
5.1 táblázat. Annak valószínűsége, hogy az emberi DNS-hez hasonló örökítőanyag-szakaszokat találunk más élőlényekben - attól függően, hogy fehérjekódoló vagy a gének között elhelyezkedő DNS-szakaszt veszünk alapul.
//////////////
Amikor az 1980-as évek elején a Yale Egyetemen kutattam, a DNS-kód néhány száz betűjének megfejtése is rendkívül fáradságos vállalkozásnak számított. A módszer igen érzékeny volt mindenféle zavaró hatásra; számos előkészítő lépést igényelt, illetve veszélyes és drága reagensek használatát. Például radioaktív vegyszerekét és ultravékony gélekét, amelyeket kézzel kellett csurgatni, így szinte mindig buborékok és más hibák keletkeztek bennük. A részletek nem is fontosak, a lényeg az, hogy a folyamat egy örökkévalóságig tartott. A kutató rengeteget próbálkozott, és rendszerint kudarcot vallott. Mindezt azért, hogy az emberi DNS csupán néhány száz betűjét meghatározza.
E nehézségek ellenére első humán genetikai tudományos közleményem a DNS-szekvenálásról szólt. Egyetlen fehérje termelődését vizsgáltam, amely a méhben fejlődő emberi magzat vörösvértestjeiben található, és fokozatosan eltűnik a születés után, amikor a babák már saját tüdejükkel lélegeznek. Ezt a fehérjét magzati hemoglobinnak nevezzük. A hemoglobin olyan protein, amely lehetővé teszi, hogy a vörösvértestek elszállítsák az oxigént a tüdőből a test más részeibe. Az emberek és néhány emberszabású majom születésük előtt a hemoglobin egy különleges változatát használják, amely képes kivonni az oxigént az anya véréből, hogy ezzel táplálja a fejlődő magzatot. Az első életévben ez a magzati hemoglobin fokozatosan megszűnik működni, a szervezet pedig a molekula „felnőtt” változatát kezdi termelni helyette. Egy jamaicai családban, amelyet alkalmam nyílt megvizsgálni, a magzati hemoglobin felnőttkorban is nagy mennyiségben jelen volt. A magzati hemoglobin „örökletes fennmaradása” rengeteg tudóst foglalkoztatott. Ha ugyanis rájöttünk volna, hogyan indítható be újra a magzati hemoglobin termelése, azzal nagyban csökkenthettük volna a sarlósejtes vérszegénység okozta szenvedéseket. Ha a magzati hemoglobinnak csupán 20 százalékát vissza tudtuk volna állítani a sarlósejtes vérszegénységben szenvedők vörösvértesteiben, azzal gyakorlatilag meg tudtuk volna szüntetni a betegség fájdalmas tüneteit, és meg tudtuk volna előzni a visszafordíthatatlan szervkárosodást.
Soha nem felejtem el azt a napot, amikor szekvenálásra irányuló erőfeszítéseim eredményre vezettek. Kiderült, hogy a jamaicai család DNS-ének egy különleges helyén G áll a szokásos C helyett, az egyik olyan gén előtt, amely beindítja a magzati hemoglobin termelését. Mint kiderült, ennek az egy betűnek a megváltozása okozta, hogy a magzati program a felnőttekben is tovább futott. A felismerés lenyűgözött, ugyanakkor teljesen kimerült voltam - ugyanis tizennyolc hónapig tartott, hogy felfedezzünk egyetlen megváltozott betűt a DNS-kódban.
Így aztán igencsak megdöbbentem, amikor három évvel később arról értesültem, hogy néhány látnoki képességekkel megáldott tudós értekezni kezdett róla, hogyan lehetne meghatározni a teljes emberi genom DNS-szekvenciáját, amelynek hosszát hárommilliárd bázispárra becsülték. Na, ez az a cél - gondoltam amelyet az én életemben biztosan nem érünk el.
Akkoriban viszonylag keveset tudtunk arról, mit tartalmazhat a teljes genom. Valójában még senki nem látta az emberi gének kémiai szerkezetét mikroszkóp alatt - ehhez ugyanis túl kicsik.
Addig csak néhány száz gént jellemeztek, és teljesen eltérő becslések születtek arról, hogy összesen hány gént is tartalmazhat a teljes genom. Még a gén definíciója sem volt egyértelmű - és ez a mai napig nem változott. Az egyszerű definíció szerint a gén olyan DNS-szakaszt tartalmaz, amely egy bizonyos fehérjét kódol. Csakhogy ez a meghatározás megdőlt, amikor felfedezték, hogy a géneken belül a fehérjekódoló szakaszokat nem kódoló szakaszok szakítják meg, amelyeket intronoknak nevezünk. Attól függően, hogy a kódoló régiók később, az RNS-másolat készítésekor hogyan illesztődnek össze, egy gén időnként számos különböző, bár rokon fehérjét is kódolhat. Továbbá olyan hosszú DNS-szakaszokat is találtak a gének között, amelyeknek mintha semmi szerepük nem lett volna - néhányan el is nevezték őket „hulladék DNS-nek” (junk DNA). Persze hatalmas elbizakodottságra vall, hogy a DNS valamelyik szakaszát bárki is „hulladéknak” merte nevezni, tekintve, hogy akkor még alig tudtunk valamit a DNS valódi működéséről.
Minden bizonytalanság ellenére sem volt kérdéses, hogy milyen rendkívüli eredmény lenne, ha ismernénk a teljes emberi genom kódsorrendjét (szekvenciáját). Hiszen ebben a hatalmas használati utasításban elrejtve ott találhatjuk valahol az ember biológiai alkatrészeinek felsorolását, illetve a szükséges tudást egy sor kevéssé értett és emiatt rosszul kezelhető betegség gyógyításához. Orvosként számomra elképesztően izgalmas volt annak a lehetősége, hogy feltárhatom a létező legtöbb tudást tartalmazó orvosi tankönyv lapjait. Bár még a tudományos ranglétra alján álltam, és nem is voltam biztos benne, hogy egy efféle vakmerő terv megvalósítható-e a gyakorlatban, mégis bekapcsolódtam a vitába. Amellett érveltem, hogy szükség lenne egy szervezett programra az emberi génkészlet meghatározásához - ez hamarosan humán genom projektként vált ismertté.
A vágyam, hogy teljesen feltárjuk az emberi genomot, a következő években csak erősödött. Egy szárnyait bontogató kutatólaboratóriumot vezettem, amelyben eltökélt és nagy munkabírású egyetemisták és posztdoktori kutatók dolgoztak. Úgy döntöttem, olyan betegségek genetikai alapjait fogjuk vizsgálni, amelyek egyelőre ellenálltak a hátterük feltárására irányuló próbálkozásoknak. Ezek közül a legfontosabb a cisztás fibrózis volt, az észak-európaiak körében leggyakoribb, potenciálisan halálos genetikai rendellenesség. A betegséget általában olyan újszülötteknél, illetve kisgyermekeknél diagnosztizálják, akik nem növekednek megfelelően, emellett újabb és újabb légzőszervi fertőzéseket kapnak el. A jó megfigyelőképességgel rendelkező anyák észrevették, hogy ezeknek a gyerekeknek sós az ízük, amikor megpuszilják őket. Az orvosok pedig ebből az információból kiindulva felismerték: a betegség legkönnyebben észlelhető jele, hogy az érintett gyerekek verejtékében túl magas a klór koncentrációja. Azt is tudtuk, hogy a cisztás fibrózisban szenvedő betegek tüdejében és hasnyálmirigyében sűrű, ragadós váladék gyűlik fel - de nem tudtuk, mi lehet annak a génnek a pontos funkciója, amely hibásan működik.
Először akkor találkoztam a cisztás fibrózissal, amikor orvos gyakornokként dolgoztam az 1970-es évek végén. Az ötvenes években ezek a beteg gyerekek ritkán élték meg a tizedik születésnapjukat. De folyamatosan fejlődött a tünetek kezelése és szinten tartása - például a hasnyálmirigyenzimek kiváltása, a táplálkozás és a fizikoterápia erősítése, illetve a tüdőfertőzések egyre hatékonyabb antibiotikumokkal történő kezelése által. Így fokozatosan sikerült meghosszabbítani a cisztás fibrózisban szenvedők élettartamát. A hetvenes években sokan közülük már elég hosszú ideig éltek ahhoz, hogy egyetemre járjanak, megházasodjanak, és munkát vállaljanak. A hosszú távú gyógyítás azonban továbbra is reménytelennek tűnt. Mivel nem ismertük a genetikai defektus mibenlétet, a kutatóorvosok úgy érezték, csak a sötétben tapogatóznak. Mindössze annyit tudtunk, hogy az emberi DNS-kód hárommilliárd betűje közül legalább egy, egy fontos helyen lévő betű hibás.
Egy ilyen apró eltérés megtalálása abban az időben szinte lehetetlen feladatnak tűnt. Viszont tudtunk még valamit a cisztás fibrózisról: azt, hogy recesszív módon öröklődik. Hogy megértsük ennek jelentőségét, tudnunk kell, hogy minden génünkből két másolattal rendelkezünk. Egyiket édesanyánktól, a másikat apánktól örököljük. (Kivételt képeznek ez alól azok a gének, amelyek az X, illetve az Y nemi kromoszómán foglalnak helyet, mivel ezekből a férfiakban csak egy másolat található.) A cisztás fibrózishoz hasonló, recesszív betegségek esetén a gyermek csak akkor betegszik meg, ha mindkét öröklött génje - ezeket alléleknek nevezzük - hibás. Ahhoz, hogy ez bekövetkezzen, mindkét szülőnek rendelkeznie kell egy hibás génmásolattal. Viszont azok, akik egy hibás és egy egészséges változatot hordoznak a génből, teljesen egészségesek, ezért nekik általában fogalmunk sincs az állapotukról. Nagyjából minden harminc, észak-európai felmenőkkel rendelkező emberből egy hordozza a hibás cisztásfibrózis-gént, de többségük családjában sosem fordult elő a betegség.
A cisztás fibrózis ily módon érdekes lehetőséget biztosított a DNS-ben való nyomozásra. Anélkül, hogy bármit is tudtak volna a betegségért felelős génről, a kutatók vissza tudták követni a DNS több száz, véletlenszerűen kiválasztott szakaszának öröklődését olyan családokban, amelyekben megjelent a cisztás fibrózis, és több testvér született. Olyan DNS-részleteket kerestek, amelyek összefüggésben állhattak azzal, hogy melyik testvér betegszik meg cisztás fibrózisban, és melyik nem. Ezeknek a részleteknek - más néven fragmentumoknak - szükségszerűen a cisztás fibrózisért felelős gén közelében kellett elhelyezkedniük a genomban. Nem tudtuk elolvasni mind a hárommilliárd betűpárt, de véletlenszerűen meg tudtunk világítani néhány milliót itt, néhány milliót amott, és próbáltunk összefüggést találni köztük, illetve a betegség között. Mindezt sok száz alkalommal el kellett végeznünk, de mivel a genom információtartalma véges, biztosak lehettünk benne, hogy ha nem adjuk fel, előbb-utóbb rátalálunk a cisztás fibrózisért felelős gén helyére.
Végül 1985-ben sikerült. A kutatók és az érintett családok ugyanúgy megdöbbentek az eredményen, és ugyanúgy örültek neki. Kimutattuk, hogy a cisztás fibrózist okozó génnek a hetedik kromoszóma kétmillió bázispár hosszúságú szakaszán kell lennie. A feladat nehezebbik része viszont csak most következett. Akkoriban a következő analógiával próbáltam rávilágítani, hogy miért volt rendkívül nehezen megoldható ez a probléma. A keresés ahhoz hasonlított, mintha egyetlen kiégett villanykörtét próbáltunk volna megtalálni egy ház alagsorában, valahol az Egyesült Államokban. A családkutatás megfelelő kezdetnek bizonyult - abban az értelemben, hogy segítségével sikerült meghatároznunk, melyik államban, majd melyik megyében van a kérdéses villanykörte. De ezáltal is csak úgy szemlélhettük a genomot, mintha egy repülőről néztünk volna le, és így kerestük volna a kiégett villanykörtét. Ez a stratégia egy adott ponton túl már nem vitt közelebb a megoldáshoz. Házról házra, izzóról izzóra kellett átvizsgálnunk a környéket.
Ráadásul nem volt térképünk a kérdéses területről. A hetes kromoszómának ezt a területét - a genom nagy részéhez hasonlóan - 1985 előtt még soha nem vizsgálták. Hogy folytassam a fenti hasonlatot, nem rendelkeztünk térképpel az átkutatandó városokról és falvakról, nem ismertük az épületek alaprajzát, és nem voltak jegyzékeink a villanykörtékről. Az elvégzendő munka embertelennek tűnt.
Csapatommal kifejlesztettünk egy módszert, amelyet „kromoszómaugrásnak” neveztük el. Ennek segítségével úgy haladhattunk a kétmillió bázispár hosszúságú célterület átvizsgálásával, mintha csak hétmérföldes csizmában lépkedtünk volna - ahelyett, hogy a hagyományos módon araszoltunk volna. A sebességet tovább tudtuk növelni azáltal, hogy a házról házra folytatott kutatást egyszerre több helyszínen kezdtük meg. De a feladat még így is szinte teljesíthetetlennek tűnt. A tudományos közösség számos tagja azon a véleményen volt, hogy ez a megközelítés kivitelezhetetlen, egy emberi betegség genetikai hátterének meghatározására sosem lesz használható. Aztán 1987-ben, miközben anyagi forrásaink fogyatkozóban voltak, frusztrációnk pedig nőttön-nőtt, kutatócsoportom összefogott Lap-Chee Tsui laborjával, aki tehetséges PhD-kutatóként a torontói gyermekkórházban dolgozott. Egyesített laborjaink újult erővel folytatták a kutatómunkát. A keresés egy detektívtörténethez hasonlított: tudtuk, hogy a rejtély végül biztosan megoldódik majd az utolsó oldalon, de azt nem is sejtettük, hogy mikor jutunk el oda. Számos nyomra bukkantunk, de legalább ugyanennyiszer vakvágányra is futottunk. Időközben már harmadjára vagy negyedjére fogott el minket a lehetséges megoldás miatt érzett izgalom - aztán másnap minden reményünk odalett, amikor szembesültünk az új adatokkal. Egy idő után már óvakodtunk a túlzott optimizmustól, bármi is történt. Egyre nehezebb volt megmagyarázni a kollégáknak, hogy miért nem leltünk rá még mindig a génre - vagy éppenséggel azt, hogy miért nem adtuk már fel az egészet. Egy alkalommal - hogy újabb metaforát használjak a probléma nehézségének érzékeltetésére - még egy helyi michigani farmra is ellátogattam, hogy ott lefényképezzenek egy hatalmas szénakazalon ülve, egy varrótűvel a kezemben.
1989 májusának egyik esős éjjelén végül megszületett a válasz. A Yale Egyetem kollégiumában, ahol Lap-Chee és jómagam éppen értekezletet tartottunk, a faxgép egyszer csak kiadta az aznapi laboratóriumi munka eredményeit. Amelyek egyértelműen azt mutatták, hogy a betegek nagy részénél egy addig nem ismert gén fehérjekódoló részéből hiányzik mindössze három betű - CTT, hogy pontosak legyünk -, és ez okozza a cisztás fibrózist. Nem sokkal később nekünk és másoknak is sikerült kimutatnunk, hogy ez a mutáció és ugyanennek a génnek (most már CFRT-nek nevezzük) néhány más, kevésbé gyakori „elírása” okozzák a betegséget gyakorlatilag minden érintett személynél.
Így hát bebizonyítottuk, hogy igenis meg lehet találni egy kiégett villanykörtét - vagyis azonosítani tudtuk a betegségért felelős gént azáltal, hogy folyamatosan szűkítettük lehetséges elhelyezkedését a kromoszómán. Nagy pillanat volt ez, és jó okunk volt az ünneplésre. Az idáig vezető út hosszú és nehéz volt, de okkal remélhettük, hogy a gyógymódhoz vezető kutatást immár semmi nem állíthatja meg.
A cisztás fibrózis kutatóinak, a családoknak és a klinikusoknak egy későbbi, több ezer fős találkozójára még egy dalt is írtam, megörökítendő a gén felfedezését. A zene mindig segítségemre volt abban, hogy kifejezzem és átéljem azokat az érzéseket, amelyeket puszta szavakkal nem tudtam megragadni. Habár gitártudásom nagy jóindulattal is csak szerénynek nevezhető, hatalmas örömmel töltött el az a pillanat, amikor több ezer ember egyszerre énekelte a dalt. Ez az élmény inkább szellemi volt számomra, mint tudományos. Nem tudtam visszatartani a könnyeimet, amikor ez a sok remek ember felállt a székéről, és együtt énekelték a refrént.
Merj álmodni, merj álmodni,
Minden testvéred szabadon tud lélegezni.
Nem félünk, szívünk immár szabadon dobog,
Hiszen a tisztás fibrózis története már történelem.
A következő lépések azonban nehezebbnek bizonyultak, mint vártuk, és a cisztás fibrózis sajnos továbbra sem a múlt betegsége. A gén felfedezése ettől függetlenül örömteli érzés volt, és a cisztás fibrózis kutatását olyan pályára állította, amelytől mind azt várjuk, hogy egyszer majd elvezet a végső győzelemhez. Ha összeadjuk mindazt a munkát, amelyet a világ több mint két tucat kutató-csoportja végzett a cisztás fibrózis génjének megtalálása érdekében, kiderül, hogy a munka több mint tíz évbe és ötvenmillió dollárba került. Egyetlen betegség egyetlen génjének felkutatásáról van szó. Ráadásul a cisztás fibrózist azok között a betegségek között tartották számon, amelyek génjét a legkönnyebb megtalálni - mivel viszonylag gyakori betegség, amelynek öröklődése pontosan követi a mendeli törvényeket. Hogyan lehetne ennek fényében kiterjeszteni a kutatómunkát azoknak a sokkal ritkább betegségeknek a százaira, amelyeknek legsürgősebb lett volna feltárni a genetikai hátterét? Még ennél is nehezebbnek tűnt, hogy ezt a stratégiát olyan betegségekre is alkalmazzuk, mint például a diabétesz, a skizofrénia, a szív- és érrendszeri megbetegedések vagy a rák gyakori formái. Ezekről ugyan tudjuk, hogy az örökletes faktorok rendkívül fontosak a kialakulásukban, de jelenlegi tudásunk szerint számos gén egyszerre fejti ki a hatását a betegség kockázatának növelésében. Ráadásul nincs egyetlen olyan gén sem, amelynek hatása domináns lenne. Ezekben az esetekben tucatnyi vagy még több villanykörtét kellett volna megtalálnunk, amelyek még csak nem is égtek ki feltétlenül - mindössze némileg halványabban világítottak, mint kellett volna.
Nyilvánvalóvá vált: ahhoz, hogy bármi reményünk legyen a sikerre ezekben a sokkal bonyolultabb esetekben, részletes információval kell rendelkeznünk az emberi genom minden zegéről és zugáról. Olyan nagy felbontású térképre volt szükségünk az egész országról, amelyen minden egyes ház külön fel van tüntetve.
Az 1980-as években élénk vita folyt arról, van-e értelme egy efféle vállalkozásnak.1 A legtöbb kutató egyetértett abban, hogy az ily módon megszerezhető információ végül is hasznos lehet, a projekt nyilvánvaló grandiózussága miatt sokan mégis úgy gondolták, hogy a célok gyakorlatilag elérhetetlenek. Ekkorra már az is kiderült, hogy a genomnak csak kis hányada kódol fehérjét - a maradék (a hulladék DNS) szekvenálásának értelme legalábbis vitatható volt. Egy közismert tudós azt írta: „A genom szekvenálása nagyjából annyira lenne hasznos, mintha lefordítanánk Shakespeare összes művét ékírásra. Csak kevésbé lenne megvalósítható, és nehezebben lehetne értelmezni.” Másvalaki azt írta: „Ennek az egésznek semmi értelme... A genetikusoknak az értelmetlenség tengerén kellene átgázolniuk ahhoz, hogy végül csakis az információ néhány apró szigetén érezhessenek szárazföldet a talpuk alatt.”
A kétségek többsége megalapozott volt, hiszen egy efféle vállalkozás költségei csillagászatinak ígérkeztek, és fennállt a veszélye annak, hogy ez a pénzigény elszivattyúzná az igencsak korlátozott anyagi forrásokat más orvosi kutatásoktól. A lehetséges veszélyek leghatékonyabb ellenszerének az tűnt, ha sikerül kiterjesztenünk a vállalkozás céljait, ezáltal újabb forrásokat bevonva a finanszírozásba. Ezt tökéletesen megoldotta az Egyesült Államok genomprojektjének új igazgatója: nem más, mint Jim Watson, a DNS kettős spiráljának egyik felfedezője. Watson, aki abban az időben a biológia abszolút rocksztárjának számított, meggyőzte a Kongresszust, hogy támogasson egy efféle vállalkozást.
Jim Watson rendkívül ügyesen irányította az amerikai humán genom projektet az első két évben. Genomkutató központokat hozott létre, és nemzedékük legjobb és legzseniálisabb kutatóit győzte meg arról, hogy a projekten dolgozzanak. Sokan szkeptikusak maradtak azzal kapcsolatban, hogy a projekt képes lesz-e eredményeket felmutatni a kitűzött tizenöt éves határidő lejárta előtt - hiszen a célok eléréséhez szükséges technológia jó részét még fel sem találták. 1992-ben aztán válságba került a projekt. Watson váratlanul otthagyta a vállalkozást, miután nyilvános vitába bonyolódott a Nemzeti Egészségügyi Intézet (National Institutes of Health) (Az orvosbiológiai kutatások amerikai csúcsszerve) igazgatójával arról, hogy szabad-e a genom egyes részeit szabadalmaztatni. Watson ezt határozottan ellenezte.
Lázas fejvadászat kezdődött országszerte, hogy megtaláljuk a projekt következő igazgatóját. Senki nem lepődött meg nálam jobban, amikor kiderült, hogy a kiválasztási folyamat során én váltam a legesélyesebb jelöltté. Mivel akkoriban kifejezetten boldog voltam a Michigani Egyetem genomkutató központjának igazgatójaként, és eszembe sem jutott, hogy egyszer még szövetségi közalkalmazott válhat belőlem, kezdetben nem érdekelt a lehetőség. Aztán lassanként megváltozott a hozzáállásom. Végül is csak egy humán genom projekt létezett. Mindezt csak egyszer viszik majd véghez az emberiség történetében. Ha sikerrel járunk, annak példátlan következményei lesznek az orvoslás szempontjából. Istenhívő emberként arra gondoltam: vajon ez egyike azoknak a pillanatoknak, amikor arra hív az Úr, hogy vállaljak nagyobb szerepet egy olyan vállalkozásban, amely talán mélyreható következményekkel jár, és általa sokkal jobban megismerhetjük saját magunkat? Esélyünk nyílik rá, hogy megérthessük Isten nyelvét, hogy meghatározhassuk annak a folyamatnak a részleteit, hogyan alakultunk ki mi, emberek. Megtehetem-e, hogy visszautasítom ezt a lehetőséget? Mindig is szkeptikus voltam azokkal szemben, akik azt állították, hogy az efféle pillanatokban hallani vélik Isten akaratát. De ennek a kalandnak az elképesztő fontosságát, illetve az emberiség és az Úr kapcsolatára gyakorolt lehetséges következményeit lehetetlenség volt figyelmen kívül hagyni.
Amikor 1992 novemberében meglátogattam a lányomat Észak-Karolinában, egy hosszú délutánt töltöttem imádkozással egy kis kápolnában, útmutatást kérve az igazgatói pozíció elvállalása ügyében. Nem „hallottam” Istent beszélni - valójában még sosem volt részem ebben az élményben. De azokban az órákban, amelyek végül az engem meglepetésként érő esti istentiszteletbe torkolltak, békére leltem. Néhány nappal később pedig elfogadtam az ajánlatot.
A következő tíz év maga volt az élmények vad hullámvasútja. A humán genom projekt eredeti célkitűzései hihetetlenül ambiciózusnak bizonyultak. Fontos mérföldköveket állítottunk magunk elé, és számon is kértük magunkon, hogy elértük-e részleges céljainkat. Voltak pillanatok, amikor frusztrációnk az egekbe szökött. Például amikor az előzetes tesztek során reménykeltőnek tűnő módszerekről kiderült, hogy nagyobb léptékben alkalmazva látványosan kudarcot vallanak. Tudományos tanácsadó testületünk tagjai között időnként ellentétek alakultak ki - ilyenkor az én feladatom volt, hogy közvetítőként csillapítsam a kedélyeket. Egyes kutatóintézetek nem bírták tartani az általunk diktált tempót, így el kellett búcsúznunk tőlük - ami persze egyáltalán nem tetszett a vezetőiknek. Ugyanakkor diadalmas pillanatokban is volt részünk, amint sikerült elérnünk az ambiciózus célokat, és sokasodni kezdtek az általunk elért eredmények orvosi alkalmazásának lehetőségei. 1996-ra már készen álltunk, hogy elkezdjük tesztelni a humán genom tényleges, nagy léptékű szekvenálását.
Ehhez egy olyan módszert használtunk, amely technikailag összehasonlíthatatlanul fejlettebb, egyúttal sokkal költséghatékonyabb volt, mint az, amelyet 1985-ben használtunk a cisztás fibrózis génje utáni hajtóvadászatban. A projekt egy meghatározó pillanatában mi, a nemzetközi konzorcium vezetői elhatároztuk, hogy bárki csak akkor vehet részt a kutatásban, ha azonnal hozzáférést biztosít az adataihoz az egész világ számára. Megállapodtunk abban is, hogy semmiféle szabadalmat nem nyújtunk be a DNS egyetlen részletére sem. Nem hagyhattuk, hogy azok a kutatók, akik világszerte fontos orvosi problémák megoldásán fáradoznak, ne férhessenek hozzá szabadon a projekt eredményeként megismert adatokhoz.
A következő három év rendkívül gyümölcsözőnek bizonyult. 1999-ben pedig már készen álltunk, hogy jelentősen felgyorsítsuk a szekvenálási folyamatot. Ekkor azonban újabb nehézséggel szembesültünk. A teljes emberi genom szekvenálása iránt korábban nem érdeklődött a privát szektor, mivel kereskedelmi vállalkozásként nem tűnt kifizetődőnek. A feltárt információk értéke azonban napról napra nyilvánvalóbbá vált, a szekvenálás költsége pedig zuhanni kezdett. Így a közpénzből működtetett humán genom projekt legfőbb kihívójává egy magánvállalkozás vált. Craig Venter, annak a cégnek a vezetője, amely nem sokkal később a Celera nevet kapta, bejelentette, hogy a humán genom nagy léptékű szekvenálására készül. Sok gént szabadalmaztatni fog, az adatokat pedig előfizetéses adatbázisban fogja tárolni, amelyhez csak jókora összeg ellenében lehet majd hozzáférni.
Az elképzelés, miszerint az emberi genomszekvencia magántulajdonná válhat, komolyan nyugtalanított bennünket. Még jobban aggódtunk amiatt, hogy a Kongresszusban kérdezgetni kezdték, mi értelme az adófizetők pénzét olyasmire költeni, amit a privát szektor talán hatékonyabban meg tudna valósítani. Csakhogy a Celera kutatói ekkor meg semmilyen adatot nem tettek nyilvánossá, és az a tudományos stratégia, amelyet Venter megcélzott, nem tűnt valóban teljes és pontos eredménnyel kecsegtetőnek. Ennek ellenére az egyre nagyobb hatékonyságról szóló állítások folyamatosan áramlottak a Celera jól olajozott közkapcsolati gépezetéből. Egyúttal igyekeztek a közpénzből finanszírozott projektet lassúnak és bürokratikusnak beállítani. Mivel a humán genom projekt munkálatain a világ legjobb egyetemein dolgoztak a bolygó legkreatívabb és legelhivatottabb kutatói, ezt elég nehéz volt lenyelni. A sajtó persze imádta az ellenségeskedést. Rengeteg cikket írtak a genom megszekvenálásáért folytatott versenyfutásról, Venter jachtjáról és az én motorbiciklimről. Mekkora badarság! A külső szemlélők többsége nem tudhatta, hogy itt alapvetően már nem arról volt szó, ki fogja gyorsabban és olcsóbban elvégezni a munkát. Ugyanis ekkorra már mind a Celera, mind a közpénzből működő vállalkozás képes volt erre. Valójában az ideák csatájáról volt szó. Vajon a humán genomszekvencia, közös örökségünk kereskedelmi árucikké válik-e, vagy univerzális közkincs lesz?
Ekkor már egy pillanatra sem lassíthattunk. A hat országban működő húsz állami genomkutató központ napi huszonnégy órában dolgozott. Másodpercenként ezer bázispárt fejtettünk meg a hét hét napján, a nap huszonnégy órájában. Így mindössze tizennyolc hónappal később a kezünkben volt az emberi genom 90 százalékának előzetes vázlata. Továbbra is minden adatot naponta nyilvánosságra hoztunk. A Celera eközben szintén hatalmas adatmennyiséget termelt, de ezt elzárva tartották, senki nem férhetett hozzá. Később rájöttek, hogy a nyilvánosan hozzáférhető adatokat is hasznosíthatják. Így a Celera végül felhagyott a munkával, nagyjából az előre eltervezett célok felénél. Később kiderült, hogy a Celera által szekvenált genomadatok több mint fele valójában publikus adatokat tartalmazott.
A „versenyfutást” kísérő figyelem lassan kifejezetten kártékonnyá vált, és azzal fenyegetett, hogy a közvélemény szemében lecsökkenti a végső cél jelentőségét. 2000 áprilisában, amikor mind a Celera, mind a közpénzből működő projekt arra készült, hogy bejelentse a vázlatos genomszekvencia elérését, egy közös barátunkon, Ari Patrinoson, az energiaügyi minisztérium genomprogramjának munkatársán keresztül megkerestem Ventert, és bizalmas találkozót kértem tőle. Ari pincéjében egy sör és néhány szelet pizza fölött Venter és én kidolgoztuk az egyidejű bejelentés terveit.
Így aztán, ahogyan a könyv első oldalain már leírtam, 2000. június 26-án ott találtam magam az Egyesült Államok elnöke mellett a Fehér Ház Keleti szobájában: bejelentettük, hogy elkészült az emberi használati utasítás első vázlata. Feltártuk Isten ábécéjét.
A következő három évben abban a kiváltságban lehetett részem, hogy tovább vezethettem a közpénzből működő projektet. A cél az volt, hogy tovább finomítsuk a vázlatos szekvenciát, kitöltsük a fennmaradt réseket, és a lehető legpontosabb információkat közöljük. Továbbra is nyilvánosan hozzáférhető adatbázisokban helyeztük el az adatokat minden egyes nap. 2003 áprilisában, annak a napnak az ötvenedik évfordulóján, amikor Watson és Crick először publikálták a DNS kettőshélix-szerkezetét, bejelentettük, hogy a humán genom projekt minden célját elértük. Mint a vállalkozás projektmenedzsere, nagyon büszke voltam arra a több mint kétezer tudósra, akiknek a segítségével elértük ezt a rendkívüli eredményt. Hiszem, hogy ezer év múlva is az emberiség egyik legnagyszerűbb teljesítményének fogják tartani.
A Genetic Alliance (Genetikai Szövetség) nevű jótékonysági szervezet azokat a családokat támogatja és lelkesíti, amelyek ritka genetikai betegségekkel küzdenek. Egy általuk szervezett későbbi rendezvényre átírtam az All the Good People (Minden jó ember) című közismert népdalt, hogy jobban megfeleljen az alkalomnak. Mindenki együtt énekelte a refrént.
Ez a dal minden jó emberhez szól,
Minden jó emberhez, aki tagja e családnak.
Ez a dal minden jó emberhez szól,
A közös ügy köt össze minket.
Írtam egy új versszakot is arról, mi mindenen kell keresztülmennie sok családnak, miközben az őket vagy a gyermekeiket megtámadó ritka betegségekkel küzdenek.
Ez a dal azokhoz szól, akik szenvednek,
Az erőtök és elhivatottságotok mindannyiunkat megérint.
Az elszántságotok minket is inspirál,
A bátorságotok miatt tudunk állva maradni.
Végül a dal végére illesztettem még egy versszakot a genomról.
Ez egy használati utasítás, a történelem lenyomata,
Egy orvosi tankönyv, amely mindannyiunk közös öröksége.
Az emberek tulajdona, hiszen az emberek nélkül nem létezhet,
Az emberekért létezik, érted és értem is.
Hívő emberként az emberi genomszekvencia feltárása számomra további jelentőséggel is bírt. Hiszen ez a könyv azon a DNS-nyelven íródott, amelyet Isten beszélt, amikor megteremtette az életet. A csodálat túláradó érzése fogott el, miközben a biológiai tudás legfontosabb írását olvashattam. Ezt a könyvet olyan nyelven írták, amelyet egyelőre alig-alig értünk, és évtizedeket, ha nem évszázadokat fog igénybe venni, mire megértjük a benne foglalt utasításokat. De egy egyirányú hídon keresztül végérvényesen átkeltünk a minden tekintetben ismeretlennek számító túlpartra.
Számos könyvet írtak már a humán genom projektről, talán túl sokat is.2 Egy nap talán én is megírom a sajátomat, és remélhetőleg elegendő belátás lesz bennem ahhoz, hogy ne ismételjem meg a jelenleg elérhető, népszerűsítő beszámolók néhány lélegzetelállító állítását. Ennek a könyvnek viszont nem célja, hogy hosszasan elmélkedjen e rendkívüli élmény tanulságain. Sokkal inkább azt szeretném bemutatni, hogy a modern tudományos ismereteket hogyan lehet összhangba hozni az istenhittel.
Ennek érdekében érdemes alaposan megvizsgálnunk az emberi genom szerkezetét, és összevetnünk sok más élőlény génkészleteivel, amelyeket mára már szintén megszekvenáltak. Amint feltártuk az emberi genom nagy részét, vagyis mintegy 3,1 milliárd DNS-betűt, amelyek 24 kromoszómán foglalnak helyet, számos meglepő tény vált nyilvánvalóvá.
Az egyik meglepetés az volt, hogy a genom milyen kis része felelős a fehérjék kódolásáért. Bár kísérleti és számítástechnikai módszereink korlátozott lehetőségei miatt nem tudtunk egészen pontos becsléseket tenni, mégis úgy tűnt, hogy mindössze 20-25 ezer fehérjekódoló gén található az emberi genomban. Ráadásul ezek a fehérjekódoló gének a teljes DNS-állomány alig 1,5 százalékát foglalják el. Egy évtizeden keresztül arra számítottunk, hogy a humán genomban legalább 100 ezer gént fogunk találni. Sokakat tehát igencsak meglepett, hogy Isten valójában csak rövidke novellákat írt az emberiségről. Ez a felfedezés akkor tűnik igazán sokkolónak, ha belegondolunk, hogy a nálunk jóval egyszerűbb élőlények, például a férgek, a legyek és az egyszerű növények génszámai nagyjából ugyanebbe a tartományba esnek: nekik is körülbelül 20 ezer génjük van.
Néhány megfigyelő ezt a felfedezést valóságos sértésként értékelte az ember összetett, komplex voltára nézve. Lehet, hogy mindeddig csak áltattuk magunkat azzal, hogy különleges helyet foglalunk el az állatvilágban? Nos, valójában nem. A génszám biztosan nem tükrözi valódi komplexitásunkat. Bármilyen becslést veszünk alapul, az ember biológiai összetettsége jelentősen meghaladja például az orsóférgekét, hiszen azoknak csupán 959 sejtjük van, annak ellenére, hogy génjeik száma hasonló. És az is biztosra vehető, hogy eddig egyetlen más élőlény sem szekvenálta meg saját genomját. Összetettségünk biztosan nem a különálló instrukciós csomagok számából adódik, hanem annak módjából, ahogyan ezeket a csomagokat hasznosítjuk. Talán a szervezetünket felépítő alkatrészek szert tettek a multitasking képességére?
Ha ismét a nyelv metaforáját hívjuk segítségül, könnyebben megérthetjük génjeink tényleges működését. Egy átlagos, iskolázott, angolul beszélő ember szókincse nagyjából 20 ezer szó. Ezeket a szavakat arra is használhatjuk, hogy meglehetősen egyszerű szövegeket fogalmazzunk meg segítségükkel - például gépkocsink használati utasítását -, de arra is alkalmasak, hogy ennél sokkal összetettebb irodalmi műveket alkossunk belőlük - például James Joyce Ulyssesét. Hasonló módon a férgeknek, a rovaroknak vagy a madaraknak, úgy tűnik, szintén a 20 ezer génből álló szókincsre van szükségük a normális működéshez, bár ők ezt a forrást nyilván kevésbé kifinomult módon hasznosítják, mint mi.
Ha összehasonlítjuk fajunk különböző képviselőinek génkészletét, a genom egy másik meglepő jellegzetességére is fényt deríthetünk. A DNS szintjén ugyanis 99,9 százalékban mindannyian azonosak vagyunk. Teljesen mindegy, hogy a Föld lakói közül melyik két embert választjuk ki, a hasonlóság mindig ugyanekkora lesz köztük. Vagyis a DNS-elemzés értelmében mi, emberek valóban egy család tagjai vagyunk. Ez a kivételesen alacsony genetikai diverzitás különböztet meg minket a bolygó szinte minden más élőlényétől, amelyekben a genetikai változatosság tízszer vagy akár ötvenszer nagyobb, mint az emberben. Ha egy földön kívüli látogatót ideküldenének azzal a feladattal, hogy vizsgálja meg a bolygó életformáit, sok érdekes dolgot elmondhatna az emberiségről, de nagyon valószínű, hogy a fajunkon belül tapasztalható, rendkívül csekély genetikai változatosságot említené elsőként.
A populációgenetika kutatói matematikai módszereket használnak arra, hogy rekonstruálják az állatok, a növények és a baktériumok populációinak múltját. Miután megvizsgálták a humán genomról szerzett adatokat, arra a következtetésre jutottak, hogy fajunk minden képviselője ugyanazoktól az „alapító atyáktól” származik, akik nagyjából tízezren lehettek, és 100-150 ezer évvel ezelőtt éltek. Ez az információ tökéletesen egybevág a fosszilis leletanyaggal, amely alapján úgy gondoljuk, hogy alapító őseink feltehetőleg Kelet-Afrikában éltek.
A sokféle genom vizsgálatának egy további, rendkívül érdekes hozománya, hogy részletesen is összehasonlíthatjuk fajunk és más élőlények génkészleteit. A számítógép segítségével kiválaszthatjuk az emberi DNS egy meghatározott szakaszát, és megvizsgálhatjuk, hogy van-e hasonló részlet más fajok örökítőanyagában is. Ha egy emberi gén kódoló régióját választjuk ki - vagyis olyan részletét, amely egy bizonyos fehérje felépítéséhez szükséges utasításokat tartalmaz -, és ehhez a génszakaszhoz hasonló DNS-részleteket keresünk más fajokban, szinte mindig fogunk olyan géneket találni más emlősökben, amelyek nagyban hasonlítanak az emberi génhez. Sőt sok génnek lesz felismerhető, bár nem tökéletesen azonos megfelelője a halakban is. Néhány emberi génnek még az olyan egyszerű életformák genomjában is megtaláljuk a megfelelőjét, mint a muslicák vagy az orsóférgek. Néhány különösen megdöbbentő esetben ez a hasonlóság egészen az élesztőig, sőt a baktériumokig nyomon követhető.
Másrészről viszont, ha olyan DNS-szakaszt választunk ki, amely a gének között foglal helyet, jóval alacsonyabb lesz annak valószínűsége, hogy ehhez a szakaszhoz hasonló szekvenciát találunk a távoli rokon élőlények génkészletében. Ami nem jelenti azt, hogy egyáltalán nincsenek ilyen hasonló szakaszok. Ha számítógéppel pontosan végignézzük más fajok genomjait, a nem kódoló DNS-részletek felét párhuzamba állíthatjuk más emlősök DNS-ével, és szinte mindnek megtaláljuk a megfelelőjét a főemlősök genomjában. Az 5.1 táblázat azt mutatja, mekkora valószínűséggel találunk egyezést attól függően, milyen DNS-szakaszokat hasonlítunk össze.
5.1 táblázat. Annak valószínűsége, hogy az emberi DNS-hez hasonló örökítőanyag-szakaszokat találunk más élőlényekben - attól függően, hogy fehérjekódoló vagy a gének között elhelyezkedő DNS-szakaszt veszünk alapul.

5.1 ábra. Ezen az oldalon a törzsfáról alkotott mai elképzelésünk látható: a különböző emlősfajok közti kapcsolatokat egyedül a DNS-szekvenciáik közötti hasonlóság alapján határozták meg. Az ágak hosszúsága a fajok közötti különbség mértékét tükrözi. Például az egér és a patkány DNS-szekvenciája közelebbi rokonságban áll egymással, mint az egér és a mókus génkészlete. Az ember és a csimpánz DNS-e pedig jobban hasonlít egymáshoz, mint az ember és a makákó genomja. A szemben lévő oldalon pedig, érdekes történelmi párhuzamként, Darwin 1837-es jegyzetfüzetének egyik oldala. Az „Azt hiszem” (I think) megjegyzés után ezen Darwin elképzelése látható a fajok törzsfájáról.

Mit jelent mindez? Két különböző szinten is meggyőző bizonyítékkal szolgál Darwin evolúcióelméletéhez, miszerint az élőlények közös őstől származnak, majd a bennük véletlenszerűen megjelenő változatok közül a természetes szelekció válogat. A teljes genomot figyelembe véve a számítógép fel tudja rajzolni a törzsfát, pusztán a különféle élőlények DNS-szekvenciái közötti hasonlóságot figyelembe véve. Az eredmény az 5.1 ábrán látható. Tartsuk szem előtt, hogy ez az elemzés egyáltalán nem veszi figyelembe a fosszilis leleteket, sem a jelenleg élő életformák testfelépítésére vonatkozó megfigyeléseket. Mégis megdöbbentően hasonló eredményekre jut, mint a jelenlegi élőlényeket vagy a kihalt fajokat vizsgáló összehasonlító anatómiai kutatások. Darwin elmélete alapján arra számíthatunk, hogy azok a mutációk, amelyek nem befolyásolják az egyedek életműködéseit - vagyis a „hulladék DNS-ben” létrejött változások idővel megszaporodnak. Ugyanakkor a gének kódoló szakaszaiban történt mutációktól azt várjuk, hogy ritkábbak lesznek. Ezek többsége ugyanis a DNS bázispárjainak kivágódásával jár, és csak nagyon ritkán biztosítanak szelekciós előnyt az egyednek. Márpedig ez szükséges lenne ahhoz, hogy az evolúció során fennmaradhassanak. És pontosan ezt figyelhetjük meg: az utóbbi jelenség a gének kódoló szakaszainak legapróbb részleteire is igaz. Az előző fejezetből emlékezhetünk arra, hogy a genetikai kód túlbiztosított: a GAA és a GAG bázishármas egyaránt a glutaminsavat kódolja. Ez azt jelenti, hogy a kódoló régióban történt egyes mutációk „csendesek” maradhatnak, vagyis a DNS-szekvenciában történt változás nem okoz változást a kódolt aminosavban. Ily módon a mutáció nem is okozhat hátrányt - mondhatni, nem kerül semmibe. Amikor egymáshoz közel álló fajok DNS-szekvenciáit hasonlítjuk össze, a kódoló régiókban sokkal gyakrabban találunk csendes mutációkat, mint olyanokat, amelyek miatt megváltozik a kódolt aminosav is. Darwin elmélete pontosan ezt jósolta. Ha ezeket a genomokat egyenként teremtette volna az Úr - ahogyan egyesek érvelhetnek -, vajon miért találkoznánk ezzel a jelenséggel?
Charles Darwin meglehetősen bizonytalan volt saját elméletét illetően. Talán ezért várt közel huszonöt évet a teória kidolgozása és A fajok eredete megjelentetése között. Darwin biztosan sokszor gondolt arra, bárcsak visszamehetne több millió évet az időben, hogy ténylegesen is megfigyelhesse azokat az eseményeket, amelyeket elmélete leírt. Erre természetesen nem nyílt lehetősége, ahogyan mi sem vagyunk képesek rá. De időgép híján Darwin nehezen képzelhette volna el elméletének meggyőzőbb digitális bizonyítékát annál, mint amit a fajok DNS-einek összehasonlításával kaptunk.
A 19. század közepén Darwin nem tudhatta, mi lehet a természetes kiválogatódás útján végbemenő evolúció mechanizmusa. Mi azonban ma már látjuk, hogy a Darwin által kimutatott variáció a DNS-ben természetes módon létrejövő mutációk útján valósul meg. Ezek a mutációk a becslések szerint nemzedékenként és 100 millió bázispáronként egyszer jönnek létre. (Ez egyébként azt is jelenti, hogy az édesanyánktól és az édesapánktól örökölt kétszer hárommilliárd bázispárban nagyjából hatvan olyan mutáció keletkezik, amelyek egyik szülőnkben sem voltak jelen.)
E mutációk nagy része a genom olyan részleteiben jön létre, amelyek nem létfontosságúak, így csekély következménnyel járnak, vagy egyáltalán nem okoznak semmiféle változást. Azok viszont, amelyek a genom sérülékenyebb és fontosabb területein történnek, általában károsak, ezért hamar eltűnnek a populációból, hiszen csökkentik az egyed sikeres szaporodásának esélyét. Ritka alkalmakkor a véletlenszerűen létrejövő mutációk némi szelekciós előnyt biztosítanak hordozójuk számára. Az új DNS-betűsor így némileg nagyobb eséllyel adódik át a jövőbeli utódoknak. Nagyon hosszú idő elmúltával ezek a ritka, de előnyös események elterjedhetnek a populáció összes egyedében, és végső soron jelentős változásokat okozhatnak az életműködéseikben.
A kutatók időnként működés közben is „tetten érik” az evolúciót, hiszen ma már rendelkezünk azokkal az eszközökkel, amelyek a folyamat nyomon követéséhez szükségesek. A darwinizmus kritikusai gyakran érvelnek azzal, hogy a fosszilis leletanyagban nincs bizonyíték makroevolúcióra - vagyis a fajokban létrejövőjelentős változásokra csak a mikroevolúció - a kis változások - nyomai fedezhetők fel. Az kimutatható, mondják, hogy a pintyek csőrének formája idővel megváltozik, attól függően, hogy milyen táplálékot fogyasztanak, de új fajok megjelenését még senki nem látta.
Egyre valószínűbb, hogy ez csupán mesterséges distinkció. A Stanford Egyetem egyik kutatócsoportja például azt kutatja, hogy a pikóféle halak testpáncélja miért vált olyan sokfélévé az evolúció során. A sós vízben élő pikók testét általában három tucat, sorban álló páncéllemez fedi a fejüktől a farkukig. A világ sok részén előforduló édesvízi pikók azonban elvesztették páncéllemezeik jó részét, mivel az élőhelyükön kevesebb a ragadozó.
Úgy véljük, az édesvízi pikók tíz-húsz ezer éve jelentek meg jelenlegi élőhelyükön, miután a gleccserek az utolsó jégkorszak végén szerte a világon olvadásnak indultak. Az édesvízi pikók genomjainak alapos vizsgálata feltárt egy gént, amelyet EDA-nak neveztek el. Ennek változatai ismétlődően és egymástól függetlenül megjelentek az édesvízi környezetben, és a páncéllemezek elvesztését okozták. Érdekes módon mi, emberek szintén rendelkezünk ezzel az EDA-génnel. Az EDA-génben létrejövő spontán mutációk defektusokat okoznak a hajunkban, a fogainkban, a verejtékmirigyeinkben és a csontjainkban. Nem nehéz belátni, hogy az édesvízi és a sósvízi pikók közötti különbségek továbbfejleszthetők, és ezáltal számos különféle hal jöhetett létre. A makro- és a mikroevolúció közötti különbségtétel tehát meglehetősen önkényesnek tűnik: azok a nagy változások, amelyek egy-egy új faj létrejöttét eredményezik, a kisebb, szinte elhanyagolható változásokból indulnak ki.
Az evolúciót minden egyes nap megfigyelhetjük működés közben, amikor a különféle betegségeket okozó vírusok és baktériumok, illetve a közegészségügyi vészhelyzeteket kiváltó paraziták gyors fejlődését látjuk. 1989-ben annak ellenére kaptam el a maláriát Nyugat-Afrikában, hogy megelőzésképpen beadták nekem az ilyenkor szokásos gyógyszert, a klorokint. Csakhogy a maláriát okozó parazitában véletlenszerűen létrejövő mutációk erős szelekciónak voltak kitéve, amelyet a klorokin sokévi intenzív használata okozott a világnak ezen a táján. Ennek hatására végül olyan kórokozó fejlődött ki, amely már rezisztens volt a gyógyszerre, ezért gyorsan el tudott terjedni. Ehhez hasonlóan az AIDS betegséget okozó HIV-vírus gyors evolúciós változásai is komoly akadályokat gördítenek az oltás kifejlesztése elé. Elsősorban ezek a változások tehetnek arról is, hogy az AIDS-gyógyszerekkel kezelt betegek előbb-utóbb gyakran visszaesnek. Szintén közismert, hogy a szakemberek attól tartanak, a madárinfluenzát okozó H5Nl-es vírustörzs esetleg világméretű influenzajárványt okozhat. Félelmüknek az az alapja, hogy a jelenlegi vírusváltozat, amely a madarak, illetve a fertőző madarakkal kapcsolatba került emberek számára már most is halálos veszélyt jelent, nagy valószínűséggel továbbfejlődhet, és képes lehet akár emberről emberre is terjedni. Biztosan állítható, hogy nemcsak a biológia, de az orvostudomány sem érthető meg az evolúcióelmélet figyelembevétele nélkül.
Az evolúcióelmélet alkalmazása a pikókra egy dolog - de mi a helyzet velünk, emberekkel? Darwin kora óta a legkülönfélébb világnézetű emberek igyekeztek megérteni, hogy a biológiai és az evolúciós felfedezések hogyan alkalmazhatók az állatok egy különleges csoportjára, az emberekre.
A genom vizsgálata szükségszerűen arra a következtetésre vezet minket, hogy mi, emberek és minden más élőlény egy közös ősből fejlődtünk ki. A bizonyítékok egy része az 5.1 táblázatban látható, amelyben az ember és más élőlények genomjai közötti hasonlóságot tüntettük fel. Ez önmagában természetesen nem bizonyítja a közös ős létezését. Kreacionista nézőpontból az efféle hasonlóságok csak arra utalnak, hogy Isten jól működő tervezési alapelveket használt újra meg újra. De mint látni fogjuk - és ahogyan már utaltunk is rá, amikor a gének fehérjekódoló régióiban létrejövő „csendes” mutációkról ejtettünk szót -, a genom részletes vizsgálata ezt az értelmezést gyakorlatilag tarthatatlanná tette nem csupán a többi élőlény, de az ember esetében is.
Első példaként vegyük az emberi és az egérgenom összehasonlítását, már csak azért is, mert mindkettő nagy pontosságú szekvenciája rendelkezésünkre áll. A két genom teljes mérete nagyjából ugyanakkora, és a fehérjekódoló gének készlete is figyelemre méltó hasonlóságot mutat. A közös ős további egyértelmű jelei is azonnal nyilvánvalóvá válnak, amint a részletekre pillantunk. Például a gének sorrendje az emberi és az egérkromoszómákon a DNS jó részén azonos. Tehát ha az emberi A, B és C géneket ebben a sorrendben találjuk az emberi kromoszómán, akkor az A, a B és a C gének egérmegfelelői jó eséllyel ugyanebben a sorrendben fognak megjelenni a rágcsáló kromoszómáján - bár a gének közötti távolság némileg változhat (5.2 ábra). Egyes esetekben ez az egyezés igen nagy génszakaszok teljes hosszára is fennáll; például az emberi 17-es kromoszómának gyakorlatilag az összes génje megtalálható az egér 11-es kromoszómáján. Persze közbevethetnénk, hogy a gének sorrendje kritikus fontosságú abból a szempontból, hogy aktiválódásuk időzítése megfelelő legyen, ezért a tervező ugyanezt a sorrendet alkotta meg számos, különálló teremtési aktus alkalmával. Jelenlegi molekuláris biológiai tudásunk szerint azonban nincs bizonyíték arra, hogy ez a megkötöttség ilyen hosszú kromoszómaszakaszokra is érvényes lenne.
A közös ős még ennél is meggyőzőbb bizonyítékával szolgál az ősi ismétlődő elemeknek (ancient repetitive elements, ARE) nevezett DNS-szakaszok vizsgálata. Ezek azokból az „ugráló génekből” származnak, amelyek képesek lemásolni és beilleszteni magukat a genom különböző pontjaira, általában anélkül, hogy ez befolyásolná a génműködést. Az emlősök genomjai telis-tele vannak ezekkel az ősi szakaszokkal: az emberi génkészlet nagyjából 45 százaléka efféle genetikai hulladékból áll. Ha az emberi és az egérgenom szakaszait egymás mellé illesztjük az azonos sorrendben jelen lévő génmegfelelők szerint, gyakran az azonos ARE-kat is azonosíthatjuk a két genom nagyjából azonos helyein (5.2 ábra).
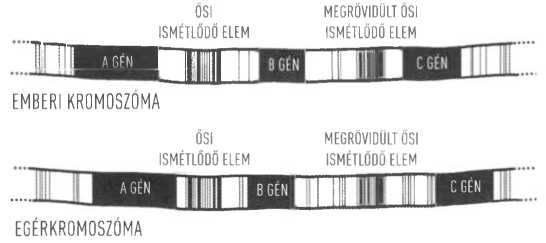
5.2 ábra. A gének elhelyezkedése a kromoszómákon gyakran követi ugyanazt a sorrendet az egerekben és az emberekben, habár a gének közötti pontos távolság némileg változhat. Tehát ha az emberi A, B és C géneket ebben a sorrendben találjuk az emberi kromoszómán, akkor jó esélyünk van arra, hogy az A, a B és a C gének egérmegfelelői ugyanebben a sorrendben fognak megjelenni az egérkromoszómán. Mivel most már az emberi és az egérgenom teljes szekvenciája rendelkezésünkre áll, azonosítani tudjuk a gének közötti szakaszokban az úgynevezett „ugráló gének” sok maradványát. Ezek a transzpozonnak is nevezett elemek képesek voltak bemásolni magukat a genom véletlenszerű pontjaira - és ezt ritkán még ma is megteszik. A DNS-szekvencia elemzése kimutatta, hogy a transzpozonok némelyike az eredeti ugráló génekhez viszonyítva számos mutációt gyűjtött össze az idők során, vagyis igen idősnek tűnnek. Ezeket gyakran hívják ősi ismétlődő elemeknek (ancient repetitive elements, ARE). Érdekes módon ezek az ősi elemek gyakran hasonló helyeken találhatók az egér és az ember genomjában - mint a fenti példán az A és a B gén között helyet foglaló ARE. Különösen érdekesek azok az esetek, amikor az ARE egy szakasza levágódott a beépülés pillanatában. Ezzel elvesztette DNS-e egy részét és minden esélyét arra, hogy működhessen a jövőben (amint azt példánkban a B és a C gén között látjuk). Amikor olyan megrövidült, ősi elemeket találunk az ember és az egér genomjában, amelyek pontosan ugyanannál a bázispárnál vágódtak el, és ugyanoda épültek be a kromoszómába, ez meggyőzően bizonyítja, hogy a beépülés az ember és az egér egy közös ősében ment végbe.
Ezek közül néhány talán elveszett egyik vagy másik fajban. De sok közülük megőrizte azt a pozícióját, ahová az ősi elemek annak idején beépültek a közös ős genomjába, és a leszármazottak azóta is hordozzák őket. Természetesen felvethető ellenérvként, hogy ezek valójában működő elemek, amelyeket a Teremtő helyezett el jó okkal a genomban. Az pedig, hogy „hulladék DNS-sé” degradáljuk őket, csak tudatlanságunk jelenlegi mértékéről árulkodik. És valóban: néhányuk talán tényleg szabályozó szerepet tölt be. Ugyanakkor bizonyos példák komolyan megkérdőjelezik az isteni beavatkozás lehetőségét. A transzpozíció folyamata - amikor a transzpozon átkerül egy másik genom-helyre - gyakran károsítja az eredeti ugráló gént. Az emberi és az egérgenom tele van olyan ARE-kkal, amelyek megrövidültek, amikor beépültek új helyükre, és ezzel működésképtelenné váltak. Sok esetben azonosíthatjuk a lefejezett és teljesen haszontalanná vált ősi elemeket az emberi és az egérgenom párhuzamba állítható helyein (5.2 ábra).
Ha nem feltételezzük, hogy Isten azért helyezte a genom meghatározott helyeire a lefejezett, ősi ismétlődő elemeket, hogy összezavarjon és félrevezessen minket, gyakorlatilag kétségbevonhatatlan a következtetés, miszerint az emberek és az egerek egykor közös őssel rendelkeztek. Vagyis a genommal kapcsolatos legújabb adatok szinte leküzdhetetlen kihívások elé állítják azt, aki mindenáron ragaszkodik hozzá, hogy minden egyes faj ex nihilo teremtetett.
Az ember elhelyezkedését az evolúciós törzsfán az is alátámasztja, ha összehasonlítjuk genomunkat legközelebbi élő rokonaink, a csimpánzok génkészletével. A faj genomszekvenciáját már megfejtették, és kiderült, hogy a csimpánz és az ember a DNS szintjén 96 százalékban azonosak.

5.3 ábra. Az emberi és a csimpánzkromoszómák, más néven kariotípusok. Vegyük észre a kromoszómák méretében és számában megmutatkozó jelentős hasonlóságot. Egyetlen fontos különbség van: a 2-es emberi kromoszóma mintha úgy jött volna létre, hogy két független csimpánzkromoszóma (az ábrán 2A-val és 2B-vel jelöltünk) összeolvadt a végüknél. (A valóságban ez természetesen nem így történt, hiszen a csimpánz nem őse az embernek, hanem közös őstől származnak mindketten.)
A közeli rokonság további bizonyítékaival szolgált az emberi és a csimpánzkromoszómák anatómiájának részletekbe menő tanulmányozása. A kromoszómák a DNS látható megtestesülései. Fénymikroszkóppal is vizsgálható, ahogyan a sejt osztódik. Minden kromoszóma gének százait tartalmazza. Az 5.3 ábrán megfigyelhetjük az emberi és a csimpánzkromoszómákat párokba rendezve. Az embernek huszonhárom pár kromoszómája van, míg a csimpánznak huszonnégy. Kromoszómaszámaink különbsége feltehetőleg abból adódik, hogy két ősi kromoszóma valamikor a régmúltban összeforrt, hogy létrehozzák a 2-es emberi kromoszómát. A gorillának és az orangutánnak ugyancsak huszonnégy pár kromoszómája van, és ezek nagyon hasonlítanak a csimpánz kromoszómájára. Ez is arra utal, hogy az emberi kromoszóma biztosan két ős összeolvadásából jött létre.
A legutóbbi időkben, miután a teljes emberi genomszekvenciát meghatároztuk, lehetőségünk nyílt megkeresni, hogy ez a kromoszóma-összeolvadás pontosan hol történhetett. A 2-es kromoszóma hosszú karján található hely DNS-szekvenciája egészen lenyűgöző. Anélkül, hogy túlságosan elmerülnénk a technikai részletekben, fogadjuk el kiindulópontként, hogy minden főemlős kromoszómáinak végén különleges DNS-szekvencia található. Ezek a szekvenciák általában sehol másutt nem fordulnak elő. Pontosan ott mégis megjelennek, ahol ezt az evolúcióelmélet alapján várhattuk: az összeforrt 2-es kromoszóma közepén. A fúzió, amely akkor történt, amikor az ember leszármazási vonala elvált a ma élő többi főemlős ágától, itt hagyta DNS-lenyomatát. Nagyon nehéz lenne megmagyarázni ezt a megfigyelést anélkül, hogy egy közös őst feltételeznénk.
És ezzel még nincs vége az ember és a csimpánz közös ősére utaló bizonyítékok sorának. Itt vannak ugyanis a genom különös, „pszcudogénnek” nevezett elemei. Ezek a gének szinte minden olyan tulajdonsággal rendelkeznek, mint a DNS működőképes információcsomagjai, kódsorrendjük azonban egy vagy több olyan hibát tartalmaz, amely értelmezhetetlenné teszi a bennük tárolt információt. Ha összehasonlítjuk az ember és a csimpánz genetikai állományát, egyes génekről kiderül, hogy teljesen működőképesek az egyik fajban, a másikban viszont nem, az idők során ugyanis egy vagy több bázistörlődéssel járó mutáció történt bennük. Az emberi kaszpáz-12-nek nevezett gén például számos bázisvesztéssel járó változást hordoz, bár ugyanott helyezkedik el, mint a csimpánzé. A csimpánz kaszpáz-12 génje ezzel szemben tökéletesen működik - ahogy szinte minden más emlős, köztük az egér homológ génjei is hibátlanul funkcionálnak. Ha az Úr természetfeletti erejét használva külön teremtette volna meg az embereket, miért vesződött volna azzal, hogy ilyen működésképtelen gént illesszen genomunk megfelelő helyére?
Ma már ahhoz is eleget tudunk, hogy megmagyarázzuk néhány, köztünk és legközelebbi rokonaink között fennálló mechanikai különbség eredetét, amelyek némelyike fontos szerepet játszott emberré villásunkban. Például úgy tűnik, hogy az MYH16 jelű, az állkapocsizmokban jelen lévő, fehérjét kódoló gén az emberben pszeudogénné változott. A többi főemlősben azonban továbbra is fontos szerepet játszik az állkapocs izomzatának fejlődésében és megerősödésében. Nyilvánvaló, hogy a gén inaktiválása csökkentette az ember állkapcsának izomtömegét. A legtöbb főemlősnek ugyanis teljes testtömegéhez viszonyítva nagyobbak és erősebbek az állkapocsizmai, mint az embernek. Az ember és az emberszabású majmok koponyájának egyik feladata, hogy rögzítést biztosítson az állkapocsizmok számára. Elképzelhető, hogy állkapocsizmaink gyengülése paradox módon megteremtette annak lehetőségét, hogy az agykoponyánk felfelé növekedjen, és így helyet adjon a nagyobb agynak. Ez persze csak spekuláció. Számos további genetikai változás is szükséges volt ahhoz, hogy agykérgünk, amelyet sokan az ember és a csimpánz közötti legfontosabb különbségnek tartanak, sokkal nagyobbra nőjön, mint az emberszabásúaké.
Egy újabb példa a FoxP2 gén, amelyet mostanában élénk tudományos érdeklődés övez, ugyanis talán szerepe van a nyelvi fejlődésben. A FoxP2 története azzal kezdődött, hogy találtak egy angol családot, amelynek tagjai már három nemzedék óta súlyos beszédproblémákkal küzdenek. Komoly kihívást jelentett számukra, hogy a nyelvtani szabályoknak megfelelően rendezzék egymás után a szavakat, és hogy megértsék a többszörösen összetett mondatokat. Az is gondot okozott nekik, hogy megfelelően mozgassák az izmokat a szájukban, az arcukban, illetve a hangszálaik környezetében annak érdekében, hogy jól ejtsenek ki egyes hangokat.
A genetikai nyomozás mesteri példájaként a kutatóknak sikerült azonosítaniuk egyetlen hibás betűt a családtagok 7-es kromoszómáján lévő FoxP2 génben. Igazi meglepetést okozott a tény, hogy egyetlen gén szinte elhanyagolható hibája ilyen súlyos nyelvi hiányosságokat képes okozni, anélkül, hogy a mutációnak bármilyen más ismert következménye lenne.
A meglepetést csak fokozta, amikor kiderült, hogy a FoxP2 gén szekvenciája kifejezetten állandó szinte az összes emlősben. Szinte az összesben. Drámai kivételként bennünk, emberekben két lényeges változás is történt a gén kódoló régiójában, méghozzá valószínűleg százezer évvel ezelőtt. A felfedezések alapján azt a hipotézist fogalmazták meg a tudósok, hogy a FoxP2 génben végbement, evolúciós léptékben viszonylag új változások valamilyen módon hozzájárulhattak, hogy az ember megtanult beszélni.
Ezen a ponton az istentelen materialisták talán igazolva látják saját világnézetüket. Ha az ember pusztán mutációk és természetes szelekció révén fejlődött ki, akkor kinek lenne szüksége Istenre, hogy megmagyarázza létünket? Erre csak azt tudom felelni: nekem. A csimpánz és az ember génszekvenciáinak összehasonlítása, bármilyen érdekes is, semmit nem mond arról, hogy mi tesz bennünket emberré. Úgy vélem, önmagában a DNS-szekvencia - még ha ki is egészítjük a biológiai funkció hatalmas tárházával - soha sem lesz képes megmagyarázni bizonyos emberi tulajdonságokat, például a morális törvény ismeretét és az istenkeresés univerzalitását. Hiába szabadítjuk meg az Urat a teremtés egyes terheitől, attól még ő marad mindannak a forrása, ami különlegessé tesz minket, embereket és magát az univerzumot is. A genetikai felfedezések mindössze arról tanúskodnak, hogy Isten hogyan vitte végbe a teremtést.
A genomvizsgálat területéről bemutatott példákat továbbiakkal is kiegészíthetnénk, amelyek ennél a kötetnél százszor hosszabb könyveket is megtöltenének. Ezek együttesen olyan molekuláris bizonyítékát adják az evolúcióelméletnek, amely gyakorlatilag minden, a szakmáját gyakorló biológust meggyőzött róla, hogy Darwin elmélete a variációkról és a természetes kiválasztásról megkérdőjelezhetetlenül helytálló. Valójában a hozzám hasonlóan genetikával foglalkozók számára szinte lehetetlen lenne összehasonlítani azokat a hatalmas adattömegeket, amelyek a genomvizsgálatokban születnek, ha nem vehetnénk figyelembe Darwin elméletének alapelveit. Ahogyan Theodosius Dobzhansky, a 20. század egyik vezető biológusa - egyébként hívő ortodox keresztény - megfogalmazta: „A biológiában semminek nincs értelme, kivéve, ha az evolúció fényében vizsgáljuk.”3
Ugyanakkor az is egyértelmű, hogy az evolúcióelmélet miatt a vallásos közösségek igen sok tagja érezhette magát meglehetősen kellemetlenül az elmúlt százötven évben. És semmi jele annak, hogy ez az ellenállás belátható időn belül enyhülne. A hívők mégis bölcsebben tennék, ha nem hagynák figyelmen kívül a tudományos bizonyítékok sorát. Ezek ugyanis mind azt támasztják alá, hogy a Föld valamennyi élőlénye - beleértve az embert is - rokonságban áll egymással. A bizonyítékok tömegét tekintve zavarba ejtő, hogy az Egyesült Államokban milyen csekély előrelépés történt az evolúció elfogadása terén. Részben talán az „elmélet” szó egyszerű félreértése okolható a problémáért. A kritikusok imádják hangsúlyozni, hogy az evolúció „csupán egy elmélet”. Ez a kijelentés azonban érthetetlen a tudósok számára, hiszen ők teljesen más értelemben használják a kifejezést. Az én Funk & Wagnalls-szótáram az alábbi két lehetséges meghatározást kínálja az „elmélet” szóra: „1. Valamiről alkotott spekulatív és feltevésen alapuló nézet; 2. a tudomány, művészetek stb. mögött rejlő alapelvek: például zeneelmélet, matematikai elmélet.” A tudósok ezt a második jelentést használják, amikor az evolúcióelméletről beszélnek - csakúgy, mint amikor a gravitációs elméletről vagy a fertőző betegségek terjedését magyarázó kórokozó-elméletről esik szó. Ebben a kontextusban az „elmélet” szóval nem bizonytalanságot akarnak kifejezni; erre a célra a kutatók inkább a „hipotézis” szót használják. A köznapi, laikus nyelvezetben ugyanakkor sokkal tágabb értelemben használjuk az „elmélet” szót, amint arra a Funk & Wagnalls-szótár első meghatározása is utalt: „Az elméletem szerint Bill bele van esve Marybe”, vagy „Linda elmélete szerint a komornyik tette”. Nyelvünkből sajnos hiányoznak a szükséges finomságok, hogy különbséget tudjunk tenni e két értelmezés között. Ez az egyszerű félreértés egy szó jelentése kapcsán sokat ártott a tudomány és a hit viszonyáról és az élőlények rokoni viszonyairól folytatott vita hangnemének.
Ha az evolúció elmélete helytálló, marad-e még hely az Isten számára? Arthur Peacocke, az elismert molekuláris biológus, akiből később anglikán lelkész lett, sokat írt a biológia és a hit határterületeiről. Nemrég megjelentetett egy könyvet Evolúció: a hit álruhába bújt barátja? címmel.4 Ez az érdekes cím azt ígéri, hogy talán kiengesztelhetek a felek. De vajon nem csak kikényszerített házasságról van szó az összeegyeztethetetlen világnézetek között? Miután kifejtettük érveinket Isten létének lehetősége mellett, és megismerhettük az univerzum, illetve a földi élet eredetéről alkotott tudományos nézeteket is, esetleg mégis rátalálhatunk a boldog és harmonikus szintézisre?
III. HIT A TUDOMÁNYBAN HIT ISTENBEN