Rachel Santymire, Heather Branvold-Faber, and Paul E. Marinari
Natural History
The black-footed ferret (Mustela nigripes) is a member of the weasel family (Mustelidae) and has long been considered one of the most endangered mammals in North America (Fig. 10.1).
Fig. 10.1. A wild female black-footed ferret in Conata Basin, Buffalo Gap National Grassland, South Dakota. Note the eyeshine, a distinctive feature of the species. (Photograph by T. Livieri/Prairie Wildlife Research.)
Often confused with the other two ferret species found in the world, the Siberian polecat (Mustela eversmanni) and the European polecat (Mustela puturious; the common ferret that has been domesticated as a companion animal), the black-footed ferret is considered a specialist and the only ferret native to North America, roaming the shortgrass and mixed grass prairie [1]. This solitary, nocturnal carnivore is almost entirely dependent on prairie dogs (Cynomys spp.) for food and uses the intricate prairie dog burrow system to raise its young and escape from predators, such as coyotes, badgers, and owls (Fig. 10.2).
Fig. 10.2. A black-tailed prairie dog in Badlands National Park, South Dakota. Prairie dogs comprise 66–90% of the black-footed ferrets' diet. (Photograph by T. Livieri/Prairie Wildlife Research.)
As European settlers moved westward, the prairie landscape was altered. Cropland conversion, habitat fragmentation, and poisoning campaigns to rid the landscape of prairie dogs was detrimental to the black-footed ferret and other plant and animal species closely associated with this unique ecosystem. Also, in the early 1900s, an exotic disease, the sylvatic plague, was inadvertently introduced into the rodent population of North America. By the 1960s, the prairie dog habitat that supported these animals was reduced from 40 million to 600,000 ha, which is less than 2% of its original geographic range [2].
By the late 1950s, black-footed ferrets were considered by most wildlife experts to be extinct. However, a small population of ferrets was discovered in 1964, associated with a black-tailed prairie dog (Cynomys ludovicianus) town in Mellette County, South Dakota [3]. Field biologists noticed continued fragmentation of the prairie dog colonies in South Dakota, and a marked decline in the number of black-footed ferrets during annual surveys. A total of nine individuals (five males, four females) were captured between 1971 and 1973 to initiate a captive breeding program at the U.S. Fish and Wildlife Service's Patuxent Wildlife Research Center in Maryland.
Biologists successfully paired members of the Mellette County population, although none of the 10 kits that were produced in 1976 and 1977 survived for more than a couple of days. The last known black-footed ferret from this population died in captivity in 1979. Once again, the black-footed ferret was presumed extinct, as no other wild members of this South Dakota population were ever seen during annual field surveys.
Then, in 1981, a discovery was made outside of Meeteetse, Wyoming. A ranch dog named Shep, owned by John and Lucille Hogg, brought home a carcass of a black-footed ferret. The Hoggs, unsure what the animal was, eventually brought the carcass to a local taxidermist, who identified it and called wildlife officials. Thanks to the Hoggs and Shep, the world was given another opportunity to rescue this species from extinction.
Biologists located and began monitoring the Meeteetse population. The population peaked in 1984 to a minimum of 129 individuals. However, a sharp decline in black-footed ferret numbers and the presence of both canine distemper virus (CDV) and sylvatic plague led to the trapping of all remaining black-footed ferrets [4,5]. A total of 24 black-footed ferrets were brought into captivity between 1985 and 1987. Six of the first group of individuals died within months of capture as a result of canine distemper. The fate of the species was left to the surviving 18 individuals; the last known black-footed ferrets in the world. These 7 males and 11 females were used to initiate the captive breeding program.
The amazing, collective efforts to save the black-footed ferret from extinction have been well documented [6–10]. In brief, the U.S. Fish and Wildlife Service, Wyoming Game and Fish Department, and the International Union for Conservation of Nature's (IUCN) Conservation Breeding Specialist Group, along with several stakeholders including zoos, developed the Black-footed Ferret Recovery Plan. Critical to the success of this plan was the strict genetic management criteria used for captive propagation of the species. The collaboration, scientific research, ex situ management, and reintroduction techniques developed and implemented for black-footed ferret recovery are considered a model on how to recover an endangered species. It also is noteworthy that the program greatly benefited from studying the more common European ferret and Siberian polecat to develop tools and knowledge that would speed the success of effectively managing and reproducing the black-footed ferret, including returning this charismatic mammal to the great plains of the American west.
Reproductive Biology
A detailed understanding of reproductive biology is the fundamental underpinning for consistently propagating any species. Specific information especially is important if the need arises for assisted breeding methods, such as artificial insemination (AI). Naturally, each species has evolved unique reproductive mechanisms. Yet, such information for wildlife species is as often rare as the animals themselves. When the recovery program was deemed necessary, it also was realized that there were no serious scientific data on black-footed ferret reproduction. The lack of research in black-footed ferrets meant the need to first study the ferret surrogates, the European and Siberian polecat species, which are known to be genetically akin to their black-footed counterpart [11]. This led to a series of investigations used to understand the basic reproductive traits [12] and develop assisted reproductive techniques, including the use of vaginal cytology to determine estrus [13], semen collection and characterization [14], and the development of laparoscopic intrauterine artificial insemination using fresh [15] and frozen-thawed semen [16]. Similar to these mustelids, the black-footed ferret is a seasonal breeder, responding to lengthening of the photoperiod, with breeding generally occurring March through May and births in April through July [12]. Males among the age of 1–5 years have similar testicular size and seminal quality, but older males have a continued loss of reproductive function [17]. Breeding records indicate that female fecundity declines after 4 years of age [17]. For males, testes enlarge, signaling the initiation of spermatogenesis and increased circulating testosterone concentrations usually occurring in December and January [12,18,19]. As females enter into estrus, vulvar swellingis observed [13]. Proestrus, in which vulva is initiated and the percentage of vaginal superficial cells increases, lasts ∼2–3 weeks, and females can remain in estrus, indicated by ≥90% vaginal superficial cells and maximum vulva size, for 32–42 days, and do not experience estrogenic bone marrow depression like the domestic ferret [12,13,20]. Like other mustelids, the black-footed ferret is an induced (or reflex) ovulator experiencing a spike in luteinizing hormone (LH) caused by copulation [12,21]. Estrus occurs once a year, but <10% of females can experience a second estrus [12]. Gestation is approximately 42 days (42–45 days), with an average litter size of 3.3 kits (one to 10 kits). In the absence of conception, an ovulating female experiences pseudopregnancy, which is a prolonged luteal phase lasting the entire gestational period [12,19].
Black-footed ferrets are sexually mature in the first year of life, but males may have a reduced length of fertility, with testosterone rising approximately 1 month later and then declining 1 month earlier than adult males [12,22]. This reduced timing of testosterone affects these yearling males' testicular tumescence and the production of spermic ejaculates comparable to adult males [22,23]. It is essential to determine if the male is producing adequate numbers of spermatozoa in the ejaculate prior to being used for breeding.
Cooperative Captive Breeding and Genetic Management
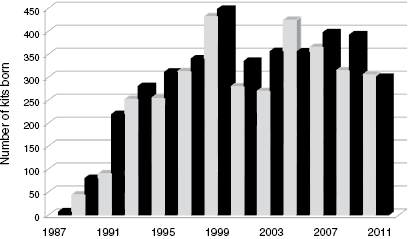
The current Black-footed Ferret Species Survival Plan® (SSP) contains approximately 300 individuals housed at six facilities across North America, including: the U.S. Fish and Wildlife Service's National Black-footed Ferret Conservation Center (Colorado), Smithsonian Conservation Biology Institute (Virginia, of the Smithsonian's National Zoological Park), Louisville Zoological Garden (Kentucky), Cheyenne Mountain Zoo (Colorado), Phoenix Zoo (Arizona), and Toronto Zoo (Ontario, Canada). These institutions collectively manage a core breeding population of 280 to 300 black-footed ferrets. The SSP's primary goal is to produce as many black-footed ferret kits as possible to sustain future captive breeding and to supply captive-born animals for ongoing reintroduction efforts (Fig. 10.3; [24]).
The SSP population has a skewed sex ratio, maintaining more females than males at a ratio of three males for every five females, thus maximizing reproductive output [24]. Prioritizing or determining which male should be paired with each female is accomplished using MateRx [25]. This analytical software program, developed jointly by the Smithsonian National Zoological Park (Washington, DC) and Lincoln Park Zoo (Chicago, IL) provides captive breeding facilities a numerical rating for every possible breeding pair. These ratings or mate suitability indices (MSI) integrate several genetic factors, including the expected change in genetic diversity due to resultant offspring produced, the relative rareness or commonness of the parent's genetic information, inbreeding coefficient of offspring produced by a pair, and proportion, if any, of unknown pedigree [24].
Reproductive Management Techniques
For females, the critical transition from proestrus to estrus is determined by closely monitoring the tumescence of the vulva and changes in vaginal cytology [12,13]. Cells are recovered via a vaginal, saline lavage of the awake female, transferred to a glass slide and then stained to determine the incidence of cellular cornification [13]. During estrus, superficial cells comprise more than 90% of observed epithelial cells, with all of these becoming fully keratinized, thereby signaling the optimal timing for pairing with a male [13]. In one study, pregnancy success was 60% when reproductive readiness of males was simply evaluated on the basis of enlarged testes compared to 80% when sperm were affirmed to be present in a prebreeding ejaculate [22].
Seminal evaluation has been conducted routinely and safely using electrojaculation of anesthetized males (using 40 mg/kg of ketamine/diazepam mixture). Using a 6 mm diameter rectal probe, low voltage electrical pulses are used to stimulate the nerves of the reproductive organs, producing an ejaculate [26]. A series of 100 electrical stimuli, from 2 to 5 V, are administered following an established protocol [14,26]. Due to the ejaculate's small volume, semen is collected using a micropipette from the glans penis and transferred to 100 µL of warmed (37°C) Test Yolk Buffer (Irvine Scientific, Santa Ana, CA), which is the standard medium used for semen handling in the black-footed ferret [17,27]. The extended semen is evaluated subjectively for percent sperm motility and forward progression (speed of forward progression on a scale of 0–5; 5 being the highest score) using described criteria [14]. Sperm concentration in the diluted suspension is evaluated using the hemocytometer method [28]. Raw, undiluted semen is preserved in 0.3% glutaraldehyde in phosphate buffered saline for subsequent evaluation of the incidence of morphologically normal versus abnormal sperm [14]. Sperm morphology and acrosomal integrity are assessed by phase contrast microscopy (×1000) of 100 sperm per sample, and classified as previously described [16,26,29].
Seminal characteristics vary among the black-footed ferret and the other two ferret species in this taxon. Specifically, the black-footed ferret has sperm motility and sperm progression (60–70%, 3.0, respectively) compared with Siberian polecat and domestic ferret (both at 80% and 3.0, respectively; [23]; Table 10.1).
Table 10.1. Ejaculate Traits in the Black-Footed Ferret (44 Males; 53 Ejaculates).
aSperm forward progression was based on a scale of 0 to 5; 5 = best.
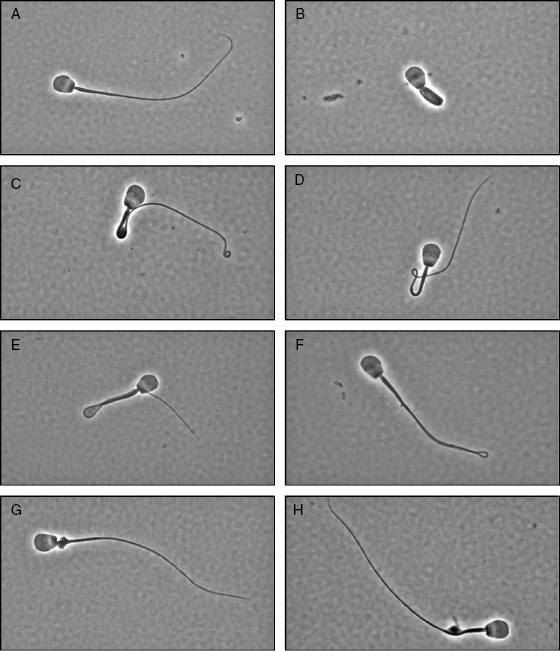
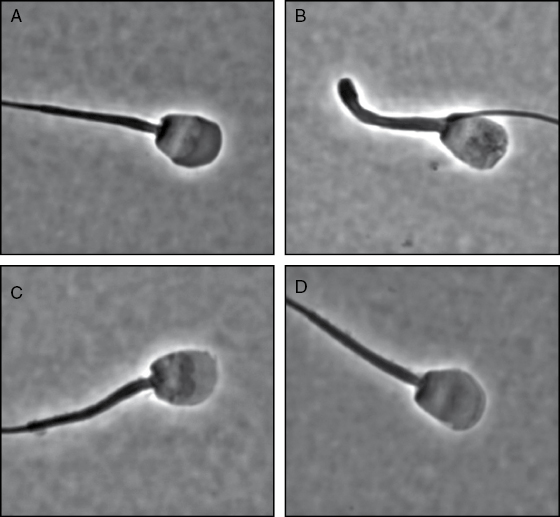
Additionally, the black-footed ferret has a lower proportion of structurally normal sperm (∼20%) compared with the two other species (both at 70%) (Table 10.1; [23,29,30]). Of the malformed spermatozoa in the black-footed ferret, 31% had an abnormal acrosome, 1–3% had a tightly coiled flagellum, 26% had a bent midpiece with a cytoplasmic droplet, 4% had a bent midpiece without a cytoplasmic droplet, 4% had a bent flagellum with a cytoplasmic droplet, 6% had a bent flagellum without a cytoplasmic droplet, 2% had a proximal cytoplasmic droplet, and 11% had a distal cytoplasmic droplet (Table 10.1; Fig. 10.4; [17,26,27,29]). It is known in other species, including the Florida panther (Puma concolor coryi, [31]), Asiatic lion (Panthera leo, [32]), and cheetah (Acinonyx jubatus, [33]), that structurally defective sperm do not participate in fertilization. The relatively high incidence of this trait in the black-footed ferret likely results from the limited number of founders and an overall low level of genetic variability as observed in other species (Peromyscus leucopus noveboracensis, [34]).
Fig. 10.4. Black-footed ferret sperm morphology. (A) normal; (B) coiled flagellum; (C) bent midpiece with cytoplasmic droplet; (D) bent midpiece without droplet; (E) bent flagellum with droplet; (F) bent flagellum without droplet; (G) proximal cytoplasmic droplet; and (H) distal cytoplasmic droplet. (Data from Reference 29.)
With the limited gene pool of the black-footed ferret population, it is important that each individual passes his or her genes on to the next generation. Using assisted reproductive techniques, such as AI, can ensure that the population genetic diversity is maintained by assisting with sexual incompatibility due to apathy or aggression [27]. In the 1988 Black-Footed Ferret Recovery Plan, it was stated that assisted reproductive techniques, such as AI with fresh or frozen sperm, had the potential to maximize genetic diversity [16,35], by ensuring that founders are equally represented and that every individual's genes are passed on to the next generation [36]. Therefore, the model species, domestic ferret and the Siberian polecat, were used to develop AI techniques for the black-footed ferret [14]. When vaginal insemination proved to be ineffective, the technique transabdominal intrauterine sperm deposition via laparoscopy was developed with high pregnancy success rates (>70%; [14]). As of 2004, 128 black-footed ferrets have been produced through AI using this laparoscopic technique [22].
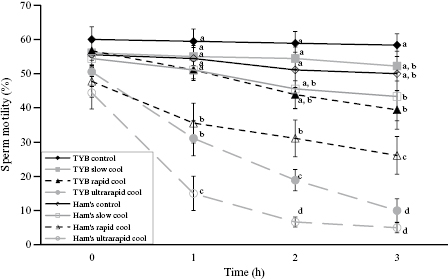
The domestic ferret has also been used as a model to develop a sperm cryopreservation protocol that could be applicable to the black-footed ferret. In one study, a seminal extender containing 3% to 4.5% glycerol as the cryoprotectant and slow cooling (10°C/h) in straws provided the highest percentage of motile sperm [37]. Other investigators determined that an egg yolk-based extender containing 4% glycerol, in combination with a pellet method of freezing, achieved the highest postthaw viability of domestic ferret spermatozoa [16]. Additionally, such sperm, when thawed and used for AI, resulted in a 70% pregnancy success rate and 31 healthy kits were born [15]. In another study using the same protocol, five of seven (71.4%) Siberian polecats became pregnant with a mean litter size of 5.3 kits. Subsequently, three of five black-footed ferrets became pregnant when inseminated with sperm using this same technique, producing a mean litter size of 2.3 [38]. However, results from AI with thawed sperm have been inconsistent since, emphasizing the need for more research into improved sperm cryopreservation for this species [39]. No doubt part of the challenge is due to the compromised sperm quality in the species as a whole. But recent studies also have revealed that black-footed ferret semen has usually high osmolality (500 mOsm) and is sensitive to hyperosmotic stress, a natural occurrence during cell freezing [29]. Additionally, it appears that slower cooling (to circa 0.2°C/min) appears optimal for retaining sperm motility and acrosomal integrity (Fig. 10.5; [30]).
Fig. 10.5. Mean (±SEM) sperm motility in nine black-footed ferrets postcollection and washed in Ham's F10 culture medium (Ham's) or test yolk buffer (TYB) and cooled at various rates either slow (0.2°C/min), rapid (1°C/min), or ultrarapid (9°C/min) cooling. Extended semen was returned to 25°C for 3 hours incubation. Control samples were maintained at 25°C. Slow cooling with TYB had improved sperm motility over time. Within each time period, different superscripts represent differences (P < 0.05) among treatments. (Data from Reference 30.)
Related to the use of AI with frozen semen is the concept of genome resource banking (GRB), which is the development of an organized repository of cryopreserved gametes—the opportunity to suspend the male's genome indefinitely to be used to produce offspring in later years, even after the donor's death. Thus, a GRB ensures the maintenance of extant genes for the future while also boosting contemporary management through the shipment of sperm from breeding center to center, without the expense and stress of translocating live animals. The Black-footed Ferret Recovery Implementation Team (BFFRIT) has been utilizing a GRB through the storage of semen samples from captive individuals and also from trapping wild-born black-footed ferrets and collecting and cryopreserving semen for potential use for AI in the captive population.
Preconditioning and Reintroduction
One of the most significant advances in black-footed ferret recovery has been the increased survivorship of captive born and reintroduced ferrets in the wild after preconditioning [40]. This practice involves placing captive-born ferrets into large-sized (∼1600 ft2) seminatural enclosures that contain a network of prairie dog burrows. Animals are then provided live prairie dogs, the number of which is determined by availability. Since 1996, all ferrets reintroduced into the wild have survived a minimum of 30 days preconditioning.
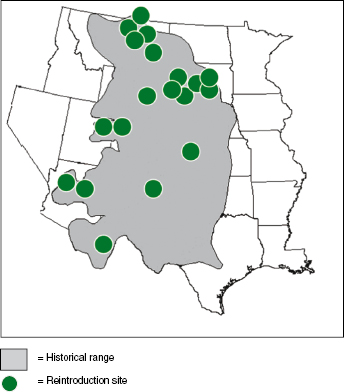
As the SSP increased the number of captive born individuals, BFFRIT members were able to achieve the next step in the road to recovery, reintroduction. In 1991, ferrets were reintroduced into Wyoming. This initial attempt was met with marginal success; however, wild-born ferrets were observed in subsequent years. The reintroduction of ferrets into the Shirley Basin of Wyoming served as a template and learning opportunity for other states to modify release procedures. Subsequently, more than 3000 ferrets have been released into South Dakota, Montana, Arizona, Utah, Colorado, Mexico, Kansas, New Mexico, Canada, and Wyoming (Fig. 10.6). Each year, ∼300–400 ferret kits are produced in the SSP. Approximately 200 kits are allocated for release on suitable prairie dog habitat, including lands owned by federal, state, tribal, nongovernment organizations, and private citizens. Additionally, about 200 wild-born ferrets have been translocated from self-sustaining release sites to other locations in an attempt to quicken the pace of recovery.
Fig. 10.6. Historical range (in gray) and the 19 reintroduction sites of black-footed ferrets as of 2012.
To determine the success of the effort, wild populations are monitored throughout the year using spotlight survey and snow-tracking techniques. Black-footed ferrets have a brilliant emerald green eyeshine that allows biologists to locate them on prairie dog colonies at night when they are most active. Individuals are identified by passive integrated transponder chips (microchips) that reveal if an individual is captive born or progeny of earlier reintroduced ferrets. Wild-born individuals may be live trapped and administered vaccines for sylvatic plague, canine distemper, and rabies.
Current Issues Influencing Species Recovery
Disease
Disease has limited the sustainability of wild black-footed ferret populations and has had negative impacts on the captive population. CDV has long been known to cause morbidity and mortality in this species, and contributed to the decline of the original Meeteetse population. Black-footed ferrets in captivity also have succumbed to CDV [41], and others injected with a modified live CDV vaccine died from vaccine-induced infection [42]. Even with supportive care, the mortality rate of CDV approaches 100%. PureVax® Ferret Distemper Vaccine, a live, monovalent canarypox-vectored vaccine developed by Merial, Inc., Athens, GA, has proven safe and effective in the Siberian polecat [43] and has been subsequently tested in the black-footed ferret. These trials have indicated that a minimum of two doses of vaccine produce protective neutralizing antibody titers [44]. Currently, all captive born ferrets and many wild ferrets receive two doses of PureVax® Ferret Distemper Vaccine beginning as early as 60 days of age.
Sylvatic plague (plague) is caused by the bacterium Yersinia pestis. It can be transmitted via flea vector, aerosol, or ingestion of contaminated food. Plague entered North America's west coast in the early 1900s on ships carrying flea-infested rats. It has since been spreading eastward, infecting and killing many native species, including prairie dogs and black-footed ferrets. Interestingly, the domestic ferret appears resistant to plague [45], and it has been long believed that the black-footed ferret was as well. But in the 1990s, the high susceptibility (with virtually 100% mortality) was discovered by Williams et al. [45] and then later affirmed by others [46]. This led to substantial research into developing a plague vaccine. The U.S. Geological Survey's National Wildlife Health Center (Madison, WI), in collaboration with the U.S. Army Medical Research Institute for Infectious Diseases, was instrumental in developing a vaccine consisting of the plague antigens F1 and V [46]. Challenge studies were conducted using two doses of the F1-V vaccine, with a 69% survival rate [46]. Now, all captive born and many wild ferrets are given two doses of F1-V vaccine as part of the recovery effort. Unfortunately, vaccination alone is insufficient to eliminate the threat of plague. A major obstacle to black-footed ferret recovery is the high susceptibility of prairie dogs to plague, making control of this disease, for both the predator and the prey, a high priority. Efforts are underway to develop a plague vaccine for prairie dogs.
Many diseases detected in the captive black-footed ferret population have not been observed (at least yet) in wild counterparts. Because ferrets are nocturnal and fossorial, it is difficult to detect and capture ill animals. However, diseases having an impact assuredly in captivity are discussed later.
Coccidiosis
Coccidiosis, caused by Eimeria ictidea and Eimeria furonis [47], has caused significant morbidity and mortality in captive black-footed ferret populations [48]. Clinical signs, apparent in both juveniles and adults, include diarrhea, anorexia, dehydration, and death. Unlike other species, it is not uncommon for a ferret to die acutely with no symptoms. Current treatment can consist of ponazuril, fluid therapy, and antibiotics to address possible secondary bacterial infection. Alternatively, sulfadimethoxine has been used.
Giardia spp.
Diarrhea is the most common symptom of this intestinal parasite, but anorexia can also occur. Treatment includes fluid therapy and fenbendazole or metronidazole. Unless there is concurrent disease, Giardiasis is not a significant cause of mortality.
Cryptosporidium spp.
The primary symptom in both juveniles and adults is chronic, mucoid diarrhea. Although rarely fatal, it is difficult to cure. Azithromycin, nitazoxanide, and paramomycin have been used to treat cryptosporidiosis with mixed results.
Cestodes and nematodes
Cestodes and nematodes are an occasional incidental finding and are treated with common antiparasitic drugs such as fenbendazole.
Influenza
This virus causes respiratory disease in black-footed and domestic ferrets. Prevention consists largely of minimizing the likelihood of exposure; keeping black-footed ferret populations closed to the public, having staff vaccinated yearly, wearing masks, and avoiding animal care when showing flu symptoms.
Amyloidosis
This noninfectious disease is commonly diagnosed in older necropsied individuals, although it is occasionally detected in 1- and 2-year-olds. Ferrets can be asymptomatic or have a variety of clinical signs, depending on the organs affected. Renal amyloidosis is a frequent cause of death in older individuals. Research is being conducted to help determine if there is a genetic predisposition for amyloidosis in the black-footed ferret. Overvaccination can contribute to amyloid deposits in other species, although it has yet to be identified as a cause in black-footed ferrets [49].
Neoplasia
Neoplasia is most often identified in older (over 4 years of age), nonreproductive black-footed ferrets [48,50], and therefore, has little impact on the captive breeding program. As most wild black-footed ferrets live for only 1 to 3 years (Livieri, pers comm), neoplasia likely is unimportant to the population in nature. Unlike in the domestic ferret [51,52], insulinomas and adrenal cortical tumors are rarely, if ever, observed in the black-footed ferret, and lymphoma is also uncommon. More than 180 different neoplasms have been identified in the black-footed ferret, ranging from benign epidermal cysts to highly malignant anal sac carcinomas [48,50]. The most frequently identified are renal tubular neoplasms, biliary cystadenoma/carcinoma, and apocrine gland tumors [50].
Gradually Reduced Fecundity over Time
The reproductive viability of the captive population appears to be decreasing with time, perhaps related to the limited number of founders and, thus, limited gene diversity. For example, over a recent 8-year interval, seminal quality has declined [17,29,30]. Specifically, the proportion of structurally normal sperm in an ejaculate has decreased from 50% to 16% with a simultaneous decline in the incidence of normal acrosomes (the sperm component that plays a critical role in fertilization) from 90% [17,27] to 70% (Fig. 10.7; [29,30]). The pregnancy success rate across the collective breeding centers has also dropped from 60% to 46% [24]. Because the black-footed ferret population is closed (i.e., there is no new source of founder genes), it is very likely that this overall reduction in reproductive vigor is related to individual relatedness or inbreeding depression. Carnivores are known to be particularly sensitive to incestuous matings and genetic homozygosity. Classic examples include: the poor ejaculate characteristics and especially high proportion of sperm pleiomorphisms in the cheetah [53], isolated populations of lions in Africa or India [32,54], and the virtually homozygous Florida panther [31]. Additionally, reduced gene diversity has resulted in fewer seminiferous tubules, spermatids, and interstitial tissue in African lions [55], and poor sperm morphology and reduced fertility in the red wolf (Canis rufus) compared with closely related, but more genetically variant canids (i.e., domestic dog; Canis familiaris and gray wolf; Canis lupus; [56–58]).
Fig. 10.7. Spermatozoan acrosomes with normal apical ridge (A), damaged apical ridge (B), missing apical ridge (C), and loose acrosomal cap (D). (Data from Reference 29).
Reduction in fecundity may also be attributed to a recent diet change for the SSP collective population from the original Sybille 60/40 black-footed ferret diet to the commercially available diet, Toronto Small Carnivore Diet (Milliken Meat Products Ltd., Markham, Ontario, Canada). Diets high in polyunsaturated fatty acids (PUFAs), like the Toronto Small Carnivore Diet, require elevated levels of vitamin E [59] to prevent nutritional steatitis (yellow fat disease), as observed in the domestic ferret [60]. Additionally, excessive vitamin A can prevent the uptake of vitamin E, a natural antioxidant [61]. Furthermore, mammalian sperm are susceptible to reactive oxygen species attack, resulting in decreased sperm motility, increased midpiece morphology defects, with deleterious effects on sperm capacitation and the acrosome reaction [62–64], and retention of cytoplasmic droplets [65]. To determine if the Toronto Small Carnivore diet was affecting serum levels of vitamin A and E, a preliminary study was conducted on 23 male black-footed ferrets. Results demonstrated that mean vitamin E (4.4 ± 0.8 µg/mL; range, 0.3–16.2 µg/mL) was lower for black-footed ferrets than mink and domestic ferrets (13–21 µg/mL); but the males had similar levels of vitamin A (0.36 ± 0.03 µg/mL; range, 0.09–0.61 µg/mL; other mustelids, 0.3–0.7 µg/mL; [66]). Because of low serum vitamin E, the next steps are to compare different diets and determine the effects on serum vitamin levels and semen quality.
Recovery Update
To date, over 8000 black-footed ferret kits have been produced by natural breeding and AI. The collaborative efforts that were used to develop a plan to recover the species in 1988 continue, and have led to advancements in reproductive technologies, nutrition, vaccine development, animal husbandry, genetic management, reintroduction techniques, and postmonitoring efficiency. Following the latest reintroductions in 2013, more than 3000 individual black-footed ferrets have been reintroduced at 21 locations (additional sites added in NM and CO) in the United States, Mexico, and Canada. Field biologists estimate that 400–500 individuals (Fig. 10.8) currently survive in the wild (Livieri, pers comm). The goal of the 1988 Black-footed Ferret Recovery Plan was to have 10 self-sustaining reintroduction sites by 2010. While enormous progress has been made, this program goal has yet to be reached. Although considered successful, the black-footed ferret recovery program faces the same obstacles that led to the species' near demise several decades ago: suitable disease-free prairie dog habitat. Fortunately, the diverse members that make up the BFFRIT are addressing these obstacles through development of an oral plague vaccine for prairie dogs, an innovative landowner incentive program, and continued genetic management and monitoring of the captive population. Biologists hope to maintain enough black-footed ferrets in the wild to move the species from endangered to threatened status by 2020. In 2013, the U.S. Fish and Wildlife Service updated the Black-footed Ferret Recovery Plan and implemented a safe harbor agreement, which allows any nonfederal landowner—including but not limited to private individuals, Tribes, states, counties, and municipalities—to volunteer their lands for the black-footed ferret recovery program serving as a reintroduction site without affecting land-use activities. This plan will lay out the framework for the recovery of the black-footed ferret so it will forever go into the night.
Fig. 10.8. A black-footed ferret passes through a specially designed transponder (microchip) reader placed over a burrow in Grasslands National Park, Saskatchewan, Canada. (Photograph by T. Livieri/Prairie Wildlife Research.)
1. Anderson E, Forrest SC, Clark TW, Richardson L (1986) Paleobiology, biogeography, and systematics of the black-footed ferret, Mustela nigripes. Great Basin Nat Mem 8: 11–62.
2. Marsh RE (1984) Ground squirrels, prairie dogs, and marmots as pest on rangeland. In: Proceedings of the Conference for Organization and Practice of Vertebrate Pest Control. Fernherst: ICI Plant Protection Division, 195–208.
3. Miller BJ, Reading RP, Forrest S (1996) Prairie night: black-footed ferrets and the recovery of endangered species. Washington, DC: Smithsonian Institution.
4. Carr A (1986) Introduction. Great Basin Nat Mem 9: 1–7.
5. Lockhart JM, Thorne TE, Gober DR (2004) A historical perspective on recovery of the black-footed ferret and the biological and political challenges affecting its future. In: Roelle JE, Miller BJ, Godbey JL, Biggins DE. Symposium on the Status of the Black-footed Ferret and Its Habitat. Scientific Investigations Report 2005-5293. Fort Collins: US Department of the Interior, 6–19.
6. Linder RL, Hillman CN (1973) Proceedings of the black-footed ferret and prairie dog workshop. Brookings, SD: South Dakota State University.
7. Anderson SH, Inkley DB (1985) Black-footed ferret workshop proceedings. Cheyenne, WY: Wyoming Game and Fish Department, pp. 1.1–33.12.
8. Seal US, Thorne ET, Bogan MA, Anderson SH (1989) Conservation biology and the black-footed ferret. New Haven, CT: Yale University Press.
9. Oldemeyer JL, Biggins DE, Miller BJ, Crete R (1993) Management of prairie dog complexes for the reintroduction of the black-footed ferret. Washington, DC: US Fish and Wildlife Service Biological Report 13.
10. Roelle JE, Miller BJ, Godbey JL, Biggins DE (2004) Recovery of the black-footed ferret: progress and continuing challenges. In: Roelle JE, Miller BJ, Godbey JL, Biggins DE. Symposium on the Status of the Black-footed Ferret and Its Habitat. Scientific Investigations Report 2005-5293. Fort Collins: US Department of the Interior.
11. O'Brien SJ, Martenson JS, Eichelberger MA, Thorne ET, Wright F (1989) Genetic variation and molecular systematics of the black-footed ferret. In: Seal US, Anderson SH, Bogan MA, eds. Conservation biology and the black-footed ferret. New Haven, CT: Yale University Press, pp. 21–33.
12. Williams ES, Thorne ET, Kwiatkowski DR, Anderson SL, Lutz K (1991) Reproductive biology and management of captive black-footed ferrets (Mustela nigripes). Zoo Biol 10: 383–398.
13. Williams ES, Thorne ET, Kwiatkowski DR, Lutz K, Anderson SL (1992) Comparative vaginal cytology of the estrus cycle of black-footed ferrets (Mustela nigripes), Siberian polecats (M. eversmanni), and domestic ferrets (M. putorius furo). J Vet Diagn Invest 4: 38–44.
14. Wildt DE, Bush M, Morton C, Morton F, Howard JG (1989) Semen characteristics and testosterone profiles in ferrets kept in long-day photoperiod, and the influence of hCG timing and sperm dilution on pregnancy rate after laparoscopic insemination. J Reprod Fertil 86: 349–358.
15. Howard JG, Hurlbut SL, Morton C, Morton F, Bush M, et al. (1989) Pregnancies in the domestic ferret after laparoscopic artificial insemination with frozen-thawed spermatozoa. Proceeding of the 14th Annual Meeting of the American Society of Andrology New Orleans, LA 52-P: abstract 135.
16. Howard JG, Bush M, Morton C, Morton F, Wildt DE (1991) Comparative semen cryopreservation in ferrets (Mustela putorius furo) and pregnancies after laparoscopic intrauterine insemination with frozen-thawed spermatozoa. J Reprod Fertil 92: 109–118.
17. Wolf KN, Wildt DE, Vargas A, Marinari PE, Kreeger JS, et al. (2000) Age dependent changes in sperm production, semen quality and testicular volume in black-footed ferrets (Mustela nigripes). Biol Reprod 63: 179–187.
18. Carvalho CF, Howard JG, Collins L, Wemmer C, Bush M, et al. (1991) Captive breeding of black-footed ferrets (Mustela nigripes) and comparative reproductive efficiency in 1-year-old versus 2-year-old animals. J Zoo Wildl Med 22(1): 96–106.
19. Brown JL (1997) Fecal steroid profiles in black-footed ferrets exposed to natural photoperiod. J Wildl Manage 61(4): 1428–1436.
20. Fox JG (1998) Biology and diseases of the ferret. Baltimore, MD: Lippincott Williams & Wilkins.
21. Carroll RS, Erskine MS, Doherty PC, Lundell LA, Baum MJ (1985) Coital stimuli controlling luteinizing hormone secretion and ovulation in the female ferret. Biol Reprod 32: 925–933.
22. Howard J, Santymire RM, Marinari PE, Kreeger JS, Williamson L, et al. (2004) Use of reproductive technology for black-footed ferret recovery. In: Roelle JE, Miller BJ, Godbey JL, Biggins DE. Symposium on the Status of the Black-footed Ferret and Its Habitat. Scientific Investigations Report 2005-5293. Fort Collins: US Department of the Interior, 28–36.
23. Howard J, Marinari PE, Wildt DE (2003) Black-footed ferret: model for assisted reproductive technologies contributing to in situ conservation. In: Holt WV, Pickard A, Rodger JC, Wildt DE, eds. Reproductive sciences and integrated conservation. Cambridge, MA: Cambridge University Press, pp. 249–266.
24. Garelle D, Marinari P, Lynch C (2011) Population Analysis and Breeding and Transfer Plan Ferret-black Footed (Mustela nigripes).
25. Ballou JD, Earnhardt J, Thompson S (2001) MATERX: Population Management Software. Washington, DC: National Zoological Park.
26. Howard JG (1993) Semen collection and analysis in nondomestic carnivores. In: Fowler ME, ed. Zoo and wild animal medicine: current therapy III. Philadelphia: W.B. Saunders, pp. 390–399.
27. Wolf KN, Wildt DE, Vargas A, Marinari PE, Ottinger MA, et al. (2000) Reproductive inefficiency in male black-footed ferrets (Mustela nigripes). Zoo Biol 19: 517–528.
28. Howard J, Bush M, Wildt DE (1986) Semen collection, analysis and cryopreservation in nondomestic animals. In: Morrow D, ed. Current therapy in theriogenology. Philadelphia: Saunders, pp. 1047–1053.
29. Santymire RM, Marinari PE, Kreeger JS, Wildt DE, Howard JG (2006) Sperm viability in the black-footed ferret (Mustela nigripes) is influenced by seminal and medium osmolality. Cryobiology 53: 37–50.
30. Santymire RM, Marinari PE, Kreeger JS, Wildt DE, Howard JG (2007) Slow cooling prevents cold-induced damage to sperm motility and acrosomal integrity in the black-footed ferret (Mustela nigripes). Reprod Fertil Dev 19: 652–663.
31. Roelke ME (1993) The consequences of demographic reduction and genetic depletion in the endangered Florida panther. Curr Biol 3: 340–350.
32. Wildt DE, Bush M, Goodrowe KL, Packer C, Pusey AE, et al. (1987) Reproductive and genetic consequences of founding isolated lion populations. Nature 329: 328–331.
33. O'Brien SJ, Wildt DE, Bush M, Caro TM, Fitzgibbon C, et al. (1987) East African cheetahs: evidence for two population bottlenecks? Proc Natl Acad Sci U S A 84: 508–511.
34. Malo AF, Martinez-Alvarez F, Alaks G, Dubach J, Lacy RC (2010) Effects of genetic captive-breeding protocols on sperm quality and fertility in the white-footed mouse. Biology of Reproduction 83: 540–548.
35. Wildt DE, Goodrowe K (1989) The potential for embryo technology in the black-footed ferret. In: Seal US, Thorne ET, Bogan MA, Anderson SH, eds. Conservation biology and the black-footed ferret. New Haven, CT: Yale University Press, pp. 160–176.
36. Howard JG, Marinari PE, Wildt DE (2001) Mustelid reproductive biology and integration of assisted reproductive technology in species recovery. In: The First International Symposium on Assisted Reproductive Technology for the Conservation and Genetic Management of Wildlife. Omaha: XY Inc, 159–165.
37. Atherton RW, Straley M, Curry P, Slaughter R, Burgess W, et al. (1989) Electroejaculation and cryopreservation of domestic ferret sperm. In: Seal US, Anderson SH, Bodan MA, eds. Conservation biology and the black-footed ferret. New Haven, CT: Yale University Press, pp. 177–189.
38. Howard JG, Kwiatkowski DR, Williams ES, Atherton RW, Kitchin RM, et al. (1996) Pregnancies in black-footed ferrets and Siberian polecats after laparoscopic artificial insemination with fresh and frozen-thawed semen. In: American Society of Andrology, 115.
39. Howard JG, Wolf KN, Vargas A, Marinari P, Kreeger J, Williamson L, Wildt DE (1997) Enhanced reproductive efficiency and pregnancies after artificial insemination in black-footed ferrets. American Association of Zoo Veterinarian, pp. 351–352.
40. Biggins DE, Godbey JL, Hanebury LR, Luce B, Marinari PE, et al. (1998) The effect of rearing methods on survival of reintroduced black-footed ferrets. J Wildl Manage 62(2): 643–653.
41. Williams ES, Thorne ET, Appel MJG, Belitsky DW (1998) Canine distemper in black-footed ferrets (Mustela nigripes) from Wyoming. J Wildl Dis 24: 385–398.
42. Carpenter JW, Appel MJG, Erickson RC, Novilla MN (1976) Fatal vaccine-induced canine distemper virus infection in black-footed ferrets. J Am Vet Med Assoc 169: 961–964.
43. Wimsatt J, Biggins DE, Innes K, Taylor B, Garrell D (2003) Evaluation of oral and subcutaneous delivery of an experimental canarypox recombinant canine distemper vaccine in the Siberian polecat (Mustela eversmanni). J Zoo Wildl Med 34: 25–35.
44. Marinari PE, Kreeger JS (2004) An adaptive management approach for black-footed ferrets in captivity. In: Roelle JE, Miller BJ, Godbey JL, Biggins DE. Symposium on the Status of the Black-footed Ferret and Its Habitat. Scientific Investigations Report 2005-5293. Fort Collins: U.S. Department of the Interior, 23–27.
45. Williams ES, Mills K, Kwiatkowski DR, Thorne ET, Boerger-Fields A (1994) Plague in a black-footed ferret (Mustela nigripes). J Wildl Dis 30: 581–585.
46. Rocke TE, Mencher J, Smith SR, Friedlander AM, Andrews GP, et al. (2004) Recombinant F1-V fusion protein protects black-footed ferrets (Mustela nigripes) against virulent Yersinia pestis infection. J Zoo Wildl Med 35: 142–146.
47. Jolley WR, Kingston N, Williams ES, Lynn C (1994) Coccidia, Giardia sp., and a physalopteran nematode parasite from black-footed ferrets (Mustela nigripes) in Wyoming. J Helm Soc Wash 61: 89–94.
48. Bronson E, Bush M, Viner T, Murray S, Wisely SM, et al. (2007) Mortality of captive black-footed ferrets (Mustela nigripes) at Smithsonian's National Zoological Park, 1989–2004. J Zoo Wildl Med 38(2): 169–176.
49. Garner MM, Raymond JT, O'Brien TD, Nordhausen RW, Russell WC (2007) Amyloidosis in the black-footed ferret (Mustela nigripes). J Zoo Wildl Med 38(1): 32–41.
50. Lair S (1998) Epidemiology and Pathology of Neoplasia in the Captive Population of the Black-footed Ferret (Mustela nigripes). Ontario: University of Guelph.
51. Weiss CA, Scott MV (1997) Clinical aspects and surgical treatment of hyperadrenocorticism in the domestic ferret: 94 cases (1994–1996). J Am Anim Hosp Assoc 33: 487–493.
52. Chen S (2008) Pancreatic endocrinopathies in ferrets. Vet Clin North Am Exot Anim Pract 11: 107–123.
53. Wildt DE, Bush M, Goodrowe KL, Packer C, Pusey AE, et al. (1983) Unique seminal quality in the South African cheetah and comparative evaluation in the domestic cat. Biol Reprod 29: 1019–1025.
54. Wildt DE (1994) Endangered species spermatozoa: diversity, research, and conservation. In: Bartke A, ed. Function of somatic cells in the testes. New York: Springer-Verlag, pp. 1–24.
55. Munson L, Brown JL, Bush M, Parker C, Janssen D, et al. (1996) Genetic diversity affects testicular morphology in free-ranging lions (Panthera leo) of the Serengeti Plains and Ngorongoro Crater. J Reprod Fertil 108: 11–15.
56. Koehler JK, Waddell WT, Jones MH, Smith R, Behrns S (1994) Spermophagy in semen of the red wolf, Canis rufus. Mol Reprod Dev 37: 898–906.
57. Koehler JK, Waddell WT, Jones MH, Smith R, Behrns S (1998) Semen parameters and electron microscope observations of spermatozoa of the red wolf, Canis rufus. J Reprod Fertil 114: 95–101.
58. Goodrowe KL, Hay MA, Platz CC, Behrns SK, Jones MH, et al. (1998) Characteristics of fresh and frozen-thawed red wolf (Canis rufus) spermatozoa. Anim Reprod Sci 53: 299–308.
59. Oyarzun SE, Self K, Valdes EV, Chavez ER (1995) An evaluation of the nutritional adequacy of the feeding program of the black-footed ferret (Mustela nigripes) at the Metropolitan Toronto Zoo. In: Nutrition Advisory Group Toronto.
60. Brooks HV, Rammel CG, Hoogenboom JJL, Taylor DES (1985) Observations on an outbreak of nutritional steatitis (Yellow fat disease) in fitch (Mustela putorius furo). N Z Vet J 33: 141–145.
61. Mazzaro LM, Dunn L, Furr HC, Clark RM (1995) Study of vitamin A supplementation in captive northern fur seals (Callorhinus ursinus) and its effect on serum vitamin E. Mar Mamm Sci 11: 545–553.
62. Lamirande ED, Gagnon C (1992) Reactive oxygen species and human spermatozoa. I. Effects on the motility of intact spermatozoa and on sperm. J Androl 13: 368–378.
63. Sikka SC (2004) Role of oxidative stress and antioxidants in andrology and assisted reproductive technology. J Androl 25: 5–17.
64. Sikka SC, Rajasekaran M, Hellstrom WJG (1995) Role of oxidative stress and antioxidants in male infertility. J Androl 16: 464–468.
65. Agarwal A, Saleh RA (2002) Role of oxidants in male infertility: Rationale, significance, and treatment. Urol Clin North Am 29(4): 1–12.
66. National Research Council (NRC) (1987) Vitamin tolerance of animals. Washington, DC: National Academy Press.