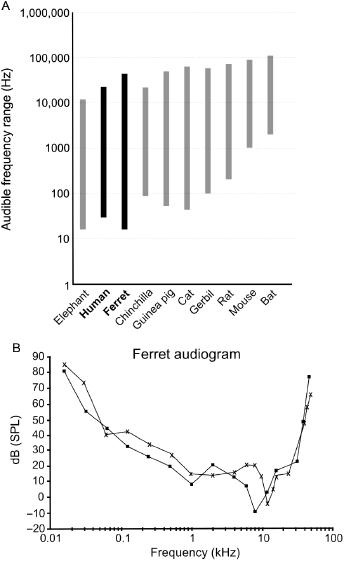
In recent years, ferrets have become increasingly popular as an animal model for studying hearing, and are now used for this purpose in a number of laboratories around the world. There are several reasons for this. First, as with their ability to see, the functional onset of hearing takes place in ferrets several weeks after birth, allowing postnatal access to an immature nervous system at a stage that can only be studied in utero in some other more precocial species. Second, the range of sound frequencies that are audible to ferrets (approximately 20 Hz–44 kHz) entirely overlaps, and extends beyond, that of humans (Fig. 29.1). In contrast to many rodents, particularly mice, whose audible range is shifted to higher frequencies, the sensitivity of ferrets to low-frequency sounds allows the study of various aspects of hearing, including pitch perception, sound localization, and the processing of communication calls, within a similar frequency range to that used by humans. Finally, because of their inquisitive nature and intelligence, ferrets can be readily trained to carry out a variety of auditory behavioral tasks, which has led to the development of paradigms for measuring their ability to detect, discriminate, and localize sounds. Moreover, for some aspects of hearing, their performance in these tasks closely resembles that of human listeners, providing an impetus for using ferrets to investigate the neural basis of auditory perception as well as the effects of hearing loss and its restoration.
Fig. 29.1. Hearing range and audiogram of the ferret. (A) The audible frequency range of different mammals, including humans, ferrets, and some of the most common species used for auditory research. Note the total overlap of the ferret's hearing range with that of humans, which makes ferrets a more suitable species in which to study the processing of low-frequency vocalizations and sound localization cues than rodents, whose vocalizations sometimes lie beyond the range of human hearing. (B) Typical audiogram of two ferrets, whose audible frequency range, as indicated in panel A, extends from 20 Hz to about 40 kHz with maximum sensitivity around 7–11 kHz. Reproduced with permission from Kelly et al. [1].
Behavioral Studies of Hearing in Ferrets
The first auditory behavioral study in ferrets used a positive conditioning paradigm to measure the range and sensitivity of the animals to tones of different frequency [1]. They were trained to initiate a trial by touching a spout positioned in front of a central start platform and to approach a peripheral water spout located at either 30° to the right when they heard a sound or to the left when they heard no sound. Correct responses were reinforced by the delivery of a water reward. The same principle was then adopted to measure minimum audible angles (MAAs), that is, the ability of ferrets to discriminate between sounds presented from one of two speakers separated by progressively smaller angles in the horizontal plane [2,3]. Subsequent studies expanded this to a 12-speaker task, measuring both the accuracy of the approach to target responses and of the initial head orienting responses made following sound presentation [4]. Ferrets are very easy to train in this type of task and it has even proved possible, using a left–right response paradigm, to measure MAAs in the vertical plane [5,6].
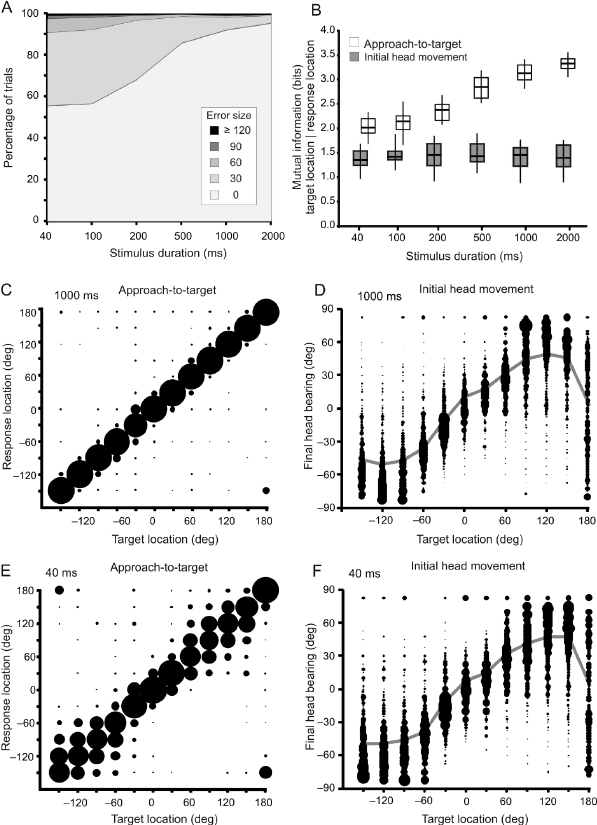
The horizontal plane MAAs measured in ferrets are comparable with those reported for other carnivores [7]. Their spatial acuity varies with the location of the stimulus from ∼10° to 15° when animals are tested with brief noise bursts around the midline to ∼25–30° at more lateral locations [3,8]. This spatial dependence in localization accuracy is also seen when ferrets are trained to approach the source of 1 of 12 loudspeakers arranged at 30° intervals in the horizontal plane [4]. The accuracy of approach-to-target responses declines as the duration of the stimulus is reduced (Fig. 29.2A,B), particularly for lateral and posterior target locations, although responses to sounds presented in the frontal region of space and directly behind the animal remain largely unchanged (Fig. 29.2C,E). Sound-evoked head movements made by the animal before it leaves the central start platform have a latency of ∼200 milliseconds and vary systematically in magnitude with the direction of the stimulus (Fig. 29.2D,F). In contrast to the approach-to-target behavior, the accuracy of the head orienting responses is less affected by changes in stimulus duration (Fig. 29.2B), suggesting that these are triggered by the onset of the sound and that the improved performance observed when animals are approaching longer duration sounds may, at least in part, be due to resampling of the stimulus after the initial head turn has been made.
Fig. 29.2. Sound localization behavior in ferrets. (A) Overall percentage across 12, equally spaced target locations of trials classified by their error size for different durations of a broadband stimulus (from 2000 to 40 milliseconds). The incidence of incorrect trials and the size of the errors increased as the stimulus duration was reduced. (B) The behavioral responses were quantified by calculating the mutual information (MI) between target location and response location. The MI values (in bits) are shown for both the initial head orienting and the subsequent approach-to-target responses at different stimulus durations. The MI between target and response location increased with stimulus duration for the approach-to-target task, indicating that the animals became more accurate, but remained constant for the relationship between final head bearing and target location. The boxes represent the interquartile range, the horizontal line is the median, and the vertical line is the full range of MI values. (C, E) Stimulus–response plots showing the distribution of approach-to-target responses for two stimulus durations, (C) 1000 and (E) 40 milliseconds. For each target location, the size of the dots is proportional to the probability of responses to different target locations. The corresponding percent correct and MI values for these stimulus durations (1000 and 40 milliseconds) are shown in panels A and B, respectively. (D, F) Stimulus–response plots showing the distribution of final head bearings (binned in steps of 7.5°) for each target location. The size of the dots is proportional to the response probability. The gray lines represent the mean final bearing for each target location. Reproduced with permission from Nodal et al. [78].
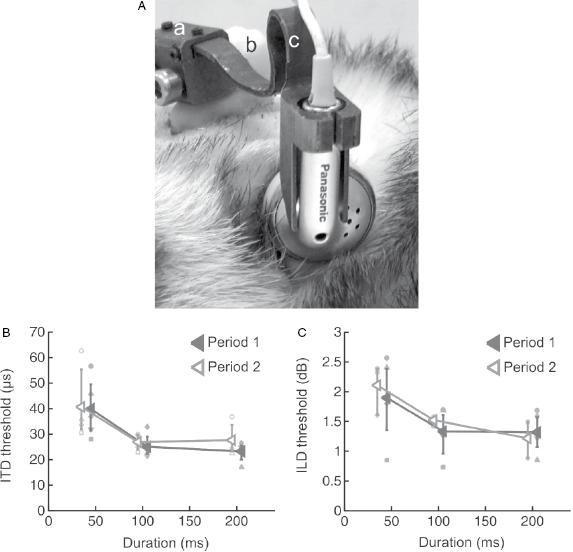
The ability of animals to localize sound depends on the geometry of the head and external ears [9]. Because of the physical separation of the ears on either side of the head, sounds originating from one side will arrive at the closer ear first. Depending on their wavelength, they may also be attenuated at the far ear as a result of the acoustic shadow cast by the head, while interactions with the folds of the external ears can modify the spectral shape of the sound. Localization of a sound source therefore relies on the sensitivity of the auditory system to a combination of interaural time differences (ITDs), interaural level differences (ILDs), and spectral cues. In order to understand the basis for localization behavior in ferrets, closed-field techniques have been developed to measure their sensitivity to binaural cues [10]. This involves attaching earphones to lightweight titanium holders that are mounted on the animal's head, so that they can be easily disconnected and consistently repositioned just in front of the ear canals (Fig. 29.3A). Ferrets are extremely sensitive to both ITDs and ILDs (Fig. 29.3B,C), with thresholds that can largely account for their ability to localize stimuli in the horizontal plane [11,12]. Indeed, their behavioral sensitivity to these cues closely resembles that found in other species, including humans, confirming the suitability of ferrets as a model for studying spatial hearing.
Fig. 29.3. Measuring sensitivity to binaural localization cues in ferrets. (A) Photograph of the head of a ferret wearing lightweight titanium headphone holders attached to a chronic implant on the skull, which enable earphones to be positioned close to the entrance of the ear canal. Reproduced from Nodal et al. [10]. (B) Mean interaural time difference (ITD) thresholds across different animals are shown for noise bursts as a function of stimulus duration. Data for individual animals are denoted by symbols of the same color and are shown for two testing sessions (indicated by the different lines) separated by 16 weeks to illustrate the stability of these measurements. (C) Corresponding data for interaural level difference (ILD) thresholds obtained from the same ferrets. Data from Keating et al. [11].
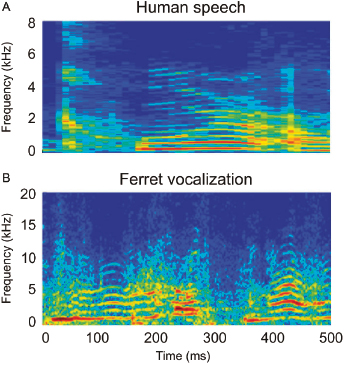
Ferrets are also used to investigate aspects of sound identification. Although little is currently known about how they perceive their own vocalizations (Fig. 29.4), their ability to discriminate the periodicity and timbre of artificial vowels that cover the same frequency range has been measured. Vowel identification involves picking out the formant peaks in the spectral envelope of the sound, and is therefore a timbre discrimination task. Ferrets readily learn to distinguish different vowels and, like humans, are able to generalize across a range of stimulus periodicities and to maintain their behavior in the presence of background noise [13].
Fig. 29.4. Comparison of human and ferret vocalizations. Spectrograms showing examples of human speech (A) and ferret vocalizations (B). In each case, the sound is produced by the repeated opening and closing of the vocal folds, resulting in glottal pulse trains that are made up of a fundamental frequency and a number of higher harmonics. Transmission of these pulse trains through the resonant cavities of the vocal tract then enhances some harmonics and suppresses others. The resulting frequency structure is visible as roughly horizontal bands in the spectrograms. Note the difference in the scale of the y-axis in the two panels: the ferret calls contain higher frequency components than human speech, reflecting differences in the size and structure of the vocal apparatus between these species.
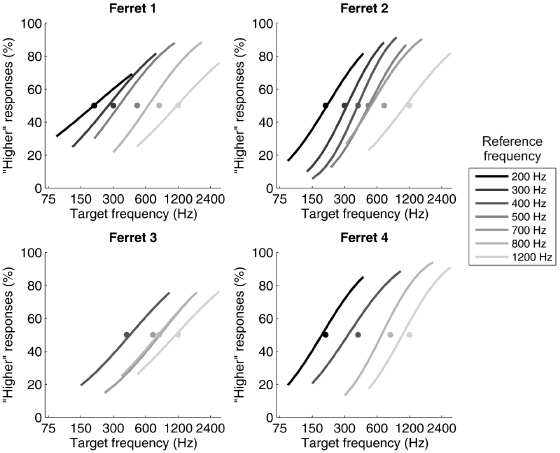
The periodicity of a sound corresponds to its perceived pitch and conveys information about speaker identity (males tend to have lower pitch voices than females), age, and emotional state. Because pitch is a perceptual attribute and is not related in a straightforward way to the physical properties of sound, it is hard to demonstrate conclusively that other species experience pitch in the same way as humans. Nevertheless, there is now plenty of evidence that nonhuman animals are sensitive to the same periodicity cues as humans [14]. For example, ferrets are sometimes able to spontaneously distinguish harmonic complex tones, which, in humans, have a clear pitch associated with them, from inharmonic tones [15], and can be trained to discriminate the relative pitch of pairs of tones that either rise or fall in frequency [16]. Walker et al. [17] showed that ferrets can be trained to discriminate two consecutive artificial vowels by indicating whether the pitch of the second sound was higher or lower than that of the first (Fig. 29.5). In this case, however, their thresholds were much poorer than those found in humans and other species. Moreover, the animals appeared to perform the two-alternative forced choice task by attending to the absolute pitch height of the second sound, rather than the direction of the pitch change between the two vowels. This does not mean, however, that ferrets are unsuitable species to use for studying pitch perception, since much lower thresholds have been found when they perform a go/no-go change detection task.
Fig. 29.5. Pitch discrimination by ferrets. The animals performed a two-alternative forced choice task, in which they indicated by approaching one of two reward spouts whether a target sound was higher or lower in pitch than a preceding reference sound. Each panel shows the psychometric functions for individual animals. The grayscale of each curve corresponds to the fundamental frequency of the reference vowel, as indicated to the right. The different reference values are represented by gray circles at 50% choice probability. Reprinted, with permission, from Walker et al. [17].
Finally, auditory temporal processing has been examined behaviorally in ferrets by measuring their thresholds for detecting short gaps in a sound [18], while other studies have tested their hearing abilities in more complex acoustic environments [19–22]. This work has shown that, like humans, ferrets use binaural cues to help them pick out sounds against other sounds originating from different directions in space [19], can suppress echoes when localizing sounds [21], and appear to be able to perceptually organize sound sequences into auditory streams [20].
Organization of the Auditory System
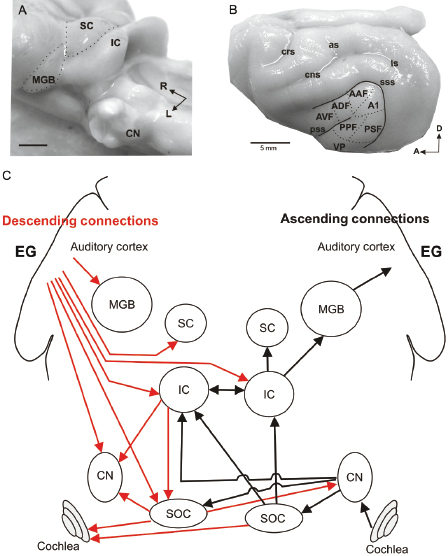
The anatomy of the ear and the organization of the central auditory pathway of the ferret follow the same principles that have been described in other mammals. Although relatively little research has been carried out on the middle ear or inner ear in this species, other than in the context of studying the effects of upper respiratory infections (e.g., Ref. 23), a great deal is known about how its immobile external ears modify incoming sounds to provide spectral cues for source location [24–26]. All the auditory nuclei or relay stations in the brain, which have been characterized in numerous mammalian species (for a review of the neuroanatomy of the auditory system, see Ref. 27), are well developed and easily distinguishable in ferrets (Fig. 29.6). Thus, after leaving the cochlea, the hearing part of the inner ear, the auditory nerve fibers enter the cochlear nucleus (CN) in the brainstem [28,29], which in turn sends bilateral projections to the superior olivary complex (SOC) [30]. This is where convergence of information from both ears is registered for the first time. Both the CN and SOC project to the inferior colliculus (IC) in the midbrain for further signal processing [28,31]. The IC and other brainstem areas provide the principal auditory input to the superior colliculus (SC) [32,33], another midbrain nucleus. The IC also projects to the medial geniculate body or auditory thalamus [34], which constitutes the gateway to the auditory cortex located in the ectosylvian gyrus (EG) (Fig. 29.6B,C). Some species-specific differences have been noted in these anatomical studies, such as the proportion of ipsi versus contralateral projections from the lateral superior olive to the IC in ferrets and cats [28]. Overall, however, the connectivity between the different auditory regions of the ferret's brain is broadly consistent with that found in other species.
Fig. 29.6. Organization of the auditory pathway in the ferret. (A) Photograph of the brainstem of a ferret after removing the cerebellum and cerebral hemispheres to expose most of the subcortical auditory relay stations. (B) Photograph of the ferret cortex in which the location of different auditory fields on the ectosylvian gyrus (EG) is indicated. (C) Simplified diagram depicting the main ascending (black) and descending (red) connections of the auditory pathway. For clarity, all lines have been drawn with the same thickness independent of the size of the projection they represent. Each of the connections is shown on one side of the brain only. AAF, anterior auditory field; A1, primary auditory field; ADF, anterior dorsal field; as, ansinate sulcus; AVF, anterior ventral field; CN, cochlear nuclei; cns, coronal sulcus; crs, cruciate sulcus; IC, inferior colliculus; SC, superior colliculus; MGB, medial geniculate body; ls, lateral sulcus; PPF, posterior pseudosylvian field; PSF, posterior suprasylvian field; pss, pseudosylvian sulcus; SOC, superior olivary complex; sss, suprasylvian sulcus; VP, ventro posterior field.
In addition to the afferent connections from cochlea to cortex, the auditory pathway is characterized by extensive efferent or descending connections (Fig. 29.6C), the size of which often outweighs those carrying information in the ascending auditory pathway. The functions of these descending pathways remain unclear, but they are thought to modulate the processing of acoustical signals at lower levels in the pathway [35]. As in other species, descending projections in ferrets link the auditory cortex with the IC [36] and SC [37], and the SOC with the cochlea [38]. As we shall discuss later in this chapter, by combining methods for selectively lesioning these pathways with behavioral testing, studies in ferrets have provided the first functional evidence for the role that descending connections might play in hearing [38,39].
Physiological Studies of Auditory Processing in Ferrets
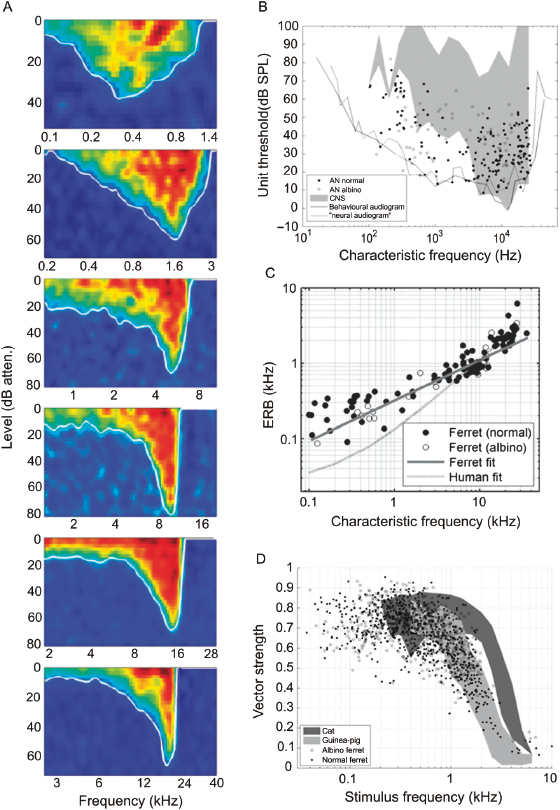
The functional organization of the central auditory pathways has been studied by measuring the responses of neurons in each region to a variety of sounds (reviewed in Refs. 9 and 40). The animal species most commonly used are guinea pigs, gerbils, and cats, with macaque monkeys and marmosets being the subject of many of the studies on auditory cortical processing. The use of ferrets in this field dates back to the 1980s [41,42], and has focused mainly on the midbrain and cortex. Indeed, the only investigation so far of the response properties of neurons at more peripheral levels was carried out by Sumner and Palmer [43], who described the sound frequency tuning, spontaneous firing rate, relationship between firing rate and sound level, and phase locking of auditory nerve fibers (Fig. 29.7). To a large extent, the coding of information about sound frequency and level by auditory nerve fibers reflects the properties of the cochlear inner hair cells to which they are connected. Perhaps surprisingly, Sumner and Palmer [43] found that the auditory nerve in the ferret more closely resembles that of the guinea pig or chinchilla than that of the cat.
Fig. 29.7. Ferret auditory nerve physiology. (A) Frequency response areas from six auditory nerve fibers in one animal with characteristic frequencies (CFs) ranging from below 0.6 to above 17 kHz. Sound level is expressed in decibel attenuation, where 0 dB attenuation is ∼100 dB sound pressure level (SPL). (B) Minimum thresholds as a function of CF. The black continuous line is the ferret audiogram [1]. The gray area is the outline of minimum thresholds reported in the IC and auditory cortex [41,42,106]. The dashed line is the outline when the frequency tuning curves from all of the fibers are superimposed. AN, auditory nerve; CNS, central nervous system. (C) Equivalent rectangular bandwidth (ERB), a measure of frequency selectivity, versus CF. The solid line shows the fit ERBkHz = 0.31CFkHz0.533. The dashed line shows a fit to human psychophysical data [ERBkHz = 0.0247 (0.00437CFkHz + 1)] [107]. (D) Phase locking as a function of stimulus frequency. Each point represents a significant vector strength value for a single stimulus frequency in a single fiber. Also shown are the range of values reported by Palmer and Russell [108] in the guinea pig (light gray) and by Johnson [109] in the cat (dark gray). Reproduced, with permission, from Sumner and Palmer [43].
Several studies have examined the response properties of neurons in the IC of the ferret (e.g., Refs. 41, 44, and 45), sometimes as part of a wider investigation of how the representation of natural sounds changes at higher levels of the auditory system [46–48]. This species has also been used extensively for investing the auditory response properties of neurons in the SC [49]. Unlike other parts of the auditory pathway, the spatial receptive fields of SC neurons are organized topographically to form a map of auditory space, which lies in spatial register with the representations of other sensory modalities. This arrangement allows the different sensory signals associated with a particular object to be registered at the same location, and often by the same neurons, within the SC, so that they can trigger orienting movements that help to redirect attention toward the object [50]. Although studies of how multisensory inputs interact to determine the responses of SC neurons have been carried out primarily in other species [51], experiments in ferrets have helped to reveal how a map of auditory space is constructed in the brain from the localization cues generated by the way sounds interact with the head and external ears [25,52,53].
In recent years, interest in ferrets as a model for auditory cortical processing has greatly increased. A common feature of all sensory systems is that they comprise multiple cortical areas that can be defined both physiologically and anatomically, and which are collectively involved in the processing of the world around us. As in other mammalian species, a number of different auditory cortical fields with distinct functional properties are found in ferrets [54–56] (Fig. 29.6B). The middle portion of the EG comprises the primary auditory cortex (A1) and the anterior auditory field (AAF): the neurons found there have short latency responses that are most sensitive to particular sound frequencies, which vary systemically in value with neuron location within each cortical area to form “tonotopic” maps. There is little doubt that an equivalent area to the region designated as A1 is found in many different mammalian species, including humans, and the same is probably true of AAF. Drawing homologies between the other fields described in the ferret auditory cortex and those described in other species has, however, proved to be less straightforward. Two more tonotopically organized, secondary cortical areas are found on the posterior EG, and the neurons found there can be distinguished from those in the primary areas by the temporal characteristics of their responses. Finally, two further fields have been described in the anterior EG, where neurons respond to sound but lack the tonotopic order that is characteristic of the rest of auditory cortex.
Each of the auditory fields in the ferret cortex has also been shown to receive visual inputs, which are likely to arise from different parts of the visual cortex [57]. In fact, the region labeled in Fig. 29.6B as the anterior ventral field (AVF) is probably more accurately designated a higher-level multisensory area, since it contains a particularly high incidence of visually responsive neurons and lies close to a somatosensory field [58]. Other reports have also highlighted the multisensory nature of neurons in the anterior EG [59,60]. Inputs from other sensory modalities to auditory cortex have been observed in a growing number of other species too, including primates [61]. Although their functions are not well understood, they appear to enhance the capacity of cortical neurons to signal the location [62] or identity [63,64] of a sound source, and are therefore likely to contribute to the well known effects of vision on auditory perception.
Recording studies in ferrets have provided novel insights into the way in which various sound attributes, including periodicity, timbre, and spatial location are represented in the cortex [65–67], and have also contributed to our understanding of how the brain encodes natural sounds such as speech [47,68,69]. Using the same artificial vowel sounds used to measure pitch and timbre perception in the behavioral studies described in an earlier section of this chapter, Bizley et al. [66] found that sensitivity to both spatial and nonspatial sound attributes is widely distributed across different cortical fields. Regional differences do exist, however, with the firing patterns of neurons in certain areas being more informative about the periodicity or timbre of the sound, whereas those in other areas are more sensitive to its location in space. This is broadly consistent with evidence in other species for distinct cortical processing streams for different aspects of the auditory scene [70,71]. At the same time, the ferret recordings have revealed that these stimulus attributes are not only coded by different spatial patterns of activation but also by different temporal patterns within the same population [67]. In other words, the firing rate of a neuron can be modulated by the identity (i.e., spectral timbre) of an artificial vowel sound in one time window and the periodicity of the vowel in a later time window. This is known as “multiplexing” and allows the neurons to carry mutually invariant information about these two perceptual features. In keeping with this neurophysiological finding, ferrets were found to detect changes in vowel identity faster than they detected changes in the periodicity of these sounds [67]. This cortical processing strategy potentially allows for a more dynamic representation of perceptual attributes or even the acquisition of representations of new attributes that become relevant or are learned through behavioral training.
The primary reason for exploring the response properties of neurons in the brain is to attempt to identify the neural basis for perception. One approach to this is to make quantitative comparisons of the psychometric performance of an animal in a given detection or discrimination task with “neurometric” measures of how well single neurons or neural ensembles can detect a change in the stimulus. Using this approach, Bizley et al. [72] found that relatively small networks of neurons throughout the auditory cortex of anesthetized ferrets can account for the ability of trained animals to detect the direction of a pitch change. This appears to be at odds with studies in other species that have reported the existence of a localized pitch center in the cortex, where neurons are tuned to different periodicities [73]. This remains controversial, however, and establishing a more direct link requires making simultaneous behavioral and electrophysiological recordings from the same subject. This approach has mainly been restricted to larger animals, particularly nonhuman primates, and has rarely been used in the auditory system. Recently, however, it has been demonstrated in ferrets trained to perform a pitch discrimination task that auditory cortical activity represents not only the periodicity of the sound but also the pitch that the animals appear to hear as assessed by their behavioral responses [74].
In order to demonstrate that a given area of the brain is involved in a particular aspect of perception, it is also necessary to show that silencing or manipulating neural activity in that area alters the animal's behavior. A commonly used approach is to lesion the brain region in question, which, in ferrets, has been used to show that the SOC [75], IC [8], and auditory cortex [2,76] are all necessary for accurate sound localization. These results support the widely accepted idea that although the different cues for sound localization are extracted and computed in the brainstem, higher-level areas, and the auditory cortex, in particular, play a critical role in determining the location of a sound source. A reversible pharmacological inactivation method that utilizes subdural placement of a polymer to release the GABAA agonist muscimol over a period of weeks has been developed in ferrets [77], and used to confirm that both primary- and higher-level auditory fields contribute to localization accuracy when animals make their responses by approaching the source of the sound for a fluid reward [77,78]. The accuracy of the initial head orienting response is preserved, however, following aspiration lesions [76] or pharmacological inactivation [77] of A1, suggesting that the neural circuitry involved in mediating these responses may be different.
Development and Plasticity of the Auditory System
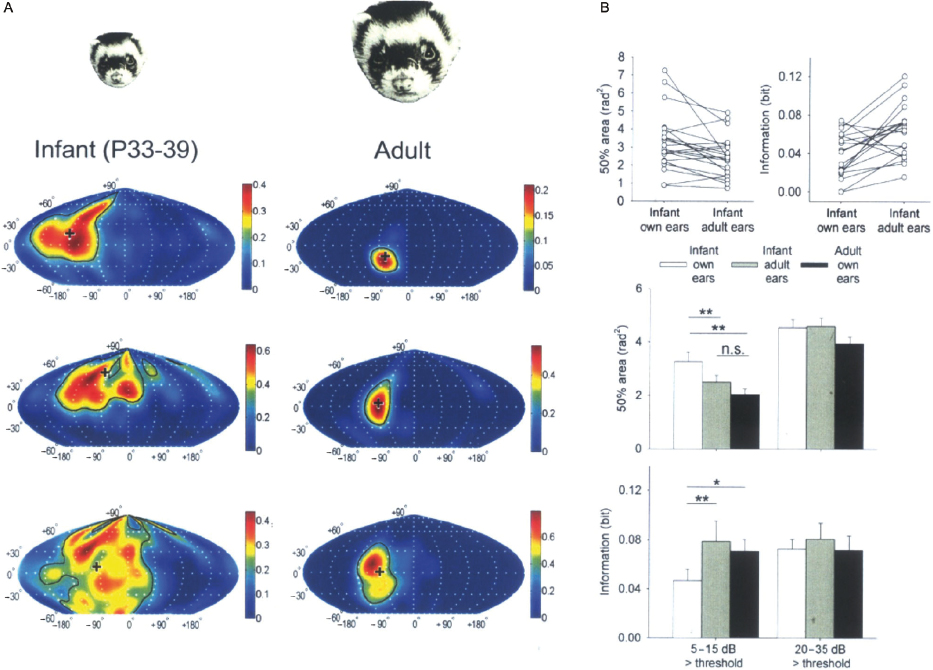
Interest in the use of ferrets for studying the auditory system was driven initially by their suitability for developmental studies. Compared with most other mammals, ferrets are particularly immature at birth and do not begin to hear until near the end of the first postnatal month [79]. Despite their relative immaturity, young ferrets are robust and highly suitable for electrophysiological recording experiments. Several studies have examined the maturation of auditory circuits at different ages [33,80], or have documented how the response properties of auditory neurons change following hearing onset. For example, recordings from the SC [81] and auditory cortex [82] have shown that the spatial tuning of the neurons is much broader in young ferrets than it is in adults. It turns out this can be explained by changes in the localization cue values that take place as the head and ears grow [83]. Thus, presenting infant ferrets with stimuli via earphones that replicate how sounds are filtered by the head and ears of adult animals led to an immediate sharpening of the spatial receptive fields (Fig. 29.8). At the same time, there is considerable evidence that central auditory circuits undergo experience-dependent changes during development and studies in ferrets have provided valuable insights into this process.
Fig. 29.8. Identifying the factors that shape the maturation of the spatial sensitivity of neurons in the auditory cortex of the ferret. (A) Representative examples of spatial receptive fields recorded at a sound level of 10 dB above neuronal threshold for different neurons in the auditory cortex of infant and adult ferrets. These plots illustrate the region of space within which a sound can modulate the firing rate of the neurons. The receptive fields recorded from infant cortex (P33–P39; left column) are more irregular and larger than those recorded in adults (right column). The cross indicates the direction of the spatial response field (SRF) centroid vector. The color bar denotes the evoked spike count per stimulus presentation, averaged over five repetitions. The black contour line runs along the half-maximal response level, demarcating the 50% response area. (B) Pair-wise comparisons of near-threshold 50% area and number of bits of information transmitted between the stimulus location and response for spatial receptive fields recorded from infant cortex through the animals' own ears (“infant own ears”) and then through virtual adult ears (“infant adult ears”). Note that the 50% areas become smaller, and the transmitted information increases when mature localization cue values are provided. The histograms show the mean 50% area and information values derived from infant (own-ear and adult-ear) SRFs and adult (own-ear) SRFs recorded at near threshold (5–15 dB above unit threshold) and at higher sound levels (20–35 dB above threshold; n = 40) (n.s., not significant; *P < 0.05; **P < 0.01). Error bars indicate S.E.M. Adapted from Mrsic-Flogel et al. [82].
One commonly used approach for investigating the plasticity of auditory circuits in the brain is to change the balance of inputs from the two ears, either by ablating the cochlea or by introducing a reversible conductive hearing loss or some other manipulation on one side. Unilateral cochlear removal in ferrets results in an increase in the number of neurons that project from the CN on the nonoperated side to the ipsilateral IC [84], and an increase in the proportion of IC neurons that are excited by the intact ear [85]. This anatomical reorganization is also seen when ferrets are raised with one ear occluded by an earplug [86].
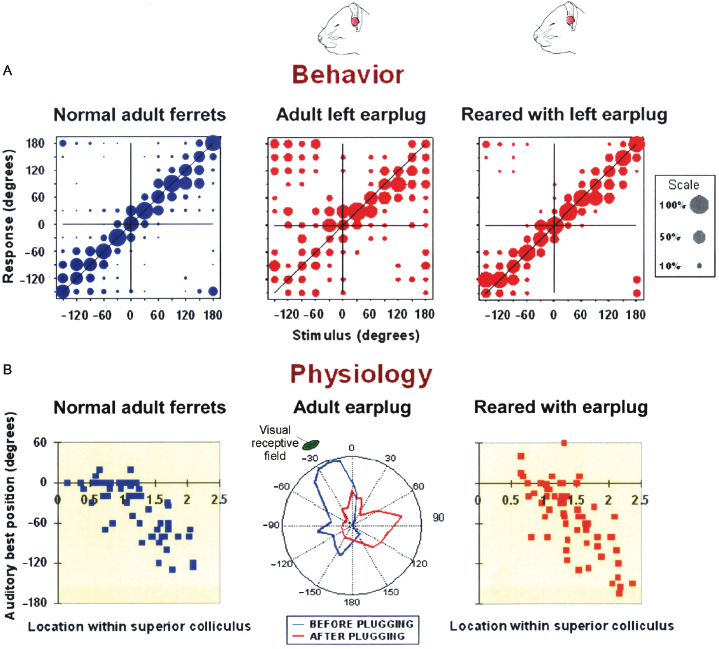
Although comparable results have been obtained in other species, these studies have been extended in ferrets to examine the functional consequences of hearing loss at different ages. Plugging one ear, either in infancy or adulthood, results in an impairment in binaural hearing when assessed by measuring the ability of ferrets to detect a tone in the presence of noise originating from other directions, with binaural unmasking gradually recovering following earplug removal [87,88] (Fig. 29.9). While these findings provide little indication of any adaptation to the partial loss of hearing in one ear, a different result was obtained when the ability of adult ferrets that had been raised with a unilateral earplug was measured [26,88]. Even though the earplug was still in place, these animals were able to localize sounds accurately (Fig. 29.10A) and the auditory spatial tuning of neurons in the SC adjusted so that a near-normal map of auditory space emerged (Fig. 29.10B). This adaptive plasticity does not involve a retuning of neurons to the altered binaural cues, but is instead brought about by developing a greater dependence on the unchanged spectral localization cues provided by the intact ear [26].
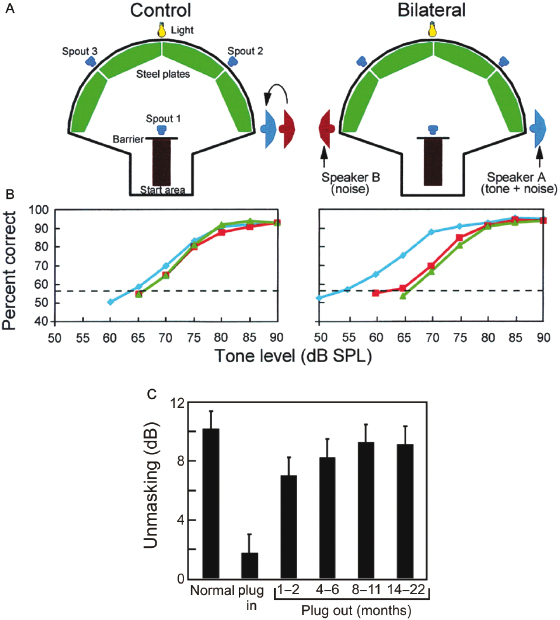
Fig. 29.9. Effect of abnormal auditory experience on the development of binaural unmasking in ferrets. (A) Adult ferrets can be trained to detect a 500-Hz tone (delivered from speaker A) in the presence of a noise (delivered from speakers A and B) when a light flashed. The noise stimuli were presented continuously. On 50% of trials, a tone was presented when the ferret contacted spout 1. Success in the task was measured by the ferret correctly identifying the presence or absence of the tone (by going to spouts 2 or 3, respectively). In the control condition (tones and noise interaurally in phase), both speakers were positioned on the ferret's right side. In the bilateral condition, the noise was made out of phase with the tone by moving speaker B (noise alone) to the ferret's left side. The difference in tone threshold between the two conditions measured binaural unmasking. Unilateral or asymmetric conductive hearing loss, as seen in middle ear disease in children and otosclerosis in adults, can lead to poor binaural hearing. In the laboratory, these diseases are modeled using earplugs in experimental animals to control for variability and to investigate mechanisms. (B) Mean psychometric functions from three groups of ferrets (blue, normally hearing animals; red, animals reared with a unilateral conductive hearing loss and tested as adults; green, animals with a unilateral conductive hearing loss that began in adulthood). Threshold (56.5% correct) is shown by the horizontal dashed line. Like humans, ferrets with normal hearing have thresholds that are about 10 dB better in the bilateral condition than in the control condition, showing binaural unmasking. Unilateral conductive hearing loss impaired bilateral thresholds without substantially affecting control thresholds. (C) Mean (and S.D.) level of binaural unmasking is shown for normally hearing ferrets and in animals that received a unilateral hearing loss for several months. The reduction in unmasking seen during earplugging takes several months to recover following its removal. Adapted from Moore et al. [87].Fig. 29.10. Auditory experience shapes the maturation of sound localization behavior and the map of auditory space in the SC. (A) Stimulus–response plots showing the combined data of three normally reared ferrets (“Normal adult ferrets”), another three animals just after inserting an earplug into the left ear (“Adult left earplug”), and three ferrets that had been raised and tested with the left ear occluded with a plug that produced 30–40 dB attenuation (“Reared with left earplug”). These plots illustrate the distribution of approach-to-target responses (ordinate) as a function of stimulus location (abscissa). The stimuli were bursts of broadband noise. The size of the dots indicates, for a given speaker angle, the proportion of responses made to different response locations. Correct responses fall on the diagonal (x = y). Occluding one ear disrupts sound localization accuracy, but adaptive changes take place during development that enable the juvenile plugged ferrets to localize sound almost as accurately as the controls. (B) The map of auditory space in the ferret SC, illustrated by plotting the best azimuth of neurons versus their location within the nucleus. Occluding one ear disrupts this spatial tuning, but, as with the behavioral data, near-normal spatial tuning is present in ferrets that were raised with one ear occluded. Based on King et al. [88].
Plasticity of auditory spatial processing in the SC has also been demonstrated by manipulating other sensory cues. As mentioned in an earlier section of this chapter, a characteristic feature of the organization of the SC across different species is the presence of topographically aligned sensory maps for each of the modalities represented there. Bringing together spatial signals that are registered by different sense organs is challenging, but studies in ferrets and barn owls have been at the forefront in showing that vision, which generally provides more precise and reliable spatial information, plays an instructive role in merging multisensory spatial information during development through its influence on the maturation of the auditory space map (reviewed in Ref. 89) (Fig. 29.11). These studies suggest that visual inputs to the SC might provide a template for guiding the development and plasticity of the auditory responses [89–91], so that information from the different senses can be combined and coordinated appropriately.
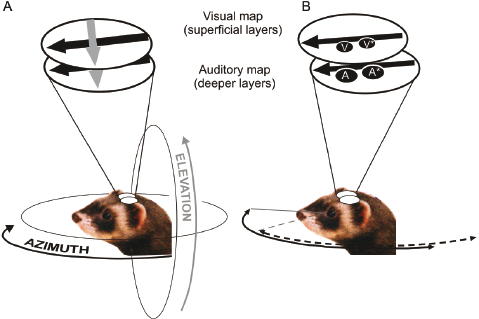
Fig. 29.11. Representation of sensory space in the ferret SC. (A) The superficial layers of the SC are exclusively visual, whereas neurons in the deeper layers respond to auditory (as well as visual and tactile) stimuli. Stimulus azimuth is mapped onto the rostrocaudal axis (black arrow), and elevation onto the mediolateral (gray arrow) axis of the nucleus. The portion of visual and auditory space represented within the SC is indicated by the arrows around the ferret's head, and corresponds approximately to the extent of the visual field of the contralateral eye. Based on King and Hutchings [49]. (B) Visual calibration of the developing auditory space map in the ferret SC. Removal of the medial rectus muscle just before natural eye opening induces a small, outward deviation of the eye. The visual field is therefore shifted laterally relative to the head. This is shown schematically as the receptive field of a superficial-layer neuron in the contralateral SC shifting from position V to V*. Electrophysiological recordings in adult ferrets revealed that the auditory receptive fields of deep SC neurons are also shifted laterally by a corresponding amount (A → A*) following this procedure. As a result of this adaptive change, the correspondence between the superficial-layer visual map and the deep-layer auditory map is preserved. Data source from King et al. [110].
Dynamic Neural Coding and Plasticity in the Adult Brain
A growing number of studies have demonstrated that sensory systems can adjust their sensitivity to compensate for the often considerable changes in input statistics between different sensory scenes. Studies in ferrets have shown that the response properties of neurons in the IC [44] and auditory cortex [92–94] are not fixed, but can change rapidly over time. For example, cortical neurons rescale their gain to compensate for the contrast between a sound and its background: when the contrast is low, the gain of the neurons increases so that they became more sensitive to level changes, but when contrast is high, they become less sensitive [92,93]. The stimulus values to which auditory neurons are tuned can also change in response to the recent history of stimulation [44,95].
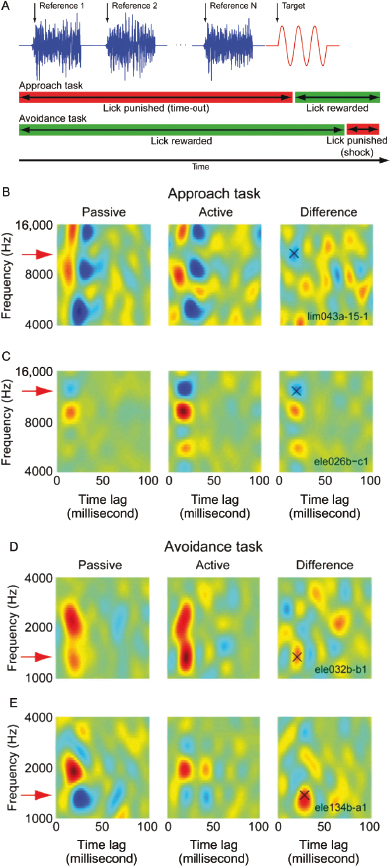
Another remarkable example of the plasticity of the adult brain has been demonstrated by studies in ferrets showing that the spectrotemporal receptive fields (STRFs) of auditory cortical neurons can change rapidly when animals are engaged in a behavioral task [96,97]. In these experiments, ferrets were trained by aversive conditioning to refrain from licking a spout when they detected a target tone or a change in the frequency of the sound in order to avoid a mild electric shock. The selectivity of the neurons was found to change rapidly, sharpening the representations of the stimuli that were relevant to the specific demands of the task. These changes, which were readily observed after just a few trials and sometimes persisted for several hours [97], are thought to reflect the influence of attention on cortical processing. However, the nature of the task is not the only factor that determines how the STRFs change. Its consequences are also important, as experiments in which ferrets were trained to make different behavioral responses in the same sound discrimination task have revealed [98] (Fig. 29.12). The animals either had to suppress their licking behavior to avoid a mild electrical shock or were rewarded for licking a spout when they correctly identified the target sound. Changes in the STRFs were observed in both paradigms, but occurred in opposite directions with responses to the target sound becoming enhanced in the avoidance task, but suppressed when the animals were rewarded. Both changes promote better discriminability, but it seems that the responses of cortical neurons to stimuli associated with aversive outcomes are selectively amplified.
Fig. 29.12. Cortical tuning changes during behavior. (A) Ferrets were trained to detect a pure tone target (red) after a random number of reference noise sounds (blue) using two different behavioral methods. During the approach behavior, the animals were positively rewarded with water for licking a water spout 0.1–1.0 second after target onset (green bar) and punished with a timeout for licking earlier (red bar). During the avoidance task, the ferrets were rewarded by licking a continuously flowing stream of water during the reference sounds and punished with a mild tail shock if they failed to cease licking 0.4 second after target offset. (B) Data from one neuron during the approach task. The spectrotemporal receptive field (STRF) of the neuron was estimated from responses to reference sounds during passive listening (left) and indicates which stimulus frequencies and time lags correlated with increased (red) or suppressed (blue) neural spiking. This neuron was excited broadly by 8000- to 16,000-Hz stimuli, which overlapped the target tone (11,046 Hz). During behavior (center), a notch appeared at the target frequency in the excitatory region, and the difference between the active and passive STRFs (right) shows a 23% decrease in gain at the target frequency (“X”). (C) STRF change for a second neuron during approach, plotted as in B. Here the target frequency (11,300 Hz) overlapped an inhibitory subregion of the passive STRF. During behavior, the inhibition grew stronger, producing a net 7% decrease at the target frequency. (D) STRF change for a neuron during avoidance behavior, with the target positioned on the shoulder of an excitatory region of the passive STRF (1250 Hz). The STRF showed a selective 10% increase at the target frequency during behavior. (E) Data from a second neuron recorded during avoidance, with the target positioned over an inhibitory region (1350 Hz). The inhibition was mostly abolished during behavior, resulting in a 30% response increase at the target frequency. Reprinted, with permission, from David et al. [98].
Plasticity of cortical responses is also associated with the perceptual improvements associated with long-term training. The sensitivity of neurons to the trained stimulus can be altered in various ways. While some studies have focused on changes in the proportion of neurons that respond to behaviorally relevant aspects of the sound (e.g., Refs. 99 and 100), recordings in ferrets have shown that learning on a complex sound discrimination task is associated with increases in the amount of information that cortical neurons transmit about these stimuli in their temporal firing patterns [101]. Moreover, behavioral studies in adult ferrets have demonstrated that the capacity of the auditory system to compensate for abnormal spatial cues is not restricted to a sensitive period of development. As in humans, plugging one ear changes the binaural cue values corresponding to each direction in space and results in a profound deficit in sound localization, which is characterized by an increase in the number and magnitude of the errors made with a notable bias toward the side of nonoccluded ear. If provided with appropriate training, however, adult ferrets wearing a unilateral earplug rapidly recover their ability to localize sound, with the extent and rate of that recovery being determined by the frequency with which they are trained [102] (Fig. 29.13A). This capacity to compensate for a conductive hearing loss in one ear has considerable implications for rehabilitation strategies following the loss or restoration of hearing, and led directly to studies that demonstrated an equivalent capacity for learning-induced plasticity of spatial hearing in humans [103].
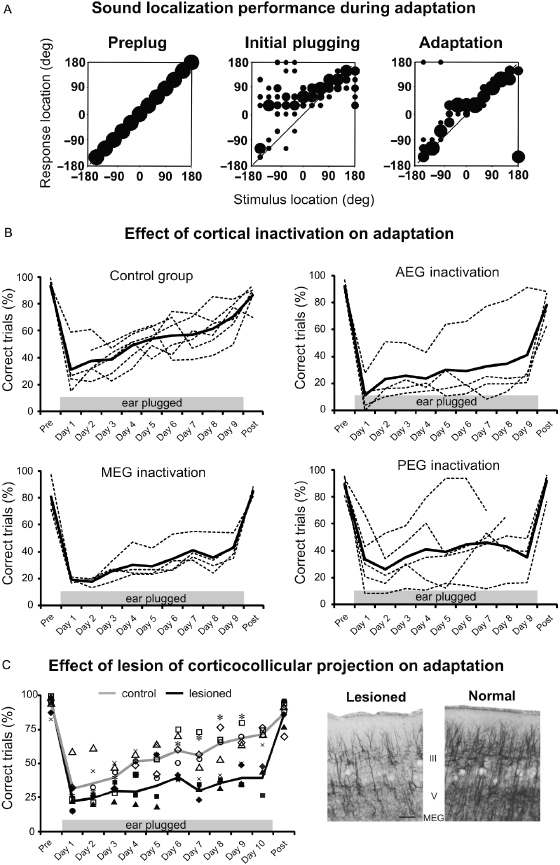
Fig. 29.13. Experience-dependent plasticity of spatial hearing in adult ferrets. (A) Stimulus–response plots showing the distribution of responses (ordinate) made by a ferret as a function of stimulus location in the horizontal plane (abscissa). Prior to occlusion of the left ear, the animal achieved 100% correct scores at all stimulus directions (left panel), but performed poorly, particularly on the side of the earplug, when the left ear was occluded (middle). Further testing with the earplug still in place, however, led to a recovery in localization accuracy (right). Modified from Kacelnik et al. [102]. (B) Effect of cortical inactivation on adaptation. Each panel shows data from a different group of animals that received a subdural implant of a slow release polymer over some part of auditory cortex. The top left panel are drug-free controls, while the other three panels show the effects of inactivating the cortex by placing muscimol-Elvax over the middle (MEG), anterior (AEG), or posterior (PEG) ectosylvian gyrus. The percentage of correct trials measured every day from individual animals and averaged across all speaker locations are shown by the dashed lines, with the mean performance for each group indicated by the solid lines. Data are shown prior to insertion of an earplug in the left ear (Pre), on each of the 9 days that the earplug was in place (days 1–9), and following its removal (Post). Deactivating any of these cortical areas impaired adaptation, whereas the drug-free controls learned normally. Modified from Nodal et al. [78]. (C) Elimination of descending corticocollicular neurons by chromophore-targeted laser photolysis also impaired the ability of ferrets to adapt to an earplug. Different symbols correspond to different animals and the lines show the mean performance of control (gray) and lesioned (black) groups. The photomicrographs on the right illustrate the loss of layer V pyramidal cells in the lesioned cortex compared with intact cortex. Modified from Bajo et al. [39].
In keeping with the physiological evidence in other species that perceptual learning is associated with changes in the response properties of neurons in early sensory cortex, the ability of adult ferrets to adapt to the altered spatial cues produced by plugging one ear is lost if the auditory cortex is silenced [76,78] (Fig. 29.13B) or if cholinergic inputs to the cortex from the basal forebrain are removed [104]. The finding that adaptation depends on cholinergic neuromodulation implies a possible role for attention in this process and is consistent with other work, suggesting that acetylcholine plays a critical role in sensory processing under challenging conditions. The experience-dependent recalibration of sound localization in adult ferrets has also been used to demonstrate the behavioral importance of descending projections in the auditory system. Thus, unilateral ablation of the projection from A1 to the IC by chromophore-targeted laser photolysis impaired the spatial plasticity that normally occurs after plugging one ear (Fig. 29.13C), without affecting the ability of the animals to localize sound under normal hearing conditions [39]. Behavioral adaptation by adult ferrets to a unilateral earplug also depends on the integrity of the olivocochlear bundle, the descending projection from the SOC, where sensitivity to binaural localization cues is first derived, to the cochlea [38].
Cochlear Implant Model
With the advent of cochlear implants, it is possible to partially restore hearing in the deaf by directly stimulating the inner ear. Although traditionally fitted in one ear only, bilateral implantation is becoming more common, particularly in children, in an attempt to improve sound localization and hearing in noisy environments, which both rely heavily on having two functioning ears. However, results from patients have been quite variable and it is likely that age at onset of deafness, duration of implant use, and targeted auditory training can influence the performance of cochlear implant users. There is, therefore, a pressing need for animal models that can be used to investigate both the behavioral and physiological outcomes of cochlear implantation and the importance of experience in restoring auditory function. Although a range of species have been used to study the effects of intracochlear electrical stimulation, the size of the speech processors that are currently available commercially restricts their use in behavioral studies to larger mammals, such as primates or carnivores.
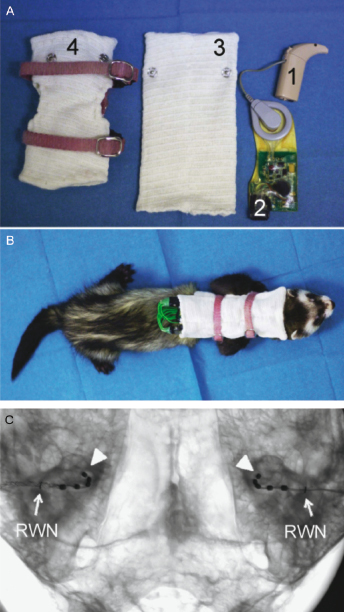
The development of a ferret model of cochlear implantation [105] (Fig. 29.14) has been driven by the substantial amount of physiological and behavioral data available on numerous aspects of hearing in this species, and, in particular, by the evidence described in the previous sections that ferrets are particularly suitable animals for investigating the development and plasticity of auditory function. Protocols have been developed for deafening ferrets with aminoglycoside administration, followed by implantation and chronic stimulation of intracochlear electrical arrays in one or both ears. By connecting the electrodes to clinical processors via modified stimulator–receivers worn within a custom-made jacket (Fig. 29.14), behaviorally relevant ILDs and ITDs can be provided during chronic stimulation, which are used by the animals to orient toward acoustic stimuli [105]. These experiments provide a unique opportunity for investigating the role of experience and training in establishing and maintaining auditory spatial abilities, and should also contribute to the improvement of neuroprosthetic therapies in humans.
Fig. 29.14. Ferret cochlear implantation. (A) Nucleus ESPrit 3G speech processor (1) attached to a modified Nucleus Cochlear implant CI24RE emulator (2). Pockets were incorporated within the neckline of a detachable “backpack” (3) that was attached to a jacket made from a ferret harness and elasticated tubular bandage (4). (B) Ferrets carried their cochlear implants within the jacket, which held the microphone of the left and right speech processor immediately posterior to the ipsilateral pinna and enabled animals to carry on with their normal activities during chronic stimulation. (C) High resolution microfocus radiograph in anterior–posterior view of a ferret skull showing bilateral intracochlear arrays inserted to an even depth into the basal turn of both cochleas relative to the round window niche (RWN) marked with a fine wire. The apical electrode in the electrode array is marked with an arrowhead. Reproduced, with permission, from Hartley et al. [105].
Concluding Remarks
In this chapter, we have attempted to provide an overview of our current knowledge about hearing in ferrets and how this species is contributing to advances in our understanding of many aspects of auditory function. Thirty years ago, the viability of the ferret as a model of auditory processing was questioned (D.R. Moore, personal communication), whereas today they are one of the most commonly used species for studying the higher auditory system. It is likely that the popularity of the ferret as a model for auditory research will continue to increase thanks to the technical advances that allow neural activity to be recorded in freely moving, behaving animals. The prospect of combining the wide repertoire of behavioral tasks that can be performed by ferrets with chronic recording of neural activity will undoubtedly further our understanding of complex functions such as speech recognition or scene analysis and is likely to be key to revealing the nature of the changes that give rise to highly prevalent conditions such as tinnitus. Moreover, the advent of new ways of controlling neural activity by light through the use of optogenetics, which are now being used in a range of species, including ferrets, is rapidly expanding the methods available for studying neural circuits, and may pave way toward new treatment possibilities for a range of neurological conditions.
Acknowledgments
The authors' research is supported by a Wellcome Trust Principal Research Fellowship (WT076508AIA) to AJK.
References
1. Kelly JB, Kavanagh GL, Dalton JC (1986) Hearing in the ferret (Mustela putorius): thresholds for pure tone detection. Hear Res 24: 269–275.
2. Kavanagh GL, Kelly JB (1987) Contribution of auditory cortex to sound localization by the ferret (Mustela putorius). J Neurophysiol 57: 1746–1766.
3. King AJ, Parsons CH (1999) Improved auditory spatial acuity in visually deprived ferrets. Eur J Neurosci 11: 3945–3956.
4. Nodal FR, Bajo VM, Parsons CH, Schnupp JW, King AJ (2008) Sound localization behavior in ferrets: comparison of acoustic orientation and approach-to-target responses. Neuroscience 154: 397–408.
5. Parsons CH, Lanyon RG, Schnupp JW, King AJ (1999) Effects of altering spectral cues in infancy on horizontal and vertical sound localization by adult ferrets. J Neurophysiol 82: 2294–2309.
6. Bizley JK, Nodal FR, Parsons CH, King AJ (2007) Role of auditory cortex in sound localization in the midsagittal plane. J Neurophysiol 98: 1763–1774.
7. Heffner RS, Heffner HE (1992) Visual factors in sound localization in mammals. J Comp Neurol 317: 219–232.
8. Kelly JB, Kavanagh GL (1994) Sound localization after unilateral lesions of inferior colliculus in the ferret (Mustela putorius). J Neurophysiol 71: 1078–1087.
9. Schnupp J, Nelken I, King A (2011) Auditory neuroscience: making sense of sound. Cambridge, MA: MIT Press.
10. Nodal FR, Keating P, King AJ (2010) Chronic detachable headphones for acoustic stimulation in freely moving animals. J Neurosci Methods 189: 44–50.
11. Keating P, Nodal FR, Gananandan K, Schulz AL, King AJ (2013) Behavioral sensitivity to broadband binaural localization cues in the ferret. J Assoc Res Otolaryngol 14: 561–572.
12. Keating P, Nodal FR, King AJ (2013) Behavioural sensitivity to binaural spatial cues in ferrets: evidence for plasticity in the duplex theory of sound localization. Eur J Neurosci 39: 197–206.
13. Bizley JK, Walker KM, King AJ, Schnupp JW (2013) Spectral timbre perception in ferrets: discrimination of artificial vowels under different listening conditions. J Acoust Soc Am 133: 365–376.
14. Walker KM, Bizley JK, King AJ, Schnupp JW (2011) Cortical encoding of pitch: recent results and open questions. Hear Res 271: 74–87.
15. Kalluri S, Depireux DA, Shamma SA (2008) Perception and cortical neural coding of harmonic fusion in ferrets. J Acoust Soc Am 123: 2701–2716.
16. Yin P, Fritz JB, Shamma SA (2010) Do ferrets perceive relative pitch? J Acoust Soc Am 127: 1673–1680.
17. Walker KM, Schnupp JW, Hart-Schnupp SM, King AJ, Bizley JK (2009) Pitch discrimination by ferrets for simple and complex sounds. J Acoust Soc Am 126: 1321–1335.
18. Kelly JB, Rooney BJ, Phillips DP (1996) Effects of bilateral auditory cortical lesions on gap-detection thresholds in the ferret (Mustela putorius). Behav Neurosci 110: 542–550.
19. Hine JE, Martin RL, Moore DR (1994) Free-field binaural unmasking in ferrets. Behav Neurosci 108: 196–205.
20. Ma L, Micheyl C, Yin P, Oxenham AJ, Shamma SA (2010) Behavioral measures of auditory streaming in ferrets (Mustela putorius). J Comp Psychol 124: 317–330.
21. Tolnai S, Litovsky RY, King AJ (2014) The precedence effect and its build-up and breakdown in ferrets and humans. J Acoust Soc Am (in press).
22. Alves-Pinto A, Sollini J, Sumner CJ (2012) Signal detection in animal psychoacoustics: analysis and simulation of sensory and decision-related influences. Neuroscience 220: 215–227.
23. Rarey KE (1985) The ferret as a model for inner ear research. Lab Anim Sci 35: 238–241.
24. Carlile S (1990) The auditory periphery of the ferret. II: the spectral transformations of the external ear and their implications for sound localization. J Acoust Soc Am 88: 2196–2204.
25. Carlile S, King AJ (1994) Monaural and binaural spectrum level cues in the ferret: acoustics and the neural representation of auditory space. J Neurophysiol 71: 785–801.
26. Keating P, Dahmen JC, King AJ (2013) Context-specific reweighting of auditory spatial cues following altered experience during development. Curr Biol 23: 1291–1299.
27. Webster DB, Popper AN, Fay RR, eds. (1992) The mammalian auditory pathway: neuroanatomy. New York: Springer-Verlag.
28. Moore DR (1988) Auditory brainstem of the ferret: sources of projections to the inferior colliculus. J Comp Neurol 269: 342–354.
29. Fuentes-Santamaría V, Alvarado JC, Henkel CK, Brunso-Bechtold JK (2007) Cochlear ablation in adult ferrets results in changes in insulin-like growth factor-1 and synaptophysin immunostaining in the cochlear nucleus. Neuroscience 148: 1033–1047.
30. Henkel CK, Gabriele ML (1999) Organization of the disynaptic pathway from the anteroventral cochlear nucleus to the lateral superior olivary nucleus in the ferret. Anat Embryol (Berl) 199: 149–160.
31. Henkel CK, Brunso-Bechtold JK (1993) Laterality of superior olive projections to the inferior colliculus in adult and developing ferret. J Comp Neurol 331: 458–468.
32. King AJ, Jiang ZD, Moore DR (1998) Auditory brainstem projections to the ferret superior colliculus: anatomical contribution to the neural coding of sound azimuth. J Comp Neurol 390: 342–365.
33. Nodal FR, Doubell TP, Jiang ZD, Thompson ID, King AJ (2005) Development of the projection from the nucleus of the brachium of the inferior colliculus to the superior colliculus in the ferret. J Comp Neurol 485: 202–217.
34. Angelucci A, Clascá F, Sur M (1998) Brainstem inputs to the ferret medial geniculate nucleus and the effect of early deafferentation on novel retinal projections to the auditory thalamus. J Comp Neurol 400: 417–439.
35. Bajo VM, King AJ (2013) Cortical modulation of auditory processing in the midbrain. Front Neural Circuits 6: 114.
36. Bajo VM, Nodal FR, Bizley JK, Moore DR, King AJ (2007) The ferret auditory cortex: descending projections to the inferior colliculus. Cereb Cortex 17: 475–491.
37. Bajo VM, Nodal FR, Bizley JK, King AJ (2010) The non-lemniscal auditory cortex in ferrets: convergence of corticotectal inputs in the superior colliculus. Front Neuroanat 4: 18.
38. Irving S, Moore DR, Liberman MC, Sumner CJ (2011) Olivocochlear efferent control in sound localization and experience-dependent learning. J Neurosci 31: 2493–2501.
39. Bajo VM, Nodal FR, Moore DR, King AJ (2010) The descending corticocollicular pathway mediates learning-induced auditory plasticity. Nat Neurosci 13: 253–260.
40. Popper AN, Fay RR, eds. (1992) The mammalian auditory pathway: neurophysiology. New York: Springer-Verlag.
41. Moore DR, Semple MN, Addison PD (1983) Some acoustic properties of neurones in the ferret inferior colliculus. Brain Res 269: 69–82.
42. Kelly JB, Judge PW, Phillips DP (1986) Representation of the cochlea in primary auditory cortex of the ferret (Mustela putorius). Hear Res 24: 111–115.
43. Sumner CJ, Palmer AR (2012) Auditory nerve fibre responses in the ferret. Eur J Neurosci 36: 2428–2439.
44. Dahmen JC, Keating P, Nodal FR, Schulz AL, King AJ (2010) Adaptation to stimulus statistics in the perception and neural representation of auditory space. Neuron 66: 937–948.
45. Shechter B, Marvit P, Depireux DA (2010) Lagged cells in the inferior colliculus of the awake ferret. Eur J Neurosci 31: 42–48.
46. Garcia-Lazaro JA, Ahmed B, Schnupp JW (2011) Emergence of tuning to natural stimulus statistics along the central auditory pathway. PLoS ONE 6: e22584.
47. Rabinowitz NC, Willmore BDB, King AJ, Schnupp JWH (2013) Constructing noise-invariant representations of sound in the auditory pathway. PLoS Biol 11: e1001710 .
48. Schnupp JW, King AJ (1997) Coding for auditory space in the nucleus of the brachium of the inferior colliculus in the ferret. J Neurophysiol 78: 2717–2731.
49. King AJ, Hutchings ME (1987) Spatial response properties of acoustically responsive neurons in the superior colliculus of the ferret: a map of auditory space. J Neurophysiol 57: 596–624.
50. King AJ (2004) The superior colliculus. Curr Biol 14: R335–R338.
51. Stein BE, Stanford TR, Rowland BA (2009) The neural basis of multisensory integration in the midbrain: its organization and maturation. Hear Res 258: 4–15.
52. Campbell RA, Doubell TP, Nodal FR, Schnupp JW, King AJ (2006) Interaural timing cues do not contribute to the map of space in the ferret superior colliculus: a virtual acoustic space study. J Neurophysiol 95: 242–254.
53. King AJ, Moore DR, Hutchings ME (1994) Topographic representation of auditory space in the superior colliculus of adult ferrets after monaural deafening in infancy. J Neurophysiol 71: 182–194.
54. Kowalski N, Versnel H, Shamma SA (1995) Comparison of responses in the anterior and primary auditory fields of the ferret cortex. J Neurophysiol 73: 1513–1523.
55. Nelken I, Bizley JK, Nodal FR, Ahmed B, Schnupp JW, King AJ (2004) Large-scale organization of ferret auditory cortex revealed using continuous acquisition of intrinsic optical signals. J Neurophysiol 92: 2574–2588.
56. Bizley JK, Nodal FR, Nelken I, King AJ (2005) Functional organization of ferret auditory cortex. Cereb Cortex 15: 1637–1653.
57. Bizley JK, Nodal FR, Bajo VM, Nelken I, King AJ (2007) Physiological and anatomical evidence for multisensory interactions in auditory cortex. Cereb Cortex 17: 2172–2189.
58. Keniston LP, Allman BL, Meredith MA, Clemo HR (2009) Somatosensory and multisensory properties of the medial bank of the ferret rostral suprasylvian sulcus. Exp Brain Res 196: 239–251.
59. Ramsay AM, Meredith MA (2004) Multiple sensory afferents to ferret pseudosylvian sulcal cortex. Neuroreport 15: 461–465.
60. Manger PR, Engler G, Moll CK, Engel AK (2005) The anterior ectosylvian visual area of the ferret: a homologue for an enigmatic visual cortical area of the cat? Eur J Neurosci 22: 706–714.
61. Kayser C, Petkov CI, Logothetis NK (2009) Multisensory interactions in primate auditory cortex: fMRI and electrophysiology. Hear Res 258: 80–88.
62. Bizley JK, King AJ (2008) Visual-auditory spatial processing in auditory cortical neurons. Brain Res 1242: 24–36.
63. Ghazanfar AA, Chandrasekaran C, Logothetis NK (2008) Interactions between the superior temporal sulcus and auditory cortex mediate dynamic face/voice integration in rhesus monkeys. J Neurosci 28: 4457–4469.
64. Kayser C, Logothetis NK, Panzeri S (2010) Visual enhancement of the information representation in auditory cortex. Curr Biol 20: 19–24.
65. Schnupp JW, Mrsic-Flogel TD, King AJ (2001) Linear processing of spatial cues in primary auditory cortex. Nature 414: 200–204.
66. Bizley JK, Walker KM, Silverman BW, King AJ, Schnupp JW (2009) Interdependent encoding of pitch, timbre, and spatial location in auditory cortex. J Neurosci 29: 2064–2075.
67. Walker KM, Bizley JK, King AJ, Schnupp JW (2011) Multiplexed and robust representations of sound features in auditory cortex. J Neurosci 31: 14565–14576.
68. Mesgarani N, David SV, Fritz JB, Shamma SA (2008) Phoneme representation and classification in primary auditory cortex. J Acoust Soc Am 123: 899–909.
69. Mesgarani N, David SV, Fritz JB, Shamma SA (2009) Influence of context and behavior on stimulus reconstruction from neural activity in primary auditory cortex. J Neurophysiol 102: 3329–3339.
70. Rauschecker JP, Tian B, Pons T, Mishkin M (1997) Serial and parallel processing in rhesus monkey auditory cortex. J Comp Neurol 382: 89–103.
71. Lomber SG, Malhotra S (2008) Double dissociation of “what” and “where” processing in auditory cortex. Nat Neurosci 11: 609–616.
72. Bizley JK, Walker KM, King AJ, Schnupp JW (2010) Neural ensemble codes for stimulus periodicity in auditory cortex. J Neurosci 30: 5078–5091.
73. Bendor D, Wang X (2005) The neuronal representation of pitch in primate auditory cortex. Nature 436: 1161–1165.
74. Bizley JK, Walker KM, Nodal FR, King AJ, Schnupp JW (2013) Auditory cortex represents both pitch judgments and the corresponding acoustic cues. Curr Biol 23: 620–625.
75. Kavanagh GL, Kelly JB (1992) Midline and lateral field sound localization in the ferret (Mustela putorius): contribution of the superior olivary complex. J Neurophysiol 67: 1643–1658.
76. Nodal FR, Kacelnik O, Bajo VM, Bizley JK, Moore DR, King AJ (2010) Lesions of the auditory cortex impair azimuthal sound localization and its recalibration in ferrets. J Neurophysiol 103: 1209–1225.
77. Smith AL, Parsons CH, Lanyon RG, Bizley JK, Akerman CJ, Baker GE, Dempster AC, Thompson ID, King AJ (2004) An investigation of the role of auditory cortex in sound localization using muscimol-releasing Elvax. Eur J Neurosci 19: 3059–3072.
78. Nodal FR, Bajo VM, King AJ (2012) Plasticity of spatial hearing: behavioural effects of cortical inactivation. J Physiol 590: 3965–3986.
79. Moore DR, Hine JE (1992) Rapid development of the auditory brainstem response threshold in individual ferrets. Brain Res Dev Brain Res 66: 229–235.
80. Henkel CK, Keiger CJ, Franklin SR, Brunso-Bechtold JK (2007) Development of banded afferent compartments in the inferior colliculus before onset of hearing in ferrets. Neuroscience146: 225–235.
81. Campbell RA, King AJ, Nodal FR, Schnupp JW, Carlile S, Doubell TP (2008) Virtual adult ears reveal the roles of acoustical factors and experience in auditory space map development. J Neurosci 28: 11557–11570.
82. Mrsic-Flogel TD, Schnupp JWH, King AJ (2003) Acoustic factors govern developmental sharpening of spatial tuning in the auditory cortex. Nat Neurosci 6: 981–988.
83. Schnupp JW, Booth J, King AJ (2003) Modeling individual differences in ferret external ear transfer functions. J Acoust Soc Am 113: 2021–2030.
84. Moore DR (1994) Auditory brainstem of the ferret: long survival following cochlear removal progressively changes projections from the cochlear nucleus to the inferior colliculus. J Comp Neurol 339: 301–310.
85. McAlpine D, Martin RL, Mossop JE, Moore DR (1997) Response properties of neurons in the inferior colliculus of the monaurally deafened ferret to acoustic stimulation of the intact ear. J Neurophysiol 78: 767–779.
86. Moore DR, Hutchings ME, King AJ, Kowalchuk NE (1989) Auditory brain stem of the ferret: some effects of rearing with a unilateral ear plug on the cochlea, cochlear nucleus, and projections to the inferior colliculus. J Neurosci 9: 1213–1222.
87. Moore DR, Hine JE, Jiang ZD, Matsuda H, Parsons CH, King AJ (1999) Conductive hearing loss produces a reversible binaural hearing impairment. J Neurosci 19: 8704–8711.
88. King AJ, Parsons CH, Moore DR (2000) Plasticity in the neural coding of auditory space in the mammalian brain. Proc Natl Acad Sci U S A 97: 11821–11828.
89. King AJ (1999) Sensory experience and the formation of a computational map of auditory space in the brain. Bioessays 21: 900–911.
90. Knudsen EI, Brainard MS (1991) Visual instruction of the neural map of auditory space in the developing optic tectum. Science 253: 85–87.
91. King AJ, Schnupp JW, Thompson ID (1998) Signals from the superficial layers of the superior colliculus enable the development of the auditory space map in the deeper layers. J Neurosci 18: 9394–9408.
92. Rabinowitz NC, Willmore BD, Schnupp JW, King AJ (2012) Spectrotemporal contrast kernels for neurons in primary auditory cortex. J Neurosci 32: 11271–11284.
93. Rabinowitz NC, Willmore BD, Schnupp JW, King AJ (2011) Contrast gain control in auditory cortex. Neuron 70: 1178–1191.
94. Depireux DA, Dobbins HD, Marvit P, Shechter B (2012) Dynamics of phase-independent spectro-temporal tuning in primary auditory cortex of the awake ferret. Neuroscience 214: 28–35.
95. Dahmen JC, Hartley DE, King AJ (2008) Stimulus-timing-dependent plasticity of cortical frequency representation. J Neurosci 28: 13629–13639.
96. Fritz JB, Elhilali M, Shamma SA (2005) Differential dynamic plasticity of A1 receptive fields during multiple spectral tasks. J Neurosci 25: 7623–7635.
97. Fritz J, Shamma S, Elhilali M, Klein D (2003) Rapid task-related plasticity of spectrotemporal receptive fields in primary auditory cortex. Nat Neurosci 6: 1216–1223.
98. David SV, Fritz JB, Shamma SA (2012) Task reward structure shapes rapid receptive field plasticity in auditory cortex. Proc Natl Acad Sci U S A 109: 2144–2149.
99. Recanzone GH, Schreiner CE, Merzenich MM (1993) Plasticity in the frequency representation of primary auditory cortex following discrimination training in adult owl monkeys. J Neurosci 13: 87–103.
101. Schnupp JW, Hall TM, Kokelaar RF, Ahmed B (2006) Plasticity of temporal pattern codes for vocalization stimuli in primary auditory cortex. J Neurosci 26: 4785–4795.
102. Kacelnik O, Nodal FR, Parsons CH, King AJ (2006) Training-induced plasticity of auditory localization in adult mammals. PLoS Biol 4: e71.
103. Kumpik DP, Kacelnik O, King AJ (2010) Adaptive reweighting of auditory localization cues in response to chronic unilateral earplugging in humans. J Neurosci 30: 4883–4894.
104. Leach ND, Nodal FR, Cordery PM, King AJ, Bajo VM (2013) Cortical cholinergic input is required for normal auditory perception and experience-dependent plasticity in adult ferrets. J Neurosci 33: 6659–6671.
105. Hartley DE, Vongpaisal T, Xu J, Shepherd RK, King AJ, Isaiah A (2010) Bilateral cochlear implantation in the ferret: a novel animal model for behavioral studies. J Neurosci Methods 190: 214–228.
106. Phillips DP, Judge PW, Kelly JB (1988) Primary auditory cortex in the ferret (Mustela putorius): neural response properties and topographic organization. Brain Res 443: 281–294.
107. Moore BCJ, Glasberg BR (1983) Formulae describing frequency selectivity as a function of frequency and level, and their use in calculating excitation pattern. Hear Res 28: 209–225.
108. Palmer AR, Russell IJ (1986) Phase-locking in the cochlear nerve of the guinea-pig and its relation to the receptor potential of inner hair cells. Hear Res 24: 1–15.
109. Johnson DH (1980) The relationship between spike rate and synchrony in responses of auditory-nerve fibers to single tones. J Acoust Soc Am 68: 1115–1122.
110. King AJ, Hutchings ME, Moore DR, Blakemore C (1988) Developmental plasticity in the visual and auditory representations in the mammalian superior colliculus. Nature 332: 73–76.