3
FROM BLOOD GROUPS TO GENES
There are few things more distinctive about a person than their hair. It is one of the very first features we ask for in any description of a new baby, a stranger or a wanted criminal. Dark or blonde, wavy or straight, thick or balding: all these different possibilities add immediately to the picture we build up in our minds of someone we have never met. We certainly know how to manipulate the way our own hair appears. Salons are full as we pay to have our hair cut and shaped. Pharmacy shelves are lined with products to lighten, darken, straighten and curl. We are all working to make the best of the hair we were born with; but it is our genes which deal out the basic raw material. The difference between a natural redhead and a blonde lies in a difference in their DNA. Within the genes for keratin and the many others involved in the process of growing hair are small differences in the DNA sequence. These are responsible for giving the hair different characteristics of colour and texture. Most of these genes have yet to be identified, but they are certain to be inherited from both parents, although not necessarily in a straightforward way – which is why it is a fairly frequent occurrence that a new baby does not have the hair colour of either of its parents.
Hair type is a highly visible distinguishing feature by which we tell individuals apart, but by far the greatest inherited differences between us are invisible and remain hidden unless something brings them to our attention. The first of these inherited differences to be revealed were the blood groups. You cannot tell just by looking at someone which blood group he or she belongs to. You can’t even tell by simply looking at a drop of their blood. All blood looks pretty much the same. It is only when you begin to mix blood from two people that the differences begin to make themselves apparent; and, since no-one had any reason to mix one person’s blood with another until blood transfusions were invented, our blood groups stayed hidden.
The first blood transfusions were recorded in Italy in 1628, but so many people died from the severe reactions that the practice was banned there, as well as in France and England. Though there were some experimental transfusions using lamb’s blood, notably by the English physician Richard Lower in the 1660s, the results were no better and the idea was given up for a couple of centuries. Transfusions with human blood started up again in the middle of the nineteenth century, to combat the frequently fatal haemorrhages that occurred after childbirth, and by 1875 there had been 347 recorded transfusions. But many patients were still suffering the sometimes fatal consequences of a bad reaction to the transfused blood.
By that time, scientists were beginning to discover the differences in blood type that were causing the problem. The nature of the reaction of one blood type with another was discovered by the French physiologist Léonard Lalois when, in 1875, he mixed the blood of animals of different species. He noticed that the blood cells clumped together and frequently burst open. But it was 1900 before the biologist Karl Landsteiner worked out what was happening and discovered the first human blood group system, which divides people into Groups A, B, AB and O. When a donor’s ABO blood group matches that of the patient receiving the transfusion, there is no bad reaction; but if there is a mismatch, the cells form clumps and break open, causing a severe reaction. There is some historical evidence that the Incas of South America had practised transfusions successfully. Since we now know that most native South Americans have the same blood group (Group O), the Inca transfusions would have been much less dangerous than attempts in Europe, because there was an excellent chance that both donor and patient would belong to Group O and thus be perfectly matched.
Unlike the complicated genetics which governs the inheritance of hair, which is still not fully understood, the rules for inheriting the ABO blood groups turned out to be very simple indeed. Precisely because the genetics were so straightforward and could be followed easily from parents to offspring, blood groups were widely used in cases of contested paternity until recently, when they were eclipsed by the much greater precision of genetic fingerprints. Their significance for our story in this book is that it was the blood groups which first launched genetics on to the world stage of human evolution. For this debut we need to go back to the First World War and to a scientific paper delivered to the Salonika Medical Society on 5 June 1918. It was translated and published the following year in the leading British medical periodical The Lancet under the title ‘Serological differences between the blood of different races: the results of research on the Macedonian Front’. To give you a flavour of the sort of thing The Lancet published in those days, the article was sandwiched between a discourse by the eminent surgeon Sir John Bland-Sutton on the third eyelid of reptiles and a War Office announcement that those nurses who had been mentioned in dispatches for their work in Egypt and France would soon be getting a certificate from the King showing his appreciation.
The authors of the blood group paper were a husband and wife team, Ludwik and Hanka Herschfeld, who worked at the central blood group testing laboratory of the Royal Serbian Army, which was part of the Allied force fighting against the Germans. The First World War had a great influence on bringing blood transfusion practice towards its modern standards. Before the war it had been customary for physicians with a patient who needed a transfusion to test the blood groups of friends and relatives until a match was found, then bleed the donor and immediately give the blood to the patient. The high demand for transfusions on the battlefields of Europe meant that ways had to be found to store donated blood in blood banks ready for immediate use. All soldiers had their blood group tested and recorded so that, should they need an urgent transfusion to treat a serious battlefield wound, compatible blood of the correct type could be immediately drawn from the blood bank.
Ludwik Herschfeld had already demonstrated, some years earlier, that blood groups A and B followed the basic genetic rules laid out by Gregor Mendel. He was not sure what to make of blood group O and set it aside, though it was later shown to obey the same rules. Herschfeld saw the war as an opportunity to discover more about blood groups, and in particular how they compared in different parts of the world. The Allies drew soldiers from many different countries, and the Herschfelds set out to collate the blood group results from as many different nationalities as possible. It was a lot of work, but easier in wartime than later, when the research would, as they put it, ‘have necessitated long years of travel’. For the obvious military reason that they were on the other side, they did not have the German data to hand, and the figures published in The Lancet were ‘quoted from memory’.
When the Herschfelds came to review the results of their work, they found very big differences in the frequencies of blood groups A and B in soldiers who came from different ‘races’ as they called them. Among the Europeans, the proportions were around 15 per cent blood group B and 40 per cent blood group A. The proportion of men with blood group B was higher in troops drawn from Africa and Russia, reaching a peak of 50 per cent in regiments of the Indian Army fighting with the British. As the proportion of blood group B increased, there was a corresponding decrease in the frequency of blood group A.
In drawing their conclusions, the Herschfelds did not flinch from interpreting the significance of their results on a grand scale. They decided that humans were made up of two different ‘biochemical races’, each with its own origin: Race A, with blood group A, and Race B, with blood group B. Because Indians had the highest frequency of blood group B, they concluded that ‘We should look to India as the cradle of one part of humanity.’ As to how blood groups, and populations, spread, they go on: ‘Both to Indo-China in the East and to the West a broad stream of Indians passed out, ever-lessening in its flow, which finally penetrated to Western Europe.’ They were unsure about the origin of Race A and thought it might have come from somewhere around north or central Europe. We know now that their conclusions are complete nonsense; but they do illustrate that geneticists, then as now, are never shy of grandiose speculation.
The basic principle behind the evolutionary inferences drawn from the Herschfelds’ blood group results was that ‘races’ or ‘populations’ that have similar proportions of the different blood groups are more likely to share a common history than those where the proportions are very different. This sounds like common sense, and it looks like a reasonable explanation for the similarities found in the different European armies. But there were also some surprises. For example, the blood group frequencies of soldiers from Madagascar and Russia were almost identical. Did this mean the Herschfelds had uncovered genetic evidence for a hitherto unrecorded Russian invasion of Madagascar, or even the reverse, an overwhelming Malagasy colonization of Russia? Or take the Senegalese from West Africa, who were almost as close in their blood group frequencies to the Russians as the English were to the Greeks, which seems a bit unusual to say the least. What was happening was that because they were working with just one genetic system – the only one available to them – their analysis produced what appear to be some very reasonable comparisons between populations and others that look distinctly odd.
In the years after the First World War, it fell to the American physician William Boyd to compile the abundant blood group data coming from transfusion centres throughout the world. As he did so, he saw inconsistencies of the Russia/Madagascar kind revealed by the original Herschfeld results time and again, so frequently, in fact, that he actively discouraged anthropologists from taking any notice of blood groups. Boyd quotes a letter from one frustrated correspondent: ‘I tried to see what blood groups would tell me about ancient man and found the results very disappointing.’ Even so, the unsuccessful attempts to explain human origins using blood groups had had their compensations for the liberal-minded Boyd. He wrote: ‘In certain parts of the world an individual will be considered inferior if he has, for instance, a dark skin but in no part of the world does possession of a blood group A gene exclude him from the best society.’
After the Second World War, William Boyd’s baton as compiler of blood group data from around the world passed to the Englishman Arthur Mourant. A native of Jersey in the Channel Islands, Mourant originally took a degree in geology but was unable to translate that training into a career. His very strict Methodist upbringing had caused him considerable emotional unhappiness, which he determined to resolve by becoming a psychoanalyst. To do this he decided first to study medicine and enrolled, at the relatively late age of thirty-four, in St Bartholomew’s Medical School in London. This was in 1939, just before the outbreak of the Second World War. To avoid the German bombing raids on the capital, his medical school was moved from London to Cambridge, and it was here that he met R. A. Fisher, the most influential geneticist of his day. Fisher had been working out the genetics of the new blood groups which were being discovered, and he had become fascinated by the particularly convoluted inheritance of one of them – the Rhesus blood group. This new group had been discovered by Karl Landsteiner and his colleague Alexander Wiener in 1940 after they mixed human blood with the blood of rabbits that had themselves been injected with cells of the Rhesus monkey (hence the name). Fisher had come up with a complicated theory to account for the way in which the different sub-types within the group were passed down from parents to their children, and this was being violently attacked by Wiener who had offered a much simpler explanation. Imagine Fisher’s delight when the new arrival, Arthur Mourant, discovered a large family of twelve siblings which provided the practical proof of his theory. Fisher found him a job at once, and the meticulous Mourant spent the rest of his working life compiling and interpreting the most detailed blood group frequency distribution maps ever produced. He never did become a psychoanalyst.
As well as being instrumental in getting Arthur Mourant a job, the Rhesus blood groups were also about to play a central role in what people were thinking about the origins of modern Europeans and in identifying the continent’s most influential genetic population – the fiercely independent Basques of north-west Spain and south-west France. The Basques are unified by their common language, Euskara, which is unique in Europe in that it has no linguistic connection with any other living language. That it survives at all in the face of its modern rivals, Castilian Spanish and French, is remarkable enough. But two thousand years ago, it was only the disruption of imperial Roman administration in that part of the empire that saved Euskara from being completely swamped by Latin, which was the fate of the now extinct Iberian language in eastern Spain and south-east France. The Basques provided us with an invaluable clue to the genetic history of the whole of Europe, as we shall see later in the book, but their elevation to special genetic status only began when Arthur Mourant started to look closely at the Rhesus blood groups.
Most people have heard about the Rhesus blood groups in connection with ‘blue baby syndrome’ or ‘haemolytic disease of the new-born’ to give it its full medical title. This serious and often fatal condition affects the second or subsequent pregnancy of mothers who are ‘Rhesus negative’ – that is, who do not possess the Rhesus antigen on the surface of their red blood cells. What happens is this. When a Rhesus negative mother bears the child of a Rhesus positive father (whose red blood cells do carry the Rhesus antigen), there is a high probability that the foetus will be Rhesus positive. This is not a problem for the first child; but, when it is being born, a few of its red blood cells may get into the mother’s circulation. The mother’s immune system recognizes these cells, with their Rhesus antigen, as foreign, and begins to make antibodies against them. That isn’t a problem for her until she becomes pregnant with her next child. If this foetus is also Rhesus positive then it will be attacked by her anti-Rhesus antibodies as they pass across the placenta. New-born babies affected in this way, who appear blue through lack of oxygen in their blood, could sometimes be rescued by a blood transfusion, but this was a risky procedure. Fortunately, ‘blue baby syndrome’ is no longer a severe clinical problem today. All Rhesus negative mothers are now given an injection of antibodies against Rhesus positive blood cells, so that if any do manage to get into her circulation during the birth of her first child they will be mopped up before her immune system has a chance to find them and start to make antibodies.
The significance of all this to the thinking about European prehistory is that Mourant realized that having two Rhesus blood groups in a single population did not make any evolutionary sense. Even the simplest calculations showed that losing so many babies was not a stable arrangement. There was no problem if everybody had the same Rhesus type. It didn’t matter whether this was Rhesus positive or Rhesus negative, just so long as it was all one or the other. It was only when there were people with different Rhesus types breeding together that these very serious problems arose. In the past, before blood transfusions and before the antibody treatment for Rhesus negative mothers, there must have been a lot of babies dying from haemolytic disease. This is a very heavy evolutionary burden, and the expected result of this unbalanced situation would be that one or other of the Rhesus blood groups would eventually disappear. And this is exactly what has happened – everywhere except in Europe. While the rest of the world is predominantly Rhesus positive, Europe stands out as having a very nearly equal frequency of both types. To Mourant, this was a signal that the population of Europe was a mixture that had not yet had time to settle down and eliminate one or other of the Rhesus types. His explanation was that modern Europe might be a relatively recent hybrid population of Rhesus positive arrivals from the Near East, probably the people who brought farming into Europe beginning about eight thousand years ago, and the descendants of an earlier Rhesus negative hunter-gathering people. But who were the Rhesus negatives?
Mourant came across the work of the French anthropologist H. V. Vallois, who described features of the skeletons of contemporary Basques as having more in common with fossil humans from about twenty thousand years ago than with modern people from other parts of Europe. Though this kind of comparison has since fallen into disrepute, it certainly catalysed Mourant’s thinking. It was already known that Basques had by far the lowest frequency of blood group B of all the population groups in Europe. Could they be the ancient reservoir of Rhesus negative as well? In 1947 Mourant arranged to meet with two Basques who were in London attempting to form a provisional government and were keen to support any attempts to prove their genetic uniqueness. Like most Basques, they were supporters of the French Resistance and totally opposed to the fascist Franco regime in Spain. Both men provided blood samples and both were Rhesus negative. Through these contacts, Mourant typed a panel of French and Spanish Basques who turned out, as he had hoped, to have a very high frequency of Rhesus negatives, in fact the highest in the world. Mourant concluded from this that the Basques were descended from the original inhabitants of Europe, whereas all other Europeans were a mixture of originals and more recent arrivals, which he thought were the first farmers from the Near East.
From that moment, the Basques assumed the status of the population against which all ideas about European genetic prehistory were to be – and to a large extent still are – judged. The fact that they alone of all the west Europeans spoke a language which was unique in Europe, and did not belong to the Indo-European family which embraces all other languages of western Europe, only enhanced their special position.
The next leap forward came from the mathematical amalgamation of the vast amount of data that had accumulated from decades of research on individual systems like the different blood groups. This was accomplished by the man who has dominated the field for the past thirty years, Luigi Luca Cavalli-Sforza. We will meet him again later. Cavalli-Sforza, working with the Cambridge statistician Anthony Edwards, achieved this amalgamation using the earliest punched-card computing machines. By averaging across several genetic systems at once they managed to eliminate most of the bizarre and counter-intuitive conclusions that had discredited the anthropological applications of blood groups when they were worked on one at a time. The weakness of using just a single system was that two populations, like the Russians and the Malagasy, could end up with the same gene frequency just by chance rather than because of a common ancestry. This was far less likely to happen if several genes were compared, because the impact of a misleading result from one of them would be diluted out by the effect of the others. There were to be no more Russian invasions of Madagascar. None the less, the underlying principle remained the same. In an evolutionary sense, populations with similar gene frequencies were more likely to be closely related to each other than populations whose gene frequencies were very different.
Anthony Edwards explained his thinking in an ingenious article in New Scientist in 1965. He imagines a tribe that carries with it a pole along which are arrayed 100 discs which are either black or white. Every year, one disc, chosen at random, is changed to the other colour. When the tribe splits into two groups, each group takes with it a copy of the pole with the discs in their current order. The following year they each make one of the random changes to the discs. The next year they make another, the next year another and so on, continuing the custom of one random change every year. Since the changes they make are completely random, the order of the discs on the two poles becomes more and more dissimilar as each year passes. It follows that if you were to look at the poles carried by the two tribes you could estimate how long ago, in a relative sense, they separated from each other by the differences in the order of the black and white discs. Providing an absolute date was very difficult from the gene frequency data alone, but the comparative separation between the two tribes, known as the genetic distance, was a useful measure of their common ancestry. The bigger the genetic distance between them, the longer they had spent apart.
This was a clever image of the process of genetic change, called genetic drift, brought about by the random survival and extinction of genes as they pass from one generation to the next. This process leads to bigger and bigger differences in the frequencies of genes as time passes. Just like the order of discs in Edwards’ analogy, gene frequencies can be used to backtrack and work out how long ago two groups of people were once together as a single population. These groups could be villages, tribes or whole populations, and there is no limit to the number of groups that can be analysed in this way. If you do it for the whole world, the outcome is a diagram like Figure 1 overleaf.
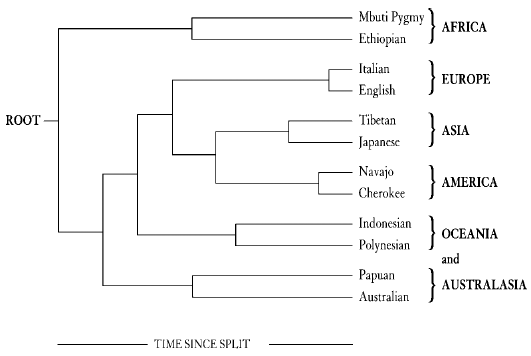
Figure 1
Along the right-hand side we have several ‘populations’ (I have picked two examples from each continent) and along the bottom is the genetic distance/time axis. This is what is called a population tree where the lines trace, from left to right, the estimated order in which ‘populations’ evolved and split from one another, as reconstructed from the assimilated frequencies of many different genes. At first glance, many of the groupings look quite sensible. The two European populations, the English and the Italians, are close together on two short ‘branches’ of the tree. The two native American tribes are connected together with their closest relatives in Asia, as we would expect if the first Americans crossed the Bering land bridge from Siberia to Alaska. The two populations from Africa are on a different branch from the rest of the world, which correctly emphasizes that continent’s great antiquity as the cradle of human evolution. This is a much more sensible-looking tree than can be drawn from the First World War blood group data which, as well as allying Russia and Madagascar, entirely missed the importance of Africa. The reason for this, as noted earlier, is that the odd quirks that arose by chance with a single system, like the ABO blood groups, get ironed out by amalgamating the results from several different genes.
Edwards acknowledged that ‘The resultant evolutionary trees will certainly not provide the last word on human evolution,’ and offered the diagrams as a way of providing the genetic information in an understandable form. Unfortunately, the population trees first drawn with this admirable and modest intention were over-interpreted and became a source of contention. Among the several reasons for this is just the way they appear. They do look as if they are real evolutionary trees and have often been portrayed as exactly that. They could only be true evolutionary trees if human evolution really were a succession of population fissions along the lines of the splits that Edwards explains in his metaphor of the tribes with their poles and discs. Then and only then would the nodes, the points on the tree from which two lines diverge, represent a real entity. These would be the populations that existed before the splits, the proto-populations. But is that what really happened in human evolution? For instance, in the European part of the tree, was there ever such a thing as the proto-Anglo-Italian population which divided, never to meet again, and became the modern inhabitants of England and Italy? That might have been the case if the English and Italians became two different species as soon as they split and could never interbreed again. But they can, and they do, and they always have done. As we will discover later in the book, humans just did not evolve like this.
Perhaps the most serious objection to these trees is that their construction demands that the things at the end of the trees, the populations, be objectively defined. This process in itself segregates people into groups in ways that can tend to perpetuate racial classifications. It gives some sort of overall genetic number to something that does not really exist. There are certainly people who live in Japan and Tibet, but there is no genetic meaning to the population of Tibet or Japan, taken as a whole. As this book will show, objectively defined races simply do not exist. Even Arthur Mourant realized that fact nearly fifty years ago, when he wrote: ‘Rather does a study of blood groups show a heterogeneity in the proudest nation and support the view that the races of the present day are but temporary integrations in the constant process of…mixing that marks the history of every living species.’ The temptation to classify the human species into categories which have no objective basis is an inevitable but regrettable consequence of the gene frequency system when it is taken too far. For several years the study of human genetics got firmly bogged down in the intellectually pointless (and morally dangerous) morass of constructing ever more detailed classifications of human population groups.
Fortunately, there was a way out of this impasse. The break-out came with the publication of a scientific paper in Nature in January 1987 by the veteran US evolutionary biochemist, the late Allan Wilson, and two of his students, Rebecca Cann and Mark Stoneking, entitled ‘Mitochondrial DNA and human evolution’. The centrepiece of this article was a diagram which bears a superficial resemblance to the trees I have just been criticizing. I have reproduced a small section of it here in Figure 2, with only sixteen individuals instead of the 134 in the original paper.
It is indeed an evolutionary tree; but this time the diagram means something. On the right of the tree the symbols at the tips of the branches represent not populations but the sixteen individuals that I have selected to illustrate the point, sixteen people from four different parts of the world: Africans, Asians, Europeans and Papuans from New Guinea. The first improvement over the other trees is that, unlike populations, there is no argument about whether people exist or not. They clearly do. The other improvement is that the nodes on the tree are also real people and not some hypothetical concept like a ‘proto-population’. They represent the last common ancestors of the two people who branch off from that point. The lines that connect the sixteen people on the diagram are drawn to reflect genetic differences between them in one very special gene called mitochondrial DNA whose unusual and useful properties I will introduce shortly. For reasons I shall explain in the next chapter, if two people have very similar mitochondrial DNA then they are more closely related, with respect to this gene, than two people with very different mitochondrial DNA. They have a common ancestor who lived more recently in the past, and so are joined by shorter branches on the diagram. People with very different mitochondrial DNA share a more remote common ancestor and are linked by longer branches.
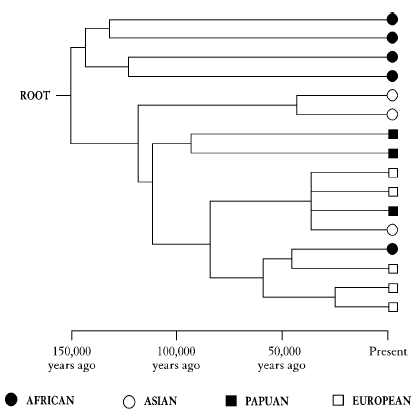
Figure 2
To see how this works we can use again the metaphor of the tribe with its pole holding black and white discs. But this time the pole is the mitochondrial DNA and the tribe that split in two is a person who has two children. Both children inherit the same mitochondrial DNA, the genetic equivalent of the same pattern of discs on the pole. When they have their own children they pass on the mitochondrial DNA to them, and so it goes on down the generations. Very occasionally, random changes, called mutations, occur in the mitochondrial DNA which alter it a little bit at a time. These occur quite by chance when the DNA is being copied as cells divide. As time passes, more random changes are added to the DNA, which are then retained and passed on to future generations. Very slowly, the mitochondrial DNA of the descendants of that first individual, their common ancestor, becomes more and more different as more random mutations are introduced one at a time.
The lines on the tree in Figure 2 are reconstructions of the relationships among these sixteen people, worked out from the differences in their mitochondrial DNA, the exact nature of which we will examine shortly. But look for the moment at the tree itself. The deep trunk at the top has four Africans at the tips, while the other deep trunk contains individuals from the rest of the world and one more African. Within this ‘rest of the world’ trunk, close branches sometimes connect people from the same part of the world, like the Asians and Papuans at the top or the Europeans at the bottom. But they also sometimes connect individuals from different places, like the branch near the middle that links a Papuan with an Asian and two Europeans. What’s going on? The deep split between the exclusively African ‘trunk’ and the rest of the world is another confirmation of the antiquity of Africa which the population trees also pick up. The confusion in the ‘rest of the world’ trunk is confirmation of exactly what Arthur Mourant had in mind. It is ‘the mixing that marks the history of every living species’. Small wonder, then, that this diagram threw a very large spanner in the works of the population tree aficionados. It shows that genetically related individuals are cropping up all over the place, in all the wrong populations. You just cannot sustain the fundamental idea of a population being a separate biological and genetic unit if individuals within one population have their closest relatives within another.
Moreover, as we shall see in greater detail later on, by using the mutation process just described we can estimate the rate at which mitochondrial DNA changes with time. This means we can work out the timescales involved. When we do that, all the branches and the trunks converge to a single point, the ‘root’ of the tree, at about 150,000 years ago. This had to mean that the whole of the human species was much younger and more closely related than many people thought.
The impact of ‘Mitochondrial DNA and human evolution’ was dramatic. It came down very firmly on one side of the argument about a fundamental question of human evolution. For many years there had been an intense and polarized debate on the origins of modern humans, based on different interpretations of fossil skeletons, mainly the skull. Both sides agreed that modern Homo sapiens, the species to which we all belong, originated in Africa. Both sides also agreed that an earlier type of human, called Homo erectus, was an evolutionary intermediate between ourselves and much older and more ape-like fossils. Homo erectus first appeared in Africa about two million years ago and by one million years ago, or perhaps even earlier, it had spread out to the warmer parts of the Old World. Homo erectus fossils have been found from Europe in the west to China and Indonesia in the east.
All that was – and is – agreed by both sides of the argument. What divides them is whether or not there was a much more recent spread of modern humans from Africa. The ‘Out of Africa’ school think there was, about 100,000 years ago, and that these new humans, our own Homo sapiens, completely replaced Homo erectus throughout its range. The opposing school of thought, the multi-regionalists, see clues in the fossils that suggest to them that Homo sapiens evolved directly from their local Homo erectus populations. This would mean that modern Chinese, for example, are directly descended from Chinese Homo erectus, and modern Europeans are similarly evolved from European Homo erectus, rather than being descendants of Homo sapiens who migrated from Africa. In the multi-regional scheme a modern European and a modern Chinese would have last shared a common ancestor at least one million years ago, while in the ‘Out of Africa’ scenario they would be linked very much more recently.
What the mitochondrial gene tree did was to introduce an objective time-depth measurement into the equation for the first time. It showed quite clearly that the common mitochondrial ancestor of all modern humans lived only about 150,000 years ago. This fitted in very well with the ‘Out of Africa’ theory and was enthusiastically welcomed by its supporters. But it came as a severe shock to the multi-regionalists. If all modern humans were related back to a common ancestor as recently as 150,000 years ago, they could not possibly have evolved in different parts of the world from local populations of Homo erectus that had been in place for well over a million years. Though the multi-regionalists, being thoroughly modern humans themselves, have refused to accept defeat, the mitochondrial gene tree dealt a wounding blow to their theory from which it has not yet recovered.
For us, it was great news. Mitochondrial DNA was catapulted by this controversy into its position as the prime molecular interpreter of the human past. A surge of research effort was bound to follow in laboratories all over the world. And that meant there would be lots of data with which we could compare our own results. If we were going to put the results from the old bones into a modern context, then we could not do better than use mitochondrial DNA.