Xingshen Sun, Ziying Yan, Xiaoming Liu, Alicia K. Olivier, and John F. Engelhardt
Assisted Reproductive Technologies for Ferrets
The domestic ferret (Mustela putorius furo) has become an important animal model in many fields of biomedical research. The development of techniques to genetically engineer domestic ferrets has dramatically expanded the potential of this species to study the molecular basis of human diseases. Genetic engineering in ferrets has been made possible through advances in assisted reproductive technologies and molecular tools capable of efficiently targeting genetic changes to the genome of somatic cells. However, unlike other species such as the mouse, pig, sheep, and dog, assisted reproductive technologies for ferrets have only recently been established. The following section will review this area that provides the basis for genetic modeling in ferrets.
In Vivo Ferret Oocyte Maturation, Fertilization, and Preimplantation Embryo Development
Efficient assisted reproductive technologies in ferrets require knowledge of reproductive physiology, oocyte maturation, fertilization, and embryonic development in this species. Since mature oocytes are the building blocks for embryo genetic manipulation, the in vivo process of oocyte maturation is key to efficient genetic engineering. The estrus cycle in domestic ferrets is unique and controlled by the length of daylight cycle. In the wild, unbred jills maintain a constant estrus between late March to early August when day length is longest [1]. In captivity, estrus can be induced in ferrets by alternating short (8-hour light and 16-hour darkness) and long (16-hour light and 8-hour darkness) day cycles such that estrus begins ∼3 weeks after the shift to long light cycle [2,3]. Thus, in the wild, ferrets are classified as monoestrus species, while in captivity, ferrets can be induced to become a seasonally polyestrous species. This feature makes ferrets particular attractive for assisted reproductive technologies.
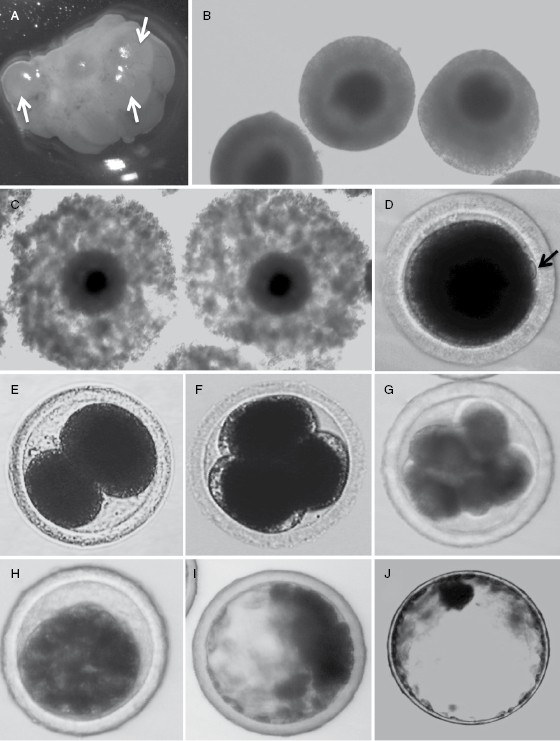
The pattern of follicular development during constant prolonged estrus is unknown. It is assumed that follicular development is continuous under these conditions with overlapping follicular maturation and atresia (i.e., breakdown of the ovarian follicles). Ferrets, like the domestic cat and rabbit, are induced ovulators, with ovulation occurring ∼30 hours following copulation [4]. However, the length of time required for maturation of primordial to preovulatory follicles remains unclear; Murphy proposed this time interval may be ∼6 days [5]. Without coital stimulation, the oocytes in preovulatory follicles remain at the germinal vesicle stage and may turn over by atresia during prolonged periods without mating. During copulation, the pressure on the cervix is associated with an increased production of luteinizing hormone (LH) [6]. Although LH increases modestly after copulation (approximately three- to four-fold), this is sufficient to initiate oocyte nuclear maturation, cumulus expansion, and ovulation [6]. Ferret oocytes contained within preovulatory ovarian follicles (Fig. 28.1A) are arrested at prophase of first meiosis (also called the germinal vesicle stage) and are surrounded by a compact cumulus (Fig. 28.1B). After copulation, increased circulating LH leads to a resumption of oocytes meiosis, formation of the first polar body, expansion of the cumulus (Fig. 28.1C), and ovulation of the oocyte. An average of ∼12 oocytes are ovulated into the ovarian bursa of virgin females, and this declines as animals are rebred [1]. Ferret oocytes are dark in appearance due to their high lipid droplet content (Fig. 28.1D) [7], making it more challenging to manipulate the pronucleus. Ferret oocytes are 140–160 μm in diameter and are surrounded by approximately three layers of follicular cells called the coronal radiata. These cells detach after fertilization, but if oocytes are not fertilized, the corona radiata remains for up to 4–5 days. The positive relation between the loss of the corona radiata and fertilization of the ferret oocyte aids in the identification of fertilized oocytes for assisted reproductive technologies. After ferret oocytes ovulate, they arrest at metaphase of second meiotic division (MII). Upon fertilization, MII stage oocytes will resume meiosis and finish their second meiotic division.
Fig. 28.1.In vivo ovarian follicle development, oocyte maturation, and preimplantation embryo development in the ferret. (A) Follicles (arrows) on the surface of preovulatory ferret ovaries at 24 hours after mating. (B) Oocyte–cumulus complexes (COCs) collected from large follicles at the germinal vesicle stage from jills in estrus (without mating). At this stage, the cumulus is still compact and the oocyte cytoplasm is darkly pigmented. (C) Following mating and thus LH stimulation, COCs with darkly pigmented oocyte cytoplasm have an expanding cumulus as the oocyte initiates meiosis. The cumulus at this stage is highly mucified, but the corona radiata remains closely attached to the oocyte. (D) Matured oocytes following formation of the first polar body (arrow). Embryos at (E) two cells, (F) three to four cells, (G) eight cells, (H) compacted morula, (I) early blastocyst, (J) and expanded blastocyst stages.
The ferret embryo has a relatively long preimplantation period of development. Six hours after copulation, spermatozoa can be found in the ovarian bursa and approximately 3.5–11.5 hours is required for sperm capacitation prior to fertilization [4,8]. Fertilization, as indicated by the appearance of pronuclei, occurs ∼40–44 hours after copulation and the second polar body is formed soon after the oocyte is penetrated by a spermatozoon [9]. Between 12 and 18 hours after spermatozoon penetration, a large male pronucleus and a smaller female pronucleus is formed. Spermatozoa remain viable with the ability to fertilize an oocyte for up to 126 hours after copulation, while oocytes retain the ability to be fertilized for only ∼12 hours after ovulation [8,10]. The first cleavage of fertilized single-cell embryos typically occurs at 50–52 hours after copulation (personal observation) (Fig. 28.1E). The first two blastomeres are often unequal sizes and also divide asynchronously (Fig. 28.1E), so uneven numbers of blastomeres in ferret embryos are often observed at later stage of development (Fig. 28.1F,G). Individual blastomeres are easily visible up to the 16-cell stage (Fig. 28.1G), but become difficult to visualize by the 32-cell stage due to increased compaction of the embryo. The embryos at this 32-cell stage are called compact embryos or morula (Fig. 28.1H). Fertilized embryos develop into morula by 5–5.5 days after copulation, at which time morula begins to transition from the oviduct into the uterus. Uterus blastocysts continue to develop in cell number from 88 cells to >6500 cells, while the diameter expands from 200 μm to >2 mm (Fig. 28.1I,J) [11]. The expanded blastocyst is spherical with the inner cell mass appearing as a dark spot at one pole of the blastocyst. By days 11–12 post copulation, the expanded blastocyst hatches and implants into uterine epithelium via trophoblasts that go on to form the zonary band of endotheliochorial placenta [1].
Embryo Culture and In Vitro Maturation of Oocytes
The development of efficient embryo culture techniques was the first critical step in the development of assisted reproductive technologies in ferrets. The first report of ferret embryo culture from in vivo generated one- to eight-cell stage embryos to blastocysts was described by Whittingham [12]. However, it was several decades before various types of embryo growth media were compared and conditions for superovulation and adoptive transfer were determined [13]. Optimum superovulation was achieved following sequential injection of virgin jills with 100 IU of equine chorionic gonadotrophin (eCG) and 150 IU of human chorionic gonadotrophin (hCG) 72 hours later. Oocytes or single-cell embryos were harvested 36 hours after the hCG treatment. Mating with a vasectomized hob immediately after hCG treatment did not significantly alter the number of oocytes ovulated, indicating that mating is not required for superovulation in ferrets. When mated to a fertile hob, superovulation gave rise to approximately 19 embryos per female, a number double that induced by mating alone [13]. In vitro culture of single-cell embryos in Chatot, Ziomek, Bavister medium (CZB) or tissue culture medium-19 (TCM-199) media supplemented with 10% fetal bovine serum (FBS) develops to blastocysts at rates of 64.5% and 47.1%, respectively. However, blastocysts developed approximately 12 hours faster in TCM-199 media, as compared with CZB media, and thus TCM-199 is now generally considered the better choice for culture of ferret embryos.
Mature oocytes are the foundation of somatic cell nuclear transfer (SCNT) technologies, and thus methods for the in vitro maturation of oocytes have also been developed. Li and colleagues evaluated the maturation of cumulus–oocyte complexes (COCs) harvested from small follicles (diameter of 0.5–2.0 mm) following superovulation as described earlier [14]. In this study, three types of media were evaluated including TCM-199 with 10% FBS; TCM-199 with 10% FBS, 10 IU/mL eCG, and 5 IU/mL hCG; or TCM-199 with 10% FBS, 10 IU/mL eCG, 5 IU/mL hCG eCG, and 2 μg/mL 17-beta-estradiol. Significantly greater maturation of oocytes (70%) was achieved in TCM-199 with 10% FBS, 10 IU/mL eCG, and 5 IU/mL hCG as compared with the other two media. Similar efficiencies of blastocyst formation (68%), following in vitro culture of 1- to 16-cell embryos in TCM-199-based medium, have also been observed with the close relative of the domestic ferret (European polecat) [15,16]. Additionally, there is one report demonstrating that in vitro matured European polecat oocytes are receptive to in vitro fertilization, with a 16% efficiency of developing to blastocysts [17].
Embryo Transfer
Embryo transfer (ET) involves the placement of in vivo- or in vitro-produced embryos into a surrogate female for gestation and delivery. This process is now well developed and widely used in most domestic species. In fact, the domestic ferret was among the first carnivorous species to adoptively transfer in vivo generated embryos [18]. ET has been the cornerstone for a number of assisted reproduction technologies, including cloning by SCNT. There are two methods for ET including surgical and nonsurgical. Kidder and colleagues developed a nonsurgical method for both collecting and transferring ferret embryos [19]. This approach involved endoscopic irrigation of the uterus with transcervical catheters for collection of embryos; the same system was used for adoptive transfer into recipient jills. The efficiency of births from adoptively transferred embryos in this study was 26%. Due to the specialized equipment needed for nonsurgical ET, most reports in ferrets use surgical ET. Several factors appear to affect the efficiency of surgical ET in the ferret including timing of hormonal treatment of the recipient, embryo stage at the time of transfer, and the number of embryos transferred to the recipient. Our work has evaluated the efficiency of ET following unilateral and bilateral transfer of zygotes or blastocysts into the oviduct or uterus, respectively, of nulliparous or primiparous recipient jills [20]. Twenty-one day fetal formation was significantly greater following bilateral oviduct transfer of zygotes in nulliparous females (71%) as compared with bilateral transfer in primiparous recipients (56%). Unilateral uterine transfer of blastocysts into nulliparous recipients gave rise to the highest percentage (90%) of fetal formation and was significantly greater than the transfer of zygotes into the oviduct (71%). Interestingly, transuterine migration of embryos occurred following all unilateral blastocyst transfers. Importantly, both in vivo generated and in vitro cultured eight-cell stage embryos from cinnamon-colored parents produced live kits when transferred into albino pseudopregnant recipients at rates of 61% and 32%, respectively [13].
Cryopreservation of Ferret Embryos
The development of effective cryopreservation techniques for the domestic ferret is critical for the long-term maintenance of valuable genetic specimens of this species. Two basic methods have been used for embryo preservation in ferrets: (1) slow-rate freezing and (2) vitrification. Initial efforts toward cryopreservation of Mustelidae began with the stoat (Mustela erminea) in 1993 [21], and it was not until a decade later that embryos from the European polecat (Mustela putorius) were successfully cryopreserved [22]. In the latter case, embryos grown to the expanded blastocyst stage were preserved using a slow-rate freezing technique with an efficiency of 10% producing live kits following adoptive transfer. The same group later reported a 16% survival rate with embryos vitrified by an open pulled straw method [23]. Recently, improvements in vitrification efficiencies with domestic ferret (M. putorius furo) embryos were described [7]. This approach used a micropipette tip-based method that significantly improves the development of thawed ferret embryos following ET, presumably through more rapid freezing rates. Ferret embryos at the morula, compact morula, and early blastocyst stages were evaluated in this study. Compact morula and early blastocyst stage embryos were determined to be better recipients for vitrification than morula stage embryos, based on in vitro development. Vitrified compact morula and early blastocyst stage ferret embryos resulted in live birth rates of 71.3% and 77.4%, respectively. This efficiency of ET with vitrified embryos was not statistically different from those observed with nonvitrified, in vivo isolated, control ferret embryos (79.2%) [7].
SCNT
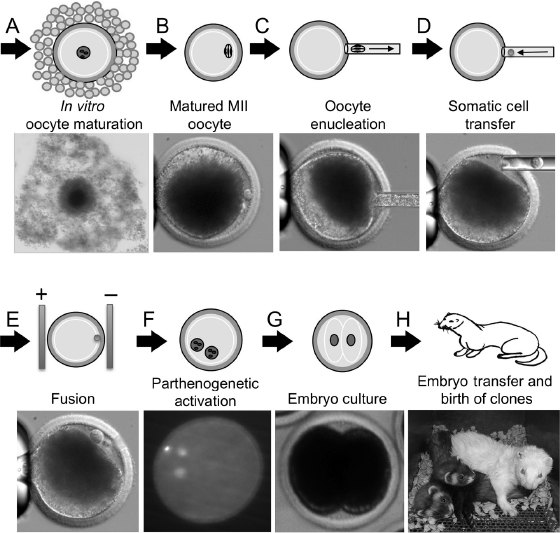
The discussed assisted reproductive technologies including embryo culture [13], oocyte maturation culture [14], and ET [20] established a key foundation for the development of SCNT technologies in ferrets. However, key additional embryologic methods critical to SCNT remained to be optimized including (1) parthogenetic oocyte activation [14], (2) enucleation of oocytes [24–26], (3) nuclear transfer and cell fusion of donor cells [24,27], and (4) the optimal cell cycle of the donor cell [25,26]. These techniques are the basis of SCNT procedures (Fig. 28.2). Optimization of these additional methods led to the first cloned ferrets in 2006 by cell fusion of cultured cumulus cells with recipient enucleated oocytes [24].
Fig. 28.2. Procedural schematic of SCNT in the ferret. (A) Isolation of oocyte–cumulus complexes (immature oocytes) and in vitro maturation. (B) Matured oocytes (MII stage with first polar body formed) are used as the recipient for nuclear transfer. (C) Oocyte enucleation removing the first polar body and MII chromosomal spindle. (D) Transferring somatic cell into perivitelline space of enucleated oocytes. (E) Electrofusion of somatic cell into oocytes. (F) Parthenogenetic activation of reconstructed embryos to initiate embryo development. (G) Reconstructed embryos are cultured to initiate the first cleavage and development to the two-cell stage. (H) Transfer of two-cell stage reconstructed embryos into pseudopregnant jills and the birth of cloned offspring.
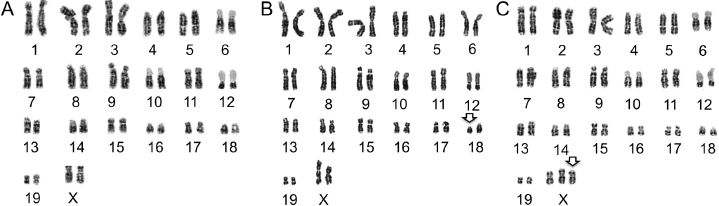
Direct piezoelectric microinjection of differentiated somatic cell nuclei into enucleated oocytes is the most efficient method of mouse SCNT [28,29]. However, for SCNT in ferrets, nuclear transfer by electrofusion is more efficient than direct nuclear injection [24,25,27]. The electrofusion method is also much more efficient when using genetically manipulated ferret fibroblasts, since such cells become senescent and larger after long selection periods, making them refractory to direct nuclear injection [30] (personal observations). Recipient oocyte quality is also a critical factor for SCNT. For unknown reasons, in vivo matured ferret oocytes are very fragile and rapidly fragment before and during in vitro manipulation for SCNT. However, in vitro matured oocytes are much more stable during manipulation (personal observations) and thus are preferred for SCNT. Additionally, although superovulation can provide a greater number of in vitro matured oocytes for SCNT, such oocyte recipients have thus far failed to produce cloned ferret births [26]. This is probably due to the adverse effects of eCG/hCG on developmental reprogramming. Thus, to improve the developmental competence of oocytes following SCNT, current optimal methods for SCNT use immature oocytes harvested directly from ovaries of nonhormonally treated jills following natural mating [24]. Additionally, we find that oocyte age following in vitro maturation is another important factor that affects developmental competence of the ferret embryo following SCNT. The optimal time frame for oocyte maturation is 40 hours, and developmental competence drops off significantly by 44 hours of maturation. Typically, pregnant SCNT recipients have only one or two fully developed fetuses, and this is usually insufficient to promote parturition. Thus, labor is most often hormonally induced in pregnant SCNT recipients and/or cloned kits are delivered by cesarean section. Currently, the efficiency of ferret SCNT averages ∼3%, but this efficiency can be significantly affected by the quality of the donor cell and whether or not they have been genetically manipulated [30]. Donor cells should also be karyotyped prior to SCNT to ensure that there are no major genetic abnormalities (Fig. 28.3A). However, even with this precaution, genetically abnormal cloned ferrets with chromosomal deletions or duplications have been observed (Fig. 28.3B,C). The largest litter of ferret clones obtained to date was seven kits derived from 58 SCNT embryos (12%) (unpublished data).
Fig. 28.3. The karyotypes of donor cells and ferret clones. (A) Normal karyotype of female donor fibroblasts used for SCNT. Ferrets have 20 pairs of nuclear chromosomes (19 pairs of autosomes and one pair of sex chromosomes). (B) Abnormal karyotype from a cloned kit demonstrating a chromosome 18 deletion (arrow). (C) Abnormal karyotype from a cloned kit demonstrating a three X chromosomes (arrow).
The Protocol for SCNT in the Ferret
The following optimal protocol of SCNT in ferrets is outlined. This protocol outlines methods for using fibroblast donor cells and in vitro matured oocytes as the recipient (Fig. 28.2).
Materials
Major equipment.
Piezopipet driver (PMM controller, Primetech, Tokyo, Japan).
Micromanipulation equipment: consisting of a Leica inverted microscope equipped with differential interference contrast optics and Eppendorf TransferMan (Brinkman, Westbury, NY) micromanipulator and microinjector.
ECM 2001 embryo manipulation system fusion machine (BTX/Genetronics, San Diego, CA).
Two stereo dissecting microscopes.
BTX fusion chamber with the gap between the two electrodes set at 1 mm.
Media.
Fibroblast cell culture medium: Dulbecco's modified Eagle's media (GIBGO/Invitrogen, Grand Island, NY) supplemented with 10% FBS, 1% penicillin, and 1% streptomycin.
Manipulation medium: Modified Dulbecco's phosphate buffered saline (PBS; GIBCO/Invitrogen, Grand Island, NY) (Dulbecco's PBS supplemented with 0.1% [w/v] D-glucose, 36 mg/L of pyruvate, and 0.4% [w/v] bovine serum albumin, BSA).
Oocyte maturation medium: Earle's TCM-199 supplied with 10% FBS, 10 IU/mL eCG (eCG, Sigma G-4527), and 5 IU/mL of hCG (hCG, Sigma-Aldrich, St. Louis, MO, C-8554).
Embryo culture medium: TCM-199 supplemented with 10% FBS, 1% penicillin, and 1% streptomycin.
Fusion medium: 0.28 M mannitol, 0.1 mM CaCl2, and 0.05 mM MgCl2.
Methods (SCNT Procedures)
Oocyte collection and maturation.
Ferrets are purchased from Marshall Farms. Female sable jills (virgin, 6–7 months of age, in estrus) are used for oocyte collection. Albino or sable jills (primipara, 9–12 months of age, in estrus) are used as surrogate and foster mothers. Vasectomized male ferrets (albino or sable, 12 months of age) are used for mating to induce follicular oocyte maturation in oocyte donor jills and to induce pseudopregnancy in surrogate recipients.
Reproductive management is a key component in the collection of viable oocytes and successful transfer of reconstructed embryos. The highest standard of animal husbandry practice must be maintained to ensure reproductive success and to satisfy regulatory guidelines and animal welfare recommendations. All ferrets are housed in separate cages under controlled temperature (20–22°C) and long daylight cycle (16-hour light/8-hour dark). Ferret chow was obtained from Marshall Farms.
Jills are mated with vasectomized hobs 24 hours prior to oocyte collection. Jills are then euthanized by administration of pentobarbital sodium injection (50–100 mg/kg, i.p., Ovation Pharmaceuticals Inc., Deerfield, IL, USA). Ovaries are then excised and immature oocytes are obtained from large preovulatory follicles (Fig. 28.1A) with sharp forceps to release COCs (Fig. 28.1C) into mPBS (Dulbecco PBS supplemented with 0.1% [w/v] D-glucose, 36 mg/L of pyruvate, and 0.4% [w/v] BSA).
COCs are then washed with mPBS several times and cultured in groups of 10–15 COCs in drops (500 μL) of oocyte maturation medium under light mineral oil at 38.5°C in 5% CO2, 95% air.
Enucleation.
After 15 hours in maturation culture, cumulus cells are removed from COCs by pipetting in mPBS containing 0.2% (w/v) hyaluronidase.
The denuded oocytes are then washed in mPBS and transferred into the mPBS containing 7.5 μg/mL of cytochalasin B (CB, Sigma C-6762) for 5 minutes at room temperature.
A small group of oocytes (10–15) is then placed into the manipulation chamber consisting of mPBS supplemented with 7.5 μg/mL CB and covered with mineral oil. The bottom of the micromanipulation chamber is glass.
Using differential interference contrast (DIC) optics, the oocytes are held in place with the first polar body at the three o'clock positions. The tip of an enucleation pipette (inner diameter of 20 μm) is then placed on the surface of the zona pellucida. A few piezopulses are used to pierce the zona pellucida and then the pipette is advanced into the perivitelline space. The polar body and a minimal volume of cytoplasm are then aspirated into the pipette. The chromosome spindle should be observable when the polar body is aspirated into the pipette.
Cell transfer and fusion.
After enucleation, donor cells are picked up with the same enucleation pipette and inserted into the hole made in the zona during enucleation. One donor cell is placed in the perivitelline space of the enucleated oocyte to form a couplet.
A group of couplets is then transferred to a dish of mPBS and then place on a heating stage (38.5°C) until fusion.
Couplets are moved into the fusion medium and one couplet is placed between the electrodes of the fusion chamber (1 mm gap fusion chamber, BTX). The couplet is manually aligned with a fine pipette so that the contact surface between the oocyte and the donor cell is parallel to the electrodes.
Cell fusion is then induced with an electrical pulse (30 μs electrical pulse of 1 DC, 180 V/mm).
After exposure to the fusion pulse, the couplets are then washed and placed into 25 μL drops of embryo culture medium and cultured at 38.5°C in 5% CO2, 95% air.
After 30 minutes, couplets are observed for fusion. Couplets that are not fused are subjected to a second electrical pulse.
Activation and embryo culture.
The fused embryos are then activated with 5 mM ionomycin for 4 minutes followed by treatment with 2 mM 6-dimethylaminopurine for 3 hours.
Activated embryos are then cultured for 24 hours in embryo culture medium at 38.5°C in 5% CO2 prior to transfer into pseudopregnant albino or sable recipient jills.
ET.
A pseudopregnant state is achieved in surrogate albino or sable primipara ferrets through mating with a vasectomized albino or sable male 24 hours prior to ET.
A stock solution of saline containing 10 mg/mL each of ketamine HCl (Abbott Laboratories, N. Chicago, IL, USA) and xylazine (Phoenix Pharmaceutical Inc., St. Joseph, MO, USA) is prepared. The surrogate ferrets are routinely anesthetized by injection of this solution to a final concentration of 20 mg/kg of ketamine and 20 mg/kg xylazine. During the surgery, 3% isofluorane is also used.
The abdomen is shaved and disinfected with 10% povidone iodine and 70% ethanol. A 1–2 cm incision is then made along the midline of the abdomen to expose the ovaries and oviducts. A 25-gauge needle is then used to generate a hole on the surface of the oviduct. Reconstructed embryos are then delivered into the oviduct through the hole using a fine glass pipette with an inner diameter slightly larger than that of the embryo.
After the surgical incision is sutured, the ferrets are returned to their cages and closely monitored until they are awake.
If natural birth does not occur at 42 days of gestation, the recipients are treated with F2 alpha-prostaglandin (Lutalyse; Pfizer; New York, NY, 0.5–1 mg, i.m.) to induce labor. If no kits are delivered within 3 hours, 0.3 mL of oxytocin is subsequently administered to the jill. If these treatments are unsuccessful to induce labor within 8 hours, a C-section is then performed.
If the recipient jills fail to produce enough milk to feed kits, the young are fostered onto another foster jill that has given birth within a 3- to 5-day window of the cloned animals.
Genetic Engineering in the Ferret
Techniques for genetically modifying animal genomes have been expanding since the first transgenic mouse was generated in 1982. There are two basic approaches to genetically engineer mice. The first includes pronuclear injection of DNA into the single-cell embryo. In this context, an expression cassette containing the promoter, gene of interest, and polyadenylation site randomly integrates into mouse genome. Offspring of the first generation (F1) founders (typically 5–12 independent founders) are then screened for expression of the transgene in the desired target organs and cell types. Mice have a rapid reproductive cycle (i.e., total gestation and time to sexual maturity) of ∼11 weeks, making this approach cost-effective. However, such an approach is significantly more costly in ferrets due to the significantly longer reproductive cycle of ∼30 weeks (42 days of gestation and ∼6 months to sexual maturity) and higher per diem rates of this species at most institutions. The second approach involves the manipulation of embryonic stem cells (ES cells) to produce “gene-targeted” mice (also called knockout or knock-in mice). In this approach, foreign DNA containing sequences homologous to the target locus are transfected into ES cells and undergoes homologous recombination at relatively high efficiency, thus introducing new genetic information at the targeted locus. Mouse ES cells appear to be uniquely suited to carry out homologous recombination and efficiently contribute to the germ line when injected into the inner cell mass of mouse blastocysts. However, ES cell lines with the ability to contribute to the germ cell linage have yet to be established for alternative nonrodent species including the ferret. Fortunately, progress in somatic cell gene targeting and SCNT cloning has provided a third approach for generating transgenic, knockout, and knock-in animal models in larger species [31–33]. This approach was the basis for generating the first genetically engineered ferrets, to be discussed later.
Generation of a Ferret Model of Cystic Fibrosis
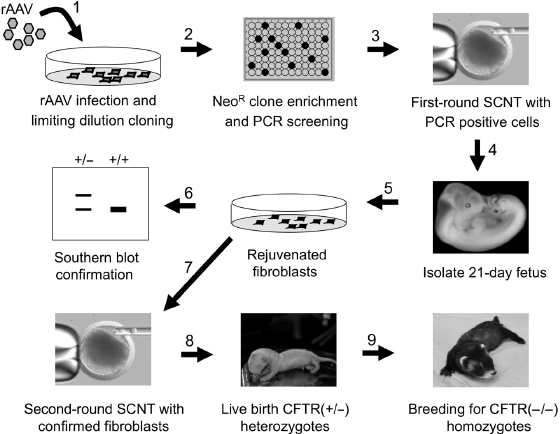
Cystic fibrosis (CF) is caused by defects in the CF transmembrane conductance regulator (CFTR) chloride channel [34]. While invaluable information has been gained using genetically engineered CF mouse models, this species does not fully recapitulate the natural progression of CF lung and pancreatic disease seen in human patients [35,36]. Ferrets are an attractive alternative species for modeling CF as its lung cell biology [37–40] and chloride channel properties of the airway epithelium [41] are highly similar to humans. Using viral-mediated gene targeting in fibroblasts, followed by SCNT, we recently created a CF ferret model [30] that develops similar organ pathologies as seen in humans CF patients [42]. The process of generating this model included three significant steps: (1) the generation of well-characterized CFTR gene disrupted fetal fibroblast cell lines that served as the nuclei donors for SCNT, (2) the generation of first-round cloned ferret embryos harboring the disrupted CFTR allele by SCNT and the characterization of the targeting event in the primary fibroblasts derived from these clones, and (3) the generation of second-round ferret clones using the rejuvenated fibroblasts generated from confirmed CFTR-targeted embryos. These cloned animals were then bred for two generations to give rise to CFTR-knockout ferrets.
Production of Ferret Fibroblasts with a Targeted cystic fibrosis transmembrane conductance regulator (CFTR) Gene
Initial attempts to target the CFTR gene in fibroblasts were based on transfection or nuclear injection of linear DNA fragments containing homology to CFTR exon-10 with an inserted selectable marker. These attempts failed badly (unpublished observations), indicating that the methods routinely adopted for gene targeting in mouse ES cells are not suitable for targeting in somatic fibroblasts. Indeed, the absolute frequency of homologous recombination in this setting (<2 × 10−7), even when positive–negative selection strategies were used to enrich for targeting events, remained two orders of magnitude less efficient than comparable approaches in mouse ES cells. Moreover, the level of nonhomologous integration remained high. These limitations were successfully overcome by large-scale screening of gene-targeting events facilitated by recombinant adeno-associated viral vectors (rAAVs).
Gene targeting with rAAV has overcome many of the limitations of linear DNA fragments for facilitating homologous recombination. AAV is a small, single-stranded DNA, helper-dependent parvovirus that has been widely used as a gene transfer vector for human gene therapy [43,44]. rAAV has unique biology that facilitates a remarkably high frequency of homologous recombination between the viral genome and genomic DNA [45,46]. We have also observed high frequencies of rAAV-mediated homologous recombination—up to 0.1% in human cell lines—even in the absence of selective enrichment [47]. This frequency is at least three to four orders of magnitude higher than that achieved by conventional targeting approaches.
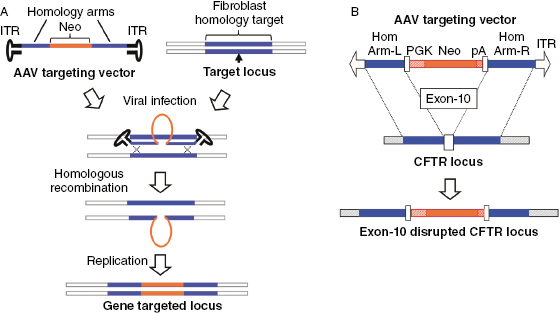
The design of rAAV-mediated gene-targeting vectors has several specific features that differ from the design of the more conventional targeting vectors used in ES cells. First, the optimal insert size for rAAV-targeting vectors (including regions of homology with the genome and a selectable marker expression cassette) is 4.4–4.5 kb. This limitation has to do with the optimal viral genome size for packaging, which is 4.7–4.8 kb and the fact that ∼300 bp of viral elements (inverted terminal repeats, ITRs) are required for packing of the recombinant DNA. For optimal efficiency of homologous recombination, a region with at least 1 kb of homology to the target sequence should be present on both sides of the selectable marker cassette (Fig. 28.4A), which confers drug resistance to enrich for recombinants prior to screening. This type of vector construction facilitates frequencies of rAAV-mediated gene targeting that are comparable with those achieved using conventional approaches involving positive–negative selection in mouse ES cells or a promoter-trap strategy in somatic cells. The AAV vectors used for targeted disruption of ferret CFTR exon-10 are shown in Fig. 28.4B. Isogenic homology arms were first isolated from 28-day fetal fibroblasts that were to be used in the gene targeting and these segments were cloned upstream and downstream to a phosphoglycerate kinase (PGK) promoter-driven neomycin (Neo) gene cassette also containing a poly-A sequence [30].
Fig. 28.4. rAAV-mediated gene targeted in fibroblasts. (A) Schematic representation of rAAV-mediated gene targeting. The rAAV vector harbors ∼2 kb of genomic DNA sequence homologous at the target locus (i.e., gene of interest), which is split into two halves flanking the targeted insertion site (marked by arrow). These two ∼1 kb stretches of DNA are cloned into the rAAV vector flanking a selectable marker (i.e., NeoR, neomycin resistance gene). Following rAAV infection, homologous recombination at the target locus occurs between the single stranded rAAV genome and one DNA strand of the target gene, thus inserting the selectable marker. After one round of DNA replication, the second DNA strand replicates the targeted insertion to disrupt the target gene. (B) The rAAV vector used to insert a neomycin expression cassette into exon-10 of the CFTR gene. The mutated CFTR locus disrupts expression of the CFTR gene and protein.
Male fetal fibroblasts were chosen for CFTR gene targeting with the rAAV2-CFTR targeting vector to allow for more rapid breeding of the gene-targeted animals. Fibroblasts were infected with rAAV2-CFTR targeting virus at a multiplicity of infection of 100,000 particles/cell and subsequently serially diluted into 96-well plates and placed under G418 selection (Fig. 28.5). During optimization of this procedure, we found that the number of cells seeded into each well and the timing of G418 section postplating were the most critical variables to the subcloning process that targeted 15–30% of wells with surviving fibroblast clones by 15 days (Fig. 28.5). Seeding 200–500 infected cells per well of a 96-well plates and initiation of selection (300 μg/mL G418) at day 2 following plating were optimal. After 15 days in selection, the 96-well plates were replicated into two plates by trypsinization and one plate was used for PCR screening of flanking sequences outside each targeting arm of the vector. The second plate was then used for expansion of targeted clones. In total, ∼500 clones were typically screened in a single experiment and targeting efficiencies following first-round PCR screens of neomycin resistant clones ranged from 0.05% to 2%. It remains unclear why different attempts at targeting in the same recipient cell line have resulted in large variability in targeting efficiencies, but we noted that the high frequency of repetitive elements in the ferret genome flanking CFTR exon-10 caused considerable deletions at the target locus, leaving clones with only one intact arm at the targeting locus.
Fig. 28.5. rAAV gene-targeting and SCNT cloning strategy used to generate CFTR-knockout ferrets. A sequential SCNT approach was used to generate CFTR gene-targeted ferrets using a 9 step approach. The various steps are marked by numbered arrows. Step 1: rAAV-CFTR targeting virus is used to infect primary fetal ferret fibroblasts. Step 2: Infected cells are cloned by limiting dilution G418 selection to obtain neomycin resistant (NeoR) fibroblast clones. These clones are replica plated and screened by PCR for both flanking insertion sites. Step 3: Positive clones with a “clean” insertion of the Neo gene within exon-10 are then used for a first round of SCNT. Step 4: 21-day embryo clones are harvested. Step 5: Rejuvenated embryonic fibroblasts are generated from embryo clones and amplified. Step 6: Fibroblasts are then screened by Southern blot to confirm the correct targeting event and karyotyped. Step 7: Rejuvenated fibroblast with Southern blot confirmed targeted events are then used for a second round of SCNT. Step 8: Live birth clones heterozygous for the targeted CFTR locus are reared. Step 9: Heterozygous CFTR+/− clones are then bred to wild-type jills and the heterozygous offspring are bred to generate homozygous CFTR−/−-knockout animals.
Regenerative Cloning Strategy of Gene-Targeted Ferret Fibroblasts
Following CFTR gene targeting, PCR selected ferret fibroblasts were then used for SCNT (Fig. 28.5). A significant obstacle to cloning animals from gene-targeted primary fibroblasts was the limited life span and rapid senescence of primary fibroblasts following selection. Consequently, gene-targeted primary fibroblasts are relatively poor SCNT nuclear donors, in general, and we observed that gene-targeted ferret fibroblast clones could not be expanded in culture to confirm gene-targeting events by Southern blot. To overcome this obstacle, a regenerative SCNT cloning strategy was used to rejuvenate fetal fibroblasts from 21-day embryos (Fig. 28.5). This process involved first-round SCNT cloning of targeted fibroblasts to produce 21-day embryos, followed by reestablishment of rejuvenated fetal fibroblasts from the cloned embryos. The rejuvenated fibroblasts were then used to confirm the targeting event and integrity of the targeting locus by Southern blot (Fig. 28.5). Those targeted embryo fibroblasts with a normal karyotype, as shown in Fig. 28.3A, were then used for a second round of SCNT cloning. The ability to clone embryos by SCNT from senescent gene-targeted fibroblasts was key to the success of modeling in ferrets and allowed for the generation of highly proliferative gene-targeted fetal fibroblasts with much higher live birth SCNT cloning efficiencies [30]. This two-step regenerative cloning strategy also appears to be much more efficient than direct SCNT cloning with gene-targeted cells in other models such as the pig [48]. In the case of generating the CFTR-knockout model, 11 recipient jills were each adoptively transferred with 35–60 reconstructed embryos derived from the optimal CFTR-targeted line and 14 live pups were born by natural birth or C-section [30]. These heterozygous founders were then bred to yield homozygous CFTR-knockout ferrets [42].
Generation of Transgenic Ferrets
The ability to clone transgenic ferrets that ectopically express a randomly inserted transgene cassette was also developed to study CF pathology. In these studies, a transgene cassette that specifically expresses the ferret CFTR complementary DNA (cDNA) in the intestine was introduced into CFTR-knockout primary fibroblasts, and these cells were used for SCNT to generate the first transgenic ferret [42]. This approach used an intestinal-specific promoter (intestinal fatty acid binding protein, FABPi) to drive expression of ferret CFTR in the intestine. The linear fragment containing the CFTR transgene cassette and a second selectable marker cassette was then transfected with LipofectamineTM (Invitrogen/Life Technologies, Grand Island, NY) into primary fibroblasts derived from a CFTR-knockout ferret. These polyclonal selected fibroblasts, containing multiple independent random insertions of the transgene, were then used for SCNT to obtain live births. The intestine was then evaluated for CFTR protein expression, and one out of four ferret clones produced sufficient CFTR protein to avoid meconium ileus. This study demonstrates the power of rapid transgenesis in the ferret model to study genetic disease processes [42].
Phenotype of CFTR-Knockout Ferrets
Cystic fibrosis transmembrane conductance regulator is a cyclic-AMP-regulated chloride channel that is expressed in many types of epithelia. Impairment of this channel results in defective transport of water and ions across epithelial membranes in the respiratory, pancreatic, gastrointestinal, hepatobiliary, sweat gland, and male reproductive systems [49]. Pathological changes include thickened mucous secretions, obstruction, and progressive inflammatory damage and scaring of organs. Currently, airways disease caused by chronic bacterial infections leads to the most morbidity and mortality in CF patients, with the development of diabetes being the second major comorbidity. To dissect CF pathogenesis and develop methods to treat the disease, animal models that reproduce the human CF phenotypes are required. The CF ferret model has begun to contribute to this field, and we will describe later the major phenotypes observed in the CFTR-knockout ferrets [42]. The CFTR-knockout ferret manifests meconium ileus, pancreatic destruction, liver disease, lung infections, and severe impaired nutrition—all features observed in humans with CF [49].
Lung Disease in CFTR-Knockout Ferrets
The lungs of newborn infants with CF are thought to be structurally normal [50], however, lethal bacterial infections may begin shortly after birth [51,52]. Similar to humans with CF, the CF ferret lung is histological normal at birth, but animals rapidly develop lethal lung infections early in life that manifest as bronchopulmonary pneumonia (Fig. 28.6A–D) [42]. Enteric bacteria are often observed in the lung of CF kits that die early in life, suggesting that CF kits are probably predisposed to aspiration-induced secondary infections. Following bacterial challenge, CF kits also fail to effectively eradicate bacteria (unpublished data), demonstrating that CFTR-dependent defects in innate immunity exist. In CF, bacterial eradication in the lung is hindered by defective CFTR-mediated anion transport and enhanced fluid absorption by the surface airway epithelium that is thought impair mucociliary clearance and innate immunity [49]. Functional studies on newborn CF ferret tracheas demonstrate defective cAMP-induced chloride permeability, suggesting that CFTR is the major channel needed for cAMP-mediated chloride movement across the airway of ferrets [42]. However, no significant difference was observed in amiloride-sensitive transepithelial resistance between CF and non-CF ferret tracheas, suggesting that enhanced Na+ absorption may not drive enhanced fluid uptake in the ferret airway. CFTR is highly expressed in serous cells of submucosal glands in the cartilaginous airways of humans and ferrets [53–55]. These structures have been proposed to be an important source of CFTR-dependent antibacterial factors and fluid that protect the airways from infection [56,57]. In the CF ferret trachea, defects in submucosal gland fluid secretion are observed similar to that seen in CF patients [42,57]. These findings suggest that the CF ferret model will be useful for studying the pathogenesis of lung infections in CF.
Fig. 28.6. Histopathology of lung and pancreatic disease in CFTR−/− ferrets. Representative lung histology from (A, B) CFTR+/+ and (C, D) CFTR−/− ferrets (5 days of age). There is locally extensive consolidation of the CFTR−/− lung by inflammatory cells, cellular debris, mucous, and bacteria (arrows) that extend from the airway (*) into the alveoli, consistent with bronchopulmonary pneumonia. Histopathology of the pancreas from newborn (E) CFTR+/+ and (F) CFTR−/− kits. Pancreas from the CFTR−/− kit has multifocal dilation of the acinar and ductular lumens by eosinophilic zymogen secretion (arrows). Pancreatic histopathology from (G) CFTR+/+ and (H) CFTR−/− ferrets of 32 days of age demonstrates the increased pancreatic damage that occurs with age. There is replacement of lobular parenchyma by bands of fibrosis (*) that surround markedly dilated ducts and acini (arrows). Bar = 500 μm (A, C), 100 μm (B, D), and 50 μm (E–H).
Pancreatic Disease in CFTR-Knockout Ferrets
Pathologic changes in the pancreas of newborn CF ferrets are relatively mild and include dilated acini and ductules with inspissated eosinophilic zymogen secretion (Fig. 28.6E,F) [42]. Such morphologic changes are similar to what is observed in the majority of human CF infants [58]. Pancreatic damage to the CF ferret exocrine pancreases increases with age, giving rise to inflammation, acinar and ductular apoptosis, loss of zymogen granules, and expansion of ductular epithelium [42]. Loss of lobular parenchyma and replacement by fibrous tissue occur in CF ferrets by 2–4 months of age (Fig. 28.6G,H). These progressive histopathologic changes in the CF ferret pancreas are quite similar to that seen in humans [59]. It is worth noting that the major histopathologic changes in the exocrine pancreas of CF ferrets preceded episodes of hyperglycemia and glycosuria, which emerged at 1–2 months of age [60]. Although CFTR can affect both exocrine and endocrine pancreas cellular components, the endocrine defects have been thought to be secondary to exocrine damage [61]. However, recent findings in newborn CF ferrets demonstrate significant abnormalities in blood glucose and insulin regulation prior to major exocrine pancreas disease [60]. Furthermore, isolated newborn CF ferret islets demonstrated dysregulated glucose-dependent insulin secretion in comparison with controls, supporting intrinsic abnormalities in CF islets [60]. Thus, the CF ferret model may be useful to study mechanisms of CF related diabetes, which occurs in 40–50% of CF patients over 30 years of age [62,63].
Meconium Ileus in CFTR-Knockout Ferrets
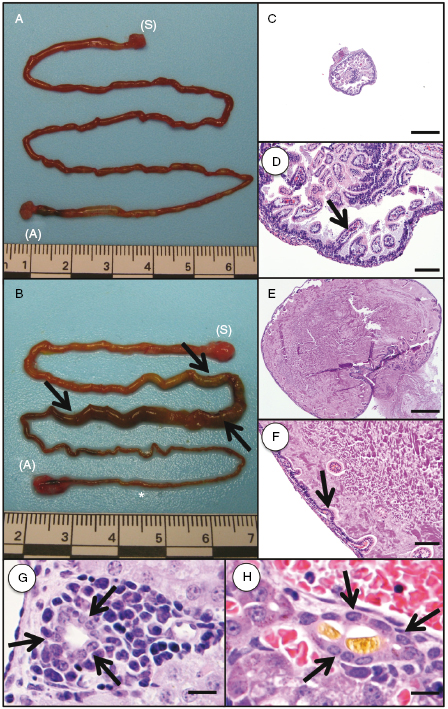
In humans, meconium ileus is the earliest manifestation of CF and occurs in ∼15% of newborn CF infants and can be detected as early as 17 weeks of gestation [59,64]. The incidence of meconium ileus in CF ferrets is ∼75% [42]. The reason for the enhanced prevalence of meconium ileus in CF ferrets is currently unclear, but this finding suggests that ferret CFTR plays a more significant role in hydration of the in utero intestine than human CFTR. Gross and histological examination of the newborn CF ferret intestine indicates that the obstruction is variably located throughout the small intestine and colon (Fig. 28.7A–F). Microcolon can also be observed in the CF ferret model, a pathology that is also seen in CF infant. Intestinal perforation and peritonitis in CF ferrets born with meconium ileus occur in the early postnatal period usually between 48 and 72 hours. In CF infants, meconium ileus is affected by heritable modifier genes [65,66]. Our studies have also demonstrated that meconium ileus in CF ferrets retain a significant genetic influence, with ranges of meconium ileus frequencies between 50% and 100% depending on the hob used for breeding [42]. These findings suggest that meconium ileus frequencies in the CF model could be reduced by selective breedings.
Fig. 28.7. Gross images and histopathology of intestinal meconium ileus and liver cholestasis. (A, B) Gross image of a normal intestine from a (A) CFTR+/+ kit (48 hours old) compared with intestine from a (B) CFTR−/− kit (48 hours old) with meconium ileus ([S] stomach, [A] anus). In the CFTR−/− intestine, there is dilation of a portion of the intestine (arrows) with narrowing of the distal intestine/colon (asterisk). (C, D) Histologic analysis from a CFTR+/+ intestine demonstrates a normal lumen diameter with normal intestinal villi (arrow). (E, F) Histologic analysis from a CFTR−/− intestine demonstrates marked dilation of the lumen by eosinophilic homogenous material and mucous with compression and loss of the intestinal villi (arrow). (G, H) Liver histology from a (G) CFTR+/+ kit demonstrating a bile duct (arrows) with an empty lumen compared with a bile duct (arrows) from a (H) CFTR−/− kit containing a bile plug (fibrillar material) within the lumen. Bile plugs within bile ducts are consistent with cholestasis. Bar = 500 μm (C, E), 100 μm (D, F), and 50 μm (G, H).
Liver Disease in CFTR-Knockout Ferrets
Liver disease in CF patients typically manifests later in life as focal biliary cirrhosis (20–30%), multilobular biliary cirrhosis (5–15%), portal hypertension (2–5%), and cholelithiasis (3–25%) [67]. Focal biliary cirrhosis is uncommon in CF infants, but it can be found in early childhood [68]. CF ferrets are born with elevated plasma alanine aminotransferase and bilirubin levels (both direct and indirect bilirubin), suggesting that early liver disease is present [42]. Such changes in CF ferrets were independent of meconium ileus and suggest some level of hepatic cholestasis. There is histologic evidence of cholestasis in the newborn CF ferret liver; however, this finding is variable (Fig. 28.7G,H) and can also be found occasionally in non-CF newborn livers. The majority of CF infants retain a very similar hepatic phenotype with >50% having abnormally elevated blood liver enzymes in the absence of histologically detectable hepatic cholestasis on biopsy [69]. Furthermore, oral ursodeoxycholic acid (UDCA) therapy normalizes these liver function markers in both newborn CF ferrets and humans [42,70,71]. CF newborn ferrets that retain meconium ileus have additional liver function test abnormalities reflected by elevated plasma alkaline phosphatase and gamma glutamyl transferase [42].
Growth and Nutrition in CFTR-Knockout Ferrets
Impaired growth and nutrition are nearly universal in CF patients due to reduced output of pancreatic enzymes and intestine dysfunction that reduce nutrient absorption [72,73]. Similar to infants with CF [74], newborn CF ferrets have early and severe growth impairment. Although newborn CF ferrets are born with similar weights to their non-CF littermates, during the first 2 weeks of life, CF kits fail to gain weight without clinical intervention [42]. CF ferrets must be treated orally with the proton-pump inhibitor omeprazole to raise gastrointestinal tract pH and gain weight. Omeprazole is thought to counteract the reduced bicarbonate in the gastrointestinal tract caused by reduced CFTR-mediated bicarbonate transport by the pancreas and intestine. At approximately 3–4 weeks, the diet of CF ferrets must also be supplemented with pancreatic enzymes to maintain weight gain, due to declining function of the exocrine pancreas [42]. The use of proton-pump inhibitors together with pancreatic enzyme supplementation is also the most common clinical strategy to treat malabsorption in CF patients.
Male Reproductive System CFTR-Knockout Ferrets
The majority (97%) of CF male adult patients suffer from infertility caused by bilateral absence of the vas deferens [75]. Most CF male infants have an intact vas deferens at birth; however, progressive mucoid obstruction of the vas deferens can lead to unilateral absence of the vas deferens as early as 2 years of age [76]. Newborn CFTR-knockout ferrets are born with a severely degenerated vas deferens at birth [42]. The vas deferens is absent in ∼50% of the spermatic cords of CF kits, with the remainder having segmentally absent vas deferens. These findings suggest that vas deferens abnormalities in CF male ferrets progress more rapidly than in CF male humans. Thus, the CF ferret model may be extremely useful in study CFTR involvement in the early development of the male reproductive organs.
Optimal Rearing Methods for CFTR-Knockout Ferrets
Although ∼75% of CF ferrets die from meconium ileus shortly after birth, the remaining 25% that pass stool within the early neonatal period also fail to thrive and die within the first week of life if untreated. Lung infections and malabsorption by the gastrointestinal tract appear to be the main causes of death in CF kits that escape meconium ileus. Thus, special clinical attention to reducing lung infections and improving nutrition is paramount to rearing methods for CF ferrets. The protocols to be discussed later for rearing CF ferrets have been previously described [42] and updated with more current experiences [60]. Since treatment conditions are often tailored for each CF animal depending on the clinical course of disease, it is also important that littermate non-CF controls be treated identically during the rearing of each CF animal to validate scientific comparisons and control for off-target treatment effects.
An important component of the rearing method is early intervention immediately after birth to maximize passage of meconium and enhance nutrition. Immediately after birth, all kits in a given litter are evaluated for the passage of meconium when tail clips are taken for genotyping. Those kits that do not pass meconium are then gavaged with 150–200 μL of Golytely (Abbott Laboratories, Columbus, OH) with a fire-polished microloader pipette tip (Eppendorf) (see Ref. 42 for example of gavage tube components). The gavage is then repeated every 6 hours, up to three times, until meconium is passed. Once genotypes are confirmed, CF and non-CF-matched littermates are identified (typically by 9–12 hours after birth) and the litter size is reduced to three to four kits by removing excess non-CF kits; this helps to facilitate better care of the CF kits by the jill. At this point, all kits that have passed meconium are then maintained using littermate-matched rearing methods to be described later.
To prevent lung infections, antibiotics are given both at birth and during the rearing. Four antibiotics have been used during the rearing process (metronidazole, Hospira, Inc., Lake Forest, IL; Zosyn, Wyeth Pharmaceuticals Inc., Philadelphia, PA; Baytril, Bayer HealthCare LLC, Shawnee Mission, Kansas; cefazolin, Steri-Pharma, LLC Syrcuse, NY). Typically, metronidazole (5 mg/kg in 100 μL dextrose saline, four times daily s.c.) and Zosyn (4 mg/kg in 100 μL dextrose saline, two times daily, s.c.) are administered at time of birth and tail clipping to all animals in the litter. Metronidazole is given to promote intestinal health and prevent bacterial overgrowth. Zosyn is given to primarily prevent lung infections. Weight gain is monitored every 6 hours and is used to gauge the health of the animals. Typically, a drop in weight gain over a 12 hours period reflects a developing lung infection in CF kits. If this is observed, Baytril (3 mg/kg in 100 μL dextrose saline, two times daily, s.c.) is given in addition to metronidazole and Zosyn. Once animals have been reared for more than 2 weeks, future drops in weight gain are managed by replacing cefazolin (30 mg/kg in 100 μL dextrose saline two times daily, s.c.) for Zosyn. Typically, animals are removed from antibiotics at the time of weaning if the goal of the studies is to evaluate lung infections in the developmentally mature lung.
In addition to antibiotics, nutritional therapies are initiated immediately after CF kits pass stool. Similar to antibiotics, both non-CF and CF littermates are treated identically. Three components are involved in this neonatal nutritional therapy and include gavage feeding with elemental diet supplemented with UDCA and omeprazole. As discussed earlier, UDCA is used to normalize liver function test and potentially improve fat absorption by reducing bile acid wasting often observed in CF patients [77,78]. Omeprazole is used to counteract deficiencies of pancreatic and intestinal bicarbonate production that reduces the pH of the small intestine and leads to inactivation of pancreatic enzymes, precipitation of bile salts, and reduced fat absorption [79–81]. Omeprazole is a proton-pump inhibitor used to enhance pancreatic enzyme function and improve fat absorption in CF patients with residual steatorrhea [82]. UDCA (5 mg/kg) and omeprazole (5 mg/kg) are administered to newborn kits by gavage of 150–200 μL Elecare (Abbott Laboratories, Columbus, OH) supplemented with 10% Golytely four times daily. The percentage of Golytely may also adjusted up to 20% based on the clinical appearance of abdominal bloating and/or the reduced ability of CF kits to pass stool. If intestinal obstruction is suspected, animals are gavaged once with 150–200 μL of 100% Golytely to promote defecation. This can be repeated as needed throughout the rearing process.
As CF ferrets approach the age of weaning (typically 5–6 weeks of age), they require supplementation with pancreatic enzyme to maintain weight gain. This is due to the progressive destruction of the exocrine pancreas. Prior to weaning, pancreatic enzymes are placed into the Elecare fed to kits beginning at 3–4 weeks of age. After weaning, CF and non-CF controls are fed a slurry containing Elecare hydrated solid chow (Marshall Farms) twice daily, mixed with 6 lipase unit of Viokase-V (Neogen Corporation Lexington, KY) per 10 g of body weight. The amount of food eaten with each feeding is recorded for these calculations. The dosing of enzymes needed for adequate weight gain can be adjusted up or down depending on the clinical status of the CF animals. Using these rearing methods, the life expectancy of CF ferrets prior to terminal lung disease ranges from 3 to 8 months.
Summary
New approaches for genetic engineering in larger animals using SCNT have brought the domestic ferret to a new level in biomedical research. The generation of knockout, knock-in, and transgenic ferrets is now feasible. The CF ferret model is the first demonstration of genetic engineering in ferrets, and this model will help to elucidate the pathogenesis of CF and to develop new mechanistic-based therapies. The unique utility of the ferret to model human influenza infection is another future area of biomedical research that may benefit from these new genetic engineering technologies.
Acknowledgments
This work was supported by NIH grants DK047967, HL108902, DK091211, DK096518, and DK54759 to JFE.
References
1. Lindeberg H (2008) Reproduction of the female ferret (Mustela putorius furo). Reprod Domest Anim 43(Suppl. 2): 150–156.
2. Donovan BT (1967) Light and the control of the oestrous cycle in the ferret. J Endocrinol 39: 105–113.
3. Ryan KD, Robinson SL (1987) A study of spontaneous sexual maturation of the female ferret. Biol Reprod 36: 333–339.
4. Hammond JW, Walton A (1934) Notes on ovulation and fertilization in the ferret. J Exp Biol 14: 307–319.
5. Murphy BD (1989) Reproductive physiology of female mustelids. In: Seal US, Thorne ET, Bogan MA, Anderson SH, eds. Conservation biology and the black-footted ferret. New Haven, CN: Yale University Press., pp. 107–123.
6. Carroll RS, Erskine MS, Doherty PC, Lundell LA, Baum MJ (1985) Coital stimuli controlling luteinizing hormone secretion and ovulation in the female ferret. Biol Reprod 32: 925–933.
7. Sun X, Li Z, Yi Y, Chen J, Leno GH, et al. (2008) Efficient term development of vitrified ferret embryos using a novel pipette chamber technique. Biol Reprod 79: 832–840.
8. Chang MC, Yanagimachi R (1963) Fertilization of ferret ova by deposition of epididymal sperm into the ovarian capsule with special reference to the fertilizable lie of ova and the capacitation of sperm. J Exp Zool 154: 175–187.
9. Chang MC (1969) Development of transferred ferret eggs in relation to the age of corpora lutea. J Exp Zool 171: 459–464.
10. Chang MC (1965) Fertilizing life of ferret sperm in the female tract. J Exp Zool 158: 87–99.
11. Kidder JD, Giles JR, Foote RH, Richmond ME, Salerno M (1999) Allocation of inner cell mass and trophectoderm cells to the preimplantation blastocyst of the domestic ferret, Mustela putorius furo. J Exp Zool 283: 202–209.
12. Whittingham DG (1975) Fertilisation, early development, and storage of mammalian ova in vitro. In: Balls M, Wild AE, eds. The early development of mammals: the second symposium of the British Society for Developmental Biology. Cambridge; New York: Cambridge University Press, p. 1–24.
13. Li Z, Jiang QS, Zhang YL, Liu XM, Engelhardt JF (2001) Successful production of offspring after superovulation and in vitro culture of embryos from domestic ferrets (Mustela putorius furos). Reproduction 122: 611–618.
14. Li Z, Jiang Q, Rezaei Sabet M, Zhang Y, Ritchie TC, et al. (2002) Conditions for in vitro maturation and artificial activation of ferret oocytes. Biol Reprod 66: 1380–1386.
15. Lindeberg H, Jarvinen M (2003) Early embryonic development and in vitro culture of in vivo produced embryos in the farmed European polecat (Mustela putorius). Theriogenology 60: 965–975.
16. Piltti K, Aalto J, Jarvinen M, Korhonen H, Kuronen V, et al. (2003) Nuclear maturation of European polecat (Mustela putorius) in vivo oocytes and success in using mustelid hybrids as recipients for embryo transfer. Theriogenology 59: 403.
17. Lindeberg H, Piltti K, Peippo J (2001) In vitro production of farmed European polecat (Mustela putorius) embryos. Theriogenology 55: 429.
18. Chang MC (1968) Reciprocal insemination and egg transfer between ferrets and mink. J Exp Zool 168: 49–59.
19. Kidder JD, Roberts PJ, Simkin ME, Foote RH, Richmond ME (1999) Nonsurgical collection and nonsurgical transfer of preimplantation embryos in the domestic rabbit (Oryctolagus cuniculus) and domestic ferret (Mustela putorius furo). J Reprod Fertil 116: 235–242.
20. Li Z, Sun X, Chen J, Leno GH, Engelhardt JF (2006) Factors affecting the efficiency of embryo transfer in the domestic ferret (Mustela putorius furo). Theriogenology 66: 183–190.
21. Amstislavsky S, Maksimovsky L, Ternovskaya Y, Trenovsky D (1993) Cryopreservation of Carnivora. Mustela erminea. Scientifur 17: 127–131.
22. Lindeberg H, Aalto J, Amstislavsky S, Piltti K, Jarvinen M, et al. (2003) Surgical recovery and successful surgical transfer of conventionally frozen-thawed embryos in the farmed European polecat (Mustela putorius). Theriogenology 60: 1515–1525.
23. Piltti K, Lindeberg H, Aalto J, Korhonen H (2004) Live cubs born after transfer of OPS vitrified-warmed embryos in the farmed European polecat (Mustela putorius). Theriogenology 61: 811–820.
24. Li Z, Sun X, Chen J, Liu X, Wisely SM, et al. (2006) Cloned ferrets produced by somatic cell nuclear transfer. Dev Biol 293: 439–448.
25. Li Z, Chen X, Sun X, Zhou Q, Chen J, et al. (2005) Nuclear transfer of M-phase ferret fibroblasts synchronized with the microtubule inhibitor demecolcine. J Exp Zool A Comp Exp Biol 303: 1126–1134.
26. Li Z, Rezaei Sabet M, Zhou Q, Liu X, Ding W, et al. (2003) Developmental capacity of ferret embryos by nuclear transfer using G0/G1-phase fetal fibroblasts. Biol Reprod 68: 2297–2303.
27. Li Z, Sun X, Chen J, Leno GH, Engelhardt JF (2005) Factors affecting the electrofusion of mouse and ferret oocytes with ferret somatic cells. Mol Reprod Dev 72: 40–47.
28. ErYao W, Yang Y, XueMei L, LiHong J, Liu1 W (2007) Effects of different nuclear transfer and activation methods on the development of mouse somatic cell cloned embryos. Chin Phys Lett 52: 209–214.
29. Wakayama T (2007) Production of cloned mice and ES cells from adult somatic cells by nuclear transfer: how to improve cloning efficiency? J Reprod Dev 53: 13–26.
30. Sun X, Yan Z, Yi Y, Li Z, Lei D, et al. (2008) Adeno-associated virus-targeted disruption of the CFTR gene in cloned ferrets. J Clin Invest 118: 1578–1583.
31. Whyte JJ, Prather RS (2011) Genetic modifications of pigs for medicine and agriculture. Mol Reprod Dev 78: 879–891.
32. Samiec M, Skrzyszowska M (2011) Transgenic mammalian species, generated by somatic cell cloning, in biomedicine, biopharmaceutical industry and human nutrition/dietetics—recent achievements. Pol J Vet Sci 14: 317–328.
33. Robl JM, Wang Z, Kasinathan P, Kuroiwa Y (2007) Transgenic animal production and animal biotechnology. Theriogenology 67: 127–133.
34. Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, et al. (1989) Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245: 1066–1073.
35. Fisher JT, Zhang Y, Engelhardt JF (2011) Comparative biology of cystic fibrosis animal models. Methods Mol Biol 742: 311–334.
37. Liu X, Engelhardt JF (2008) The glandular stem/progenitor cell niche in airway development and repair. Proc Am Thorac Soc 5: 682–688.
38. Liu X, Driskell RR, Engelhardt JF (2006) Stem cells in the lung. Methods Enzymol 419: 285–321.
39. Liu X, Driskell RR, Engelhardt JF (2004) Airway glandular development and stem cells. Curr Top Dev Biol 64: 33–56.
40. Li Z, Engelhardt JF (2003) Progress toward generating a ferret model of cystic fibrosis by somatic cell nuclear transfer. Reprod Biol Endocrinol 1: 83.
41. Liu X, Luo M, Zhang L, Ding W, Yan Z, et al. (2007) Bioelectric properties of chloride channels in human, pig, ferret, and mouse airway epithelia. Am J Respir Cell Mol Biol 36: 313–323.
42. Sun X, Sui H, Fisher JT, Yan Z, Liu X, et al. (2010) Disease phenotype of a ferret CFTR-knockout model of cystic fibrosis. J Clin Invest 120: 3149–3160.
45. Khan IF, Hirata RK, Russell DW (2011) AAV-mediated gene targeting methods for human cells. Nat Protoc 6: 482–501.
46. Inoue N, Hirata RK, Russell DW (1999) High-fidelity correction of mutations at multiple chromosomal positions by adeno-associated virus vectors. J Virol 73: 7376–7380.
47. Liu X, Yan Z, Luo M, Zak R, Li Z, et al. (2004) Targeted correction of single-base-pair mutations with adeno-associated virus vectors under nonselective conditions. J Virol 78: 4165–4175.
48. Fujimura T, Murakami H, Kurome M, Takahagi Y, Shigehisa T, et al. (2008) Effects of recloning on the efficiency of production of alpha 1,3-galactosyltransferase knockout pigs. J Reprod Dev 54: 58–62.
49. Rowe SM, Miller S, Sorscher EJ (2005) Cystic fibrosis. N Engl J Med 352: 1992–2001.
50. Oppenheimer EH (1981) Similarity of the tracheobronchial mucous glands and epithelium in infants with and without cystic fibrosis. Hum Pathol 12: 36–48.
51. Cantin A (1995) Cystic fibrosis lung inflammation: early, sustained, and severe. Am J Respir Crit Care Med 151: 939–941.
52. Zuelzer WW, Newton WA, Jr. (1949) The pathogenesis of fibrocystic disease of the pancreas; a study of 36 cases with special reference to the pulmonary lesions. Pediatrics 4: 53–69.
53. Engelhardt JF, Yankaskas JR, Ernst SA, Yang Y, Marino CR, et al. (1992) Submucosal glands are the predominant site of CFTR expression in the human bronchus. Nat Genet 2: 240–248.
54. Salinas D, Haggie PM, Thiagarajah JR, Song Y, Rosbe K, et al. (2005) Submucosal gland dysfunction as a primary defect in cystic fibrosis. FASEB J 19: 431–433.
55. Sehgal A, Presente A, Engelhardt JF (1996) Developmental expression patterns of CFTR in ferret tracheal surface airway and submucosal gland epithelia. Am J Respir Cell Mol Biol 15: 122–131.
56. Verkman AS, Song Y, Thiagarajah JR (2003) Role of airway surface liquid and submucosal glands in cystic fibrosis lung disease. Am J Physiol Cell Physiol 284: C2–C15.
57. Wine JJ, Joo NS (2004) Submucosal glands and airway defense. Proc Am Thorac Soc 1: 47–53.
58. Oppenheimer EH, Esterly JR (1973) Cystic fibrosis of the pancreas. Morphologic findings in infants with and without diagnostic pancreatic lesions. Arch Pathol 96: 149–154.
59. Durie PR, Forstner GG (1989) Pathophysiology of the exocrine pancreas in cystic fibrosis. J R Soc Med 82(Suppl. 16): 2–10.
60. Olivier AK, Yi Y, Sun X, Sui H, Liang B, et al. (2012) Abnormal endocrine pancreas function at birth in cystic fibrosis ferrets. J Clin Invest 122: 3755–3768.
61. Lohr M, Goertchen P, Nizze H, Gould NS, Gould VE, et al. (1989) Cystic fibrosis associated islet changes may provide a basis for diabetes. An immunocytochemical and morphometrical study. Virchows Arch A Pathol Anat Histopathol 414: 179–185.
62. Moran A, Becker D, Casella SJ, Gottlieb PA, Kirkman MS, et al. (2010) Epidemiology, pathophysiology, and prognostic implications of cystic fibrosis-related diabetes: a technical review. Diabetes Care 33: 2677–2683.
63. Hameed S, Jaffe A, Verge CF (2011) Cystic fibrosis related diabetes (CFRD)—the end stage of progressive insulin deficiency. Pediatr Pulmonol 46: 747–760.
64. van der Doef HP, Kokke FT, van der Ent CK, Houwen RH (2011) Intestinal obstruction syndromes in cystic fibrosis: meconium ileus, distal intestinal obstruction syndrome, and constipation. Curr Gastroenterol Rep 13: 265–270.
65. Romi H, Cohen I, Landau D, Alkrinawi S, Yerushalmi B, et al. (2012) Meconium ileus caused by mutations in GUCY2C, encoding the CFTR-activating guanylate cyclase 2C. Am J Hum Genet 90: 893–899.
66. Dorfman R, Li W, Sun L, Lin F, Wang Y, et al. (2009) Modifier gene study of meconium ileus in cystic fibrosis: statistical considerations and gene mapping results. Hum Genet 126: 763–778.
67. Herrmann U, Dockter G, Lammert F (2010) Cystic fibrosis-associated liver disease. Best Pract Res Clin Gastroenterol 24: 585–592.
68. Oppenheimer EH, Esterly JR (1975) Hepatic changes in young infants with cystic fibrosis: possible relation to focal biliary cirrhosis. J Pediatr 86: 683–689.
69. Lindblad A, Glaumann H, Strandvik B (1999) Natural history of liver disease in cystic fibrosis. Hepatology 30: 1151–1158.
70. Scher H, Bishop WP, McCray PB, Jr. (1997) Ursodeoxycholic acid improves cholestasis in infants with cystic fibrosis. Ann Pharmacother 31: 1003–1005.
71. Lepage G, Paradis K, Lacaille F, Senechal L, Ronco N, et al. (1997) Ursodeoxycholic acid improves the hepatic metabolism of essential fatty acids and retinol in children with cystic fibrosis. J Pediatr 130: 52–58.
72. Matel JL, Milla CE (2009) Nutrition in cystic fibrosis. Semin Respir Crit Care Med 30: 579–586.
73. Milla CE (2004) Association of nutritional status and pulmonary function in children with cystic fibrosis. Curr Opin Pulm Med 10: 505–509.
74. Koletzko S, Reinhardt D (2001) Nutritional challenges of infants with cystic fibrosis. Early Hum Dev 65(Suppl.): S53–S61.
75. Jarzabek K, Zbucka M, Pepinski W, Szamatowicz J, Domitrz J, et al. (2004) Cystic fibrosis as a cause of infertility. Reprod Biol 4: 119–129.
76. Oppenheimer EH, Esterly JR (1975) Pathology of cystic fibrosis review of the literature and comparison with 146 autopsied cases. Perspect Pediatr Pathol 2: 241–278.
77. O'Brien S, Mulcahy H, Fenlon H, O'Broin A, Casey M, et al. (1993) Intestinal bile acid malabsorption in cystic fibrosis. Gut 34: 1137–1141.
78. Fondacaro JD, Heubi JE, Kellogg FW (1982) Intestinal bile acid malabsorption in cystic fibrosis: a primary mucosal cell defect. Pediatr Res 16: 494–498.
79. Robinson PJ, Smith AL, Sly PD (1990) Duodenal pH in cystic fibrosis and its relationship to fat malabsorption. Dig Dis Sci 35: 1299–1304.
81. Geus WP, Gan KH, Lamers CW, Heijerman HM (1999) Simultaneously measured duodenal and gastric pH in patients with cystic fibrosis: effect of omeprazole. Neth J Med 54(Suppl.): 63.
82. Proesmans M, De Boeck K (2003) Omeprazole, a proton pump inhibitor, improves residual steatorrhoea in cystic fibrosis patients treated with high dose pancreatic enzymes. Eur J Pediatr 162: 760–763.