantle of the ferret takes place postnatally, permitting the study of cortical processes that would otherwise have to be studied in utero. Finally, there is a growing body of literature regarding localization of cortical function to guide future research.
References
1. Jackson CA, Hickey TL (1985) Use of ferrets in studies of the visual system. Lab Anim Sci 35: 211–215.
2. Lawes INC, Andrews PLR (1998) Neuroanatomy of the ferret brain. In: Fox JG, ed. Biology and diseases of the ferret. Baltimore, MD: Williams & Wilkins, pp. 71–102.
3. Lockard BI (1985) The forebrain of the ferret. Lab Anim Sci 35: 216–228.
4. Barnette AR, Neil JJ, Kroenke CD, Griffith JL, Epstein AA, et al. (2009) Characterization of brain development in the ferret via MRI. Pediatr Res 66: 80–84.
5. Kroenke CD, Taber E, Leigland LA, Knutsen AK, Bayly PV (2009) Regional patterns of cerebral cortical differentiation determined by diffusion tensor MRI. Cereb Cortex 19: 2916–2929.
6. Neal J, Takahashi M, Silva M, Tiao G, Walsh CA, et al. (2007) Insights into the gyrification of developing ferret brain by magnetic resonance imaging. J Anat 210: 66–77.
7. Sawada K, Horiuchi-Hirose M, Saito S, Aoki I (2013) MRI-based morphometric characterizations of sexual dimorphism of the cerebrum of ferrets (Mustela putorius). NeuroImage 83: 294–306.
8. Jespersen SN, Leigland LA, Cornea A, Kroenke CD (2012) Determination of axonal and dendritic orientation distributions within the developing cerebral cortex by diffusion tensor imaging. IEEE Trans Med Imaging 31: 16–32.
9. Sawada K, Watanabe M (2012) Development of cerebral sulci and gyri in ferrets (Mustela putorius). Congenit Anom (Kyoto) 52: 168–175.
10. Homman-Ludiye J, Manger PR, Bourne JA (2010) Immunohistochemical parcellation of the ferret (Mustela putorius) visual cortex reveals substantial homology with the cat (Felis catus). J Comp Neurol 518: 4439–4462.
11. Bock AS, Kroenke CD, Taber EN, Olavarria JF (2012) Retinal input influences the size and corticocortical connectivity of visual cortex during postnatal development in the ferret. J Comp Neurol 520: 914–932.
12. Manger PR, Masiello I, Innocenti GM (2002) Areal organization of the posterior parietal cortex of the ferret (Mustela putorius). Cereb Cortex 12: 1280–1297.
13. Foxworthy WA, Meredith MA (2011) An examination of somatosensory area SIII in the ferret cortex. Somatosens Mot Res 28: 1–10.
14. Mori S, Wakana S, Nagae-Poetscher LM, van Zijl PCM (2005) MRI atlas of human white matter. Amsterdam: Elsevier.
16. Jackson CA, Peduzzi JD, Hickeh TL (1989) Visual cortex development in the ferret: I. Genesis and migration of visual cortical neurons. J Neurosci 9: 1242–1253.
17. Reillo I, de Juan Romero C, Garcia-Cabezas MA, Borrell V (2010) A role for intermediate radial glia in the tangential expansion of the mammalian cerebral cortex. Cereb Cortex 21: 1674–1694.
18. Smart IHM (1983) Three dimensional growth of the mouse isocortex. J Anat 137: 683–694.
19. Sidman RL, Rakic P (1982) Development of the human central nervous system. In: Haymaker W, Adams RD, eds. Histology and histopathology of the nervous system. Springfield, IL: Charles C. Thomas, pp. 3–145.
20. McSherry GM (1984) Mapping of cortical histogenesis in the ferret. J Embryol Exp Morphol 81: 239–252.
21. Zervas M, Walkley SU (1999) Ferret pyramidal cell dendritogenesis: changes in morphology and ganglioside expression during cortical development. J Comp Neurol 413: 429–448.
22. Voigt T, De Lima AD, Beckmann M (1993) Synaptophysin immunohistochemistry reveals inside-out pattern of early synaptogenesis in ferret cerebral cortex. J Comp Neurol 330: 48–64.
23. Knutsen AK, Kroenke CD, Chang YV, Taber LA, Bayly PV (2013) Spatial and temporal variations of cortical growth during gyrogenesis in the developing ferret brain. Cereb Cortex 23: 488–498.
24. Smart IHM, McSherry GM (1986) Gyrus formation in the cerebral cortex of the ferret: II. Description of the internal histological changes. J Anat 147: 27–43.
25. Smart IHM, McSherry GM (1986) Gyrus formation in the cerebral cortex of the ferret: I. Description of the external changes. J Anat 146: 141–152.
26. Manger PR, Nakamura H, Valentiniene S, Innocenti GM (2004) Visual areas in the lateral temporal cortex of the ferret (Mustela putorius). Cereb Cortex 14: 676–689.
27. Manger PR, Kiper D, Masiello I, Murillo L, Tettoni L, et al. (2002) The representation of the visual field in three extrastriate areas of the ferret (Mustela putorius) and the relationship of retinotopy and field boundaries to callosal connectivity. Cereb Cortex 12: 423–437.
28. Innocenti GM, Manger PR, Masiello I, Colin I, Tettoni L (2002) Architecture and callosal connections of visual areas 17, 18, 19 and 21 in the ferret (Mustela putorius). Cereb Cortex 12: 411–422.
29. Cantone G, Xiao J, Levitt JB (2006) Retinotopic organization of ferret suprasylvian cortex. Vis Neurosci 23: 61–77.
30. Philipp R, Distler C, Hoffmann K-P (2006) A motion-sensitive area in ferret extrastriate visual cortex: an analysis in pigmented and albino animals. Cereb Cortex 16: 779–790.
31. Manger PR, Engler G, Moll CKE, Engel AK (2008) Location, architecture, and retinotopy of the anteromedial lateral suprasylvian visual areas (AMLS) of the ferret (Mustela putorius). Vis Neurosci 25: 27–37.
32. Law MI, Zahs KR, Stryker MP (1988) Organization of the primary visual cortex (area 17) of the ferret. J Comp Neurol 278: 157–180.
33. Olavarria JF, Bock AS, Leigland LA, Kroenke CD (2012) Deafferentation-induced plasticity of visual callosal connections: predicting critical periods and analyzing cortical abnormalities using diffusion tensor imaging. Neural Plast, 2012: 250196.
34. Olavarria JF, Van Sluyters RC (1995) Overall pattern of callosal connections in visual cortex of normal and enucleated cats. J Comp Neurol, 363: 161–176.
35. Manger PR, Engler G, Moll CKE, Engel AK (2005) The anterior ectosylvian visual area of the ferret: a homologue for an enigmatic visual cortical area of the cat? Eur J Neurosci 22: 706–714.
36. Bizley JK, Nodal FR, Nelken I, King AJ (2005) Functional organization of ferret auditory cortex. Cereb Cortex 15: 1637–1653.
37. Wallace MN, Roeda D, Harper MS (1997) Deoxyglucose uptake in the ferret auditory cortex. Exp Brain Res 117: 488–500.
38. Nelken I, Bizley JK, Nodal FR, Ahmed B, Schnupp JWH, et al. (2004) Large-scale organization of ferret auditory cortex revealed using continuous acquisition of intrinsic optical signals. J Neurophysiol 92: 2574–2588.
39. Versnel H, Mossop JE, Mrsic-Flogel TD, Ahmed B, Moore DR (2002) Optical imaging of intrinsic signals in ferret auditory cortex: responses to narrowband sound stimuli. J Neurophysiol 88: 1545–1558.
40. Ramsay AM, Meredith MA (2004) Multiple sensory afferents to ferret pseudosylvian sulcal cortex. Neuroreport 15: 461–465.
41. McLaughlin DF, Sonty RV, Juliano SL (1998) Organization of the forepaw representation in ferret somatosensory cortex. Somatosens Mot Res 15: 253–268.
42. Leclerc SS, Rice FL, Dykes RW, Pourmoghadam K, Gomez CM (1993) Electrophysiological examination of the representation of the face in the suprasylvian gyrus of the ferret: a correlative study with cytoarchitecture. Somatosens Mot Res 10: 133–159.
43. Rice FL, Gomez CM, Leclerc SS, Dykes RW, Moon JS, et al. (1993) Cytoarchitecture of the ferret suprasylvian gyrus correlated with areas containing multiunit responses elicited by stimulation of the face. Somatosens Mot Res 10: 161–188.
44. Bizley JK, Nodal FR, Bajo VM, Nelken I, King AJ (2006) Physiological and anatomical evidence for multisensory interactions in auditory cortex. Cereb Cortex 17: 2172–2189.
45. Keniston LP, Allman BL, Meredith MA, Clemo HR (2009) Somatosensory and multisensory properties of the medial bank of the ferret rostral suprasylvian sulcus. Exp Brain Res 196: 239–251.
46. Foxworthy WA, Clemo HR, Meredith MA (2013) Laminar and connectional organization of a multisensory cortex. J Comp Neurol 521: 1867–1890.
47. Duque A, McCormick DA (2010) Circuit-based localization of ferret prefrontal cortex. Cereb Cortex 20: 1020–1036.