6
CONSCIOUSNESS
Experience and Animal Life
The previous chapter described an animal, a primate, whose living patterns and mind have been permeated by culture. This has many consequences for what those animals can do, for their role in the world. Another side of all this is how it feels—what it feels like to be the outcome of an evolutionary and historical process of that kind. That is the topic of this chapter: human experience, our form of consciousness. Human conscious experience is a joint product of our animal nature and the infusion of our lives with culture.
The starting point is the fact that there’s a way the events of our lives, or some of them anyway, feel from the inside. There’s something it feels like to be us. This feature is now often referred to as “consciousness.” That is a broad use of that term, where consciousness does not require a reflective, “here I am” kind of awareness, or anything like that. The term “consciousness” now tends to be applied to even the simplest kinds of feeling; you are conscious if it feels like anything at all to be you. The same phenomenon can be called “felt experience,” “subjective experience,” or “sentience” (though that word brings with it particular connotations of pleasure and pain). Future work in this area will probably develop new and better ways of describing and categorizing what is going on, so I won’t worry much about the terms. I’ll generally use “felt experience” for the broad phenomenon, although most of the chapter is about experience in us, where the word “consciousness” is more clearly apt.
Much is unknown here and speculation is unavoidable. I won’t give arguments for my basic picture (those can be found in my other work), but will outline and apply it.
I see felt experience as widespread in animal life. Some of the features responsible for it have entered the scene in earlier chapters of this book. I looked at the evolution of action and its control by sensing. An animal is a nexus in which sensing and action meet, with a mass of causal lines coming in and radiating out. That feature of animal life is important for this book’s theme of action and transformation, and it also creates a system with a point of view. It creates a system to whom things seem a certain way—a system that is a “whom,” in fact, rather than just an object in the world.
The history of such systems starts with single-celled organisms, moves through the evolution of controlled movement, and on through the rise of animals. In animals, large numbers of cells work together to create movement, sensory systems become organized arrays of cells, and coordination of these new wholes is made possible by nervous systems. Nervous systems may have first arisen largely as a means to “pull bodies together,” to make action possible in these unwieldy new objects. When animals became more active, during the Cambrian period half a billion years ago, nervous systems together with richer senses achieved an opening-up to the world. This was a time of the focusing of subjectivity, resulting from the demands of a life featuring more engagement with other animals, and a need for decision-making in real time.
This feature that I call the “focusing of subjectivity” arises from the integrating of information from different senses, the use of these senses alongside traces of the past in memory, and, in addition, the establishment of loops between sensing and action of the kind that have been encountered in several chapters of this book. Mobile animals have to process information they get from the senses in a way that takes account of the consequences of their own actions for what they sense. In this way, a tacit “self-other” distinction is embedded in a mobile animal’s dealing with the world.
Some aspects of felt experience arise as natural consequences of the animal way of being, consequences of the fact that in animals, evolution built systems that are very much “whoms,” as I said a moment ago. All this is the first element of my picture of the biology of experience. A second element is a view about those internal controllers, nervous systems, themselves. People often think of a nervous system as a network a bit like an old-fashioned telephone exchange, with connections and relays. It’s a collection of discrete units, neurons, whose activity is firing and making others fire: neuron A’s firing affects neuron B, which affects C, and so on. In this view, a nervous system’s job could faithfully be done, at least in principle, with a very different sort of device, one not made of living cells, as long as it formed a network with the same connections between its parts.
More of what goes on in our brains matters than that, most likely. A nervous system is a physically unusual object, and those peculiarities are probably important to the explanation of how felt experience comes to exist. As well as containing those node-to-node connections I mentioned a moment ago, brains contain rhythms and patterns of electrical oscillation that span the whole system, or large parts of it. I’ll refer to these as “largescale dynamic patterns.”
An example is the oscillation in electrical activity in our brains that is picked up with an EEG (electroencephalogram) device, a mesh of electrical detectors that sits on the head. Some of what an EEG reading tracks is a great mass of those familiar neuron-to-neuron effects, the “spiking” or “firing” of cells, but it’s also picking up slower, less localized patterns in the brain’s electrical activity.
Why might large-scale patterns of this kind matter to felt experience? I want to approach this question by looking at the evolutionary side, and at several different kinds of animals. Some decades ago, Francis Crick (of DNA fame), Christof Koch, and others began to argue that some of these large-scale dynamic patterns have a role in human visual experience, because they help us to integrate the various features that we perceive—matching colors to shapes in the objects we see, for example. The large-scale rhythmic patterns that Crick and Koch wrote about are not human peculiarities, though; they are all over the animal kingdom, including in animals that can’t see at all. They don’t appear in the same form everywhere, and they have different speeds, with different triggers, and so on. But they are found all over the animal world. Why are they there?
Back in the 1960s, the neurobiologist L. M. “Mac” Passano (often working with C. B. McCullough) described large-scale electrical rhythms in simple jellyfish-like animals called Hydra. Passano conjectured that spontaneous rhythmic activity might be a feature of very early nervous systems, and might figure in behaviors such as feeding. Once a spontaneous pattern of activity of this kind has been established, that pattern can be modulated by sensory events and other goings-on. It becomes a platform on which events can be registered and responses can be coordinated. Passano suggested that these ancient rhythms could have evolved into the more elaborate electrical patterns seen in other animals. The picture is one where rhythmic activity starts up early in animal evolution. This activity recurs, restarts, in each individual as their nervous system comes into existence, and also continues on, species to species, over the millennia. The rhythms continually change in the individual (as they sleep and wake, for example) and also change on an evolutionary timescale, reaching new forms in larger nervous systems like ours.
The first thing this picture does, in the project of explaining how felt experience can be a biological phenomenon, is change our view of what nervous systems are like, and what kind of activity goes on in them. Brain processes include network-like, unit-to-unit flows of information, and they also include these more diffuse, large-scale patterns of electrical activity, continually shifting and being modulated by the events the animal encounters. The idea that all this feels like something “from the inside,” to the animal itself, is not at all surprising to me. It’s not that these rhythms and other large-scale patterns are all that matter, but they are part of what makes nervous systems into special physical objects, objects that organize nature’s energies in a unique way.
Connections can also be drawn between this picture of nervous systems and the feel of experience itself. Again stepping back in time a little, Passano gestured toward a kind of power that comes from organizing the activity of a nervous system in this manner. It can make the system responsive to very subtle signals, such as tiny glimpses in vision. When a largescale pattern of activity is set up and then modulated by events as they happen, this can also give rise to a kind of natural integration, a bringing-together, of processing. I said a moment ago that the brain’s large-scale electrical patterns will be affected by what is coming in through the senses—by sight, by touch—and they will also be affected by other things going on inside the animal: Is it running low on food? When a total state of activity is being perturbed in this way, many different influences can have effects together, in concert, on that whole. This may be useful for the animal in working out what to do, and it also has links to the nature of felt experience itself. A feature of ordinary experience that is sometimes underplayed, even denied, in recent scientific discussions is the fact that at each moment, our experience has many facets that are present together. Even if you are focusing pretty intently on one thing—this book, the road you’re driving on, or a person you’re talking with—the total “feel” of experience is affected by hints from much else. It’s affected by other things you are sensing, by your mood, by how your body is arranged, your energy level, and more. All these are part of that moment’s gestalt, even if their influence is very slight. This feature of experience is just what we’d expect to find, given the view of brain activity I’ve been outlining over the last page or two, in which large-scale patterns of activity are being modulated by many different influences at once.
This oscillatory activity in the brain might be something that arises almost automatically when a nervous system comes to exist—at least when those nervous systems are made out of the materials in animals on Earth—or it might be something that had to be built and installed, to help animals do what they do. It could be some of both of these; there’s an unknown interplay here between the tasks or functions of early nervous systems and the inherent tendencies of the materials that came to hand.
Think back for a moment to the first part of my sketch of the biology of felt experience, and the idea of a “focusing of subjectivity” as animals became more active. Some animals acted more, sensed more, and developed richer links between the two. When we initially picture this, it can sometimes seem that a simple robot could do it all just as well as an animal: all it needs is good sensors (such as eyes), a memory, a way to integrate processing, and so on. I suggest that the role played by our animal natures is greater than this. Not just any set of control devices could give rise to experience, and not just any set of control devices could enable animals to do what they do. I am making a bet here about a link between what animals do and what is inside them, the internal resources by which the animal way of being becomes possible.
That is my picture of the biology of experience. A lot is missing, and it’s more of a sketch than a worked-out theory. But it is different from other such sketches, and it makes contact with what we know about animal life, with what experience actually feels like, and with some surprising features of nervous systems across the animal kingdom. This is a view in which felt experience is not restricted to brains like ours (those of primates, or mammals). Instead, it is probably found in a great many animals, including some who are far from us in evolutionary terms: octopuses, crabs, bees. The list of sentient animals is probably long. In addition, the presence of felt experience is probably a “graded” phenomenon, with a lot of gray-area cases, rather than always being a yes-or-no matter. The history of animal life probably won’t contain sharp lines separating the animals that have experience from the ones that don’t. Animals like earthworms, scallops, and anemones may end up in a liminal region, where experience is neither fully present nor fully absent.
If this view is right, then although artificial minds may be possible in the future, they would have to be different in their physical makeup from computers of the kind we have now. It won’t be possible to build a new conscious being just by devising a very complicated program that might run on a present-day computer. The situation is different with another technology that is developing very quickly; this is the technique of building “neural organoids” by growing colonies of animal (sometimes human) stem cells that self-organize into a small living model brain. This is not a road to a wholly artificial mind, because the system is grown from cells, but those living parts could be combined with nonliving parts, and the prospect of a new kind of sentient being opens before us. If they do become sentient, then especially in the earlier stages of this work, what might experience be like for these creatures? Does their experience seem likely to be positive?
I gave a talk about biology and AI once, and ended the discussion saying that parts of my talk could be seen as an invitation to build new kinds of hardware. The next day I woke up and thought: Is that such a good idea? Perhaps a natural barrier exists between the “I” side of AI—intelligence—and felt experience, at least within the sorts of machines that exist now, and that are relatively easy to build. We might try to overcome the barrier with new hardware, but should we? A barrier of this kind might not stop AI systems from causing us all sorts of trouble in the future, but if they are not sentient, it will simplify the ethical questions about how we should deal with them. Perhaps the barrier is a blessing.
In any case, all this gives us a picture of the biology of felt experience, one we can hold in our minds as we start to think about a special case—us.
Human Minds
Part of the story of human consciousness is the story of animal experience in general. Another part involves the side of our lives described in the previous chapter of this book: culture, language, technology, and sociality. Our minds have been drenched in culture. What difference does this make to an experiencing brain?
The biggest factor is perhaps language itself. Technology of many kinds is important, but the role of language is broader, older, more ubiquitous. That is what I will look at here.
Language starts as a pattern of social interaction, and it becomes a tool for thought. There’s still a good deal of uncertainty about how deep its role is. Language has sometimes been seen as the medium for complex processing, but views like this underestimate what nonlinguistic animals can do. Rats make use of map-like representations that enable them to not only remember paths they’ve traveled, but also consider possible new ones. A 2022 paper found that crows could learn to generate “recursive” patterns, in which one structure is embedded within another, in the way also seen in mathematical systems and language. In our own case, Stanislas Dehaene, a French neuroscientist who has influenced many of my discussions of the mind, recently argued, together with a group of colleagues, that human minds make use of a number of “inner codes” that are quite distinct from the languages we speak.
Nonetheless, language seems to have a cluster of important roles in our minds, some overt, some more subtle. I’ll start with some of the subtle ones, where we find that the way language has become physically part of us has left us with some clues about its role, and about what we’d be like without it.
Language, especially speech, is lateralized in our brains; it is among the skills and abilities that differ across the left and right sides. Many parts of our brains are duplicated across left and right, including the large cerebral hemispheres. The consequences of this duplication are seen especially in individuals who have had the main connector between the two hemispheres cut, to treat severe epilepsy. These are the so-called “split-brain” patients. I have used these remarkable cases to work through some puzzles about animal and human minds in earlier books. Much of what we’ll be looking at here is less exotic; ordinary people who’ve not had separations imposed between the two sides can still give us information about differences between left and right.
In most humans, language—or at least speech—is mainly controlled by the left hemisphere. The left side of the brain also controls the right hand and receives sensory information mostly from the right side, and vice versa. Most humans (around 90 percent) are also right-handed, which means the left side of the brain has a “dominant” role. That idea of a “dominant” side should not be overinterpreted, as suggesting something about overall control and personality; for now, it involves just how different limbs and especially hands are chosen to do things, such as grasping and pointing.
Are left-handers using the right side of their brain for speech? Mainly no, it seems. Seventy percent or more use the left side, as right-handers do. The reason is not known.
The left hemisphere not only controls language but also specializes in the production of sequence-based behaviors of other kinds. The left also seems to be associated with what is known as “executive function”—top-down control, overcoming impulses, thinking things through.
What does the right hemisphere specialize in? One example is the processing of melodies. That’s interesting; we might have expected that a melody is a bit like a sentence. An important case is touch, which features a strong role for the right hemisphere and left hand. Suppose you are asked to detect (perhaps compare) shapes that are put in your hand, doing so just by touch. Are two shapes the same or not? This is done better in the left hand. The right hemisphere seems generally better with spatial tasks. And in case you were thinking the right brain is a bit primitive, it is better with numbers. That makes some sense of the question about melody that I mused about a moment ago—music is not like language, to the brain, but like mathematics.
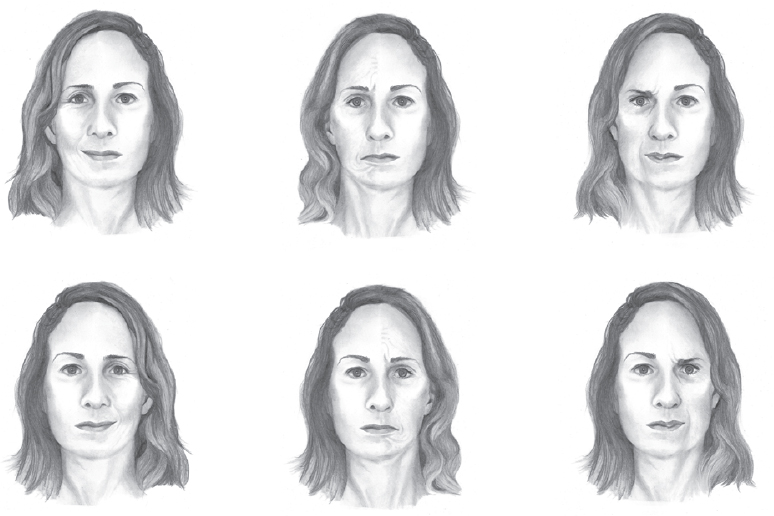
Face recognition is more a right-hemisphere skill, and the recognition of emotions is, too. Some simple but striking work has been done in which people are shown manipulated photographs or drawings of a face where one side has an emotional expression and the other side is neutral. How much emotion is seen? Again, the right hemisphere processes visual stimuli from the left side of the visual field, and if the left side of a face, as seen by the viewer, is the emotional one (happy, surprised, angry, whatever), then the face tends to look more emotional than if it’s the other side. This page has an illustration of the effect, in which the top row of faces all show emotion on the left but not the right (in sequence: happiness, sadness, anger), while the faces in the bottom row have the two sides reversed. In experiments, the faces are usually shown in pairs rather than all at once, with an emotion on just the left or the right—this is an illustration, not an image used in a study. Positive emotions seem to have a stronger bias than negative emotions, at least for some viewers. In my own case, the effect is pretty strong, especially for happiness, even when the faces are shown together like this.
A setup with these differences between the two hemispheres does make sense, as advantages can often be gained from handling different tasks with specialized mechanisms that work in a partly self-contained manner. That is fine as long as you can bring everything together when you need to, and that can be done by the corpus callosum, the highway between the hemispheres. This highway is found only in mammals, and not all mammals (not marsupials, or egg-laying monotremes like the platypus). The presence of this highway seems to allow specialization of the two sides to become more elaborate, as Michael Gazzaniga, one of the main split-brain researchers, has argued.
If the corpus callosum has been cut, then we see left-right differences in starker form, and one thing we see is a distinctive style of cognition on the left. The left, verbal hemisphere is then the one most readily interacted with, and as Gazzaniga observes, it makes up stories about things it does not and cannot know. The left looks for narratives, for patterns and unity, and tries to get things to make sense even when they don’t. In experiments, the left hand (controlled by the right hemisphere) sometimes acts on information that the left side of the brain does not have access to, and when the person is asked to comment on a behavior of that kind, they will make up an explanation. In one experiment, a patient had the word “bell” shown to his right hemisphere and the word “music” to his left. When he pointed with his left hand at a picture of a bell, the patient then explained this by saying that the last time he’d heard music it was from some bells nearby. (Gazzaniga adds that there were in fact bells nearby, but still …)
As well as engaging in this made-up storytelling, the left hemisphere tends to make other interesting mistakes. In some further experiments, split-brain patients were shown pictures representing everyday events and hours later were asked to say whether any of a second collection of pictures had also appeared in the earlier one. Both hemispheres were equally accurate in recognizing previously viewed pictures and rejecting others that were not shown earlier—as long as the new pictures, the ones not previously shown, were not thematically related to any of the earlier ones. The left hemisphere would incorrectly “remember” seeing pictures that had not been shown before if the pictures were thematically related to an earlier one. For example, the left hemisphere might see a picture of a man looking at his alarm clock in the first stage of the experiment and later see a new picture of the man brushing his teeth, and he’d be pretty likely to misremember having seen the teeth-brushing picture earlier, whereas the right hemisphere would not.
Gazzaniga’s view is that our left brains contain an “interpreter,” a side of the mind, or almost a sub-mind, that has a particular style: inquisitive, a seeker of patterns, a lover of narrative. The interpreter is so invested in unity and narrative that it makes things up when it has to. At least in a case where the person is intact, treating this as a “sub-mind” (this is my term, not his) may go too far, but the idea that the left hemisphere has a particular cognitive style, and sends us down this unity-seeking road when it can, seems a powerful one.
People have come up with a number of evolutionary sequences (narratives!) that could unify (!) and make sense (!) of all of this. Here, I make use of several of these, adding some ideas of my own.
The bilaterally symmetrical body form appeared early in animal evolution. This body form, which enabled new kinds of action, also brought with it the duplication of many parts, including parts of the brain. Along with vertebrates, many other animals seem to show some specialization or a difference in “style” across the two sides of their brain. Not all the same things end up on the same side in all animals (especially as vertebrates are the only ones with those crossed wires, with the left hand controlled by the right side of the brain, and so on). In vertebrates, the right brain seems more attuned to relations, especially social relations and relations in space, while the left side looks for categorizations. An intriguing difference that bears on conscious experience is mentioned by the lateralization expert Lesley Rogers. She writes that the left side tends to operate with “focused” attention on particular things, while the right’s attentional span is broader, more open. This left/right difference is seen in nonhuman species as well as us.
Then, in our own ancestors, language arises. It gets housed in the left (in a large majority, even most left-handers). Once installed, language has cascading effects, not only on social behavior but also on thinking. It becomes a means by which that left-side style—the storytelling, theory-seeking style—becomes strongly established.
The right side is not antisocial; as we saw, the right is better at recognizing faces and tracking emotions. The left/right difference does not map to a difference between a more solitary mind and a socially embedded one. But in those narrative-seeking, storytelling, left-side tendencies, we see something about what language and culture have done to our thinking. This is not a situation where the left and right process things the same way, but only the left can tell stories about it; as we saw in those memory experiments, the two sides don’t process things the same way.
In the memory experiments I mentioned a moment ago, we saw different tendencies across left and right. The left tends to misremember events when this serves a narrative, and the right does not. It’s natural then to ask: Which side is closer to how normal humans (those with an intact corpus callosum) behave? This is an easier question to investigate, and in some later work, Gazzaniga found that a normal, intact person shows those patterns peculiar to the left hemisphere. It is easy to induce people to make errors of falsely remembering that they’ve already seen items that are “thematically related” to ones they have seen earlier. At least in this kind of task, the unity-seeking style of thought prevails.
There’s a side of our minds that can sometimes get a bit lost in subtle psychological experiments on sensing, remembering, and so on. What I have in mind is conscious thought, especially where one pauses and reflects. This is particularly important in human projects with large effects: making plans, embarking on new ventures, and so on. Those decisions are often social—deliberation and choice are tied up with defense and discussion—but they also have a psychological side, and a role for conscious reflection.
In this sort of thinking, language is a tool with a unique combination of properties. By means of language we can engage in mental quasi-acts, internal sayings and rehearsings. These can help us assess, remember, and affirm ideas. With the aid of language you can also connect just about any idea with any other. The power for free recombination of ideas that gives language much of its social importance gives it power in our minds as well. The internal version of language can go further, in some ways, than an external version. You can run along a verbalized soliloquy and introduce a remembered image or smell as if it was another word. Inner speech can orchestrate an internal multimedia event.
Some of what I’m talking about here falls into the category of reasoning. In their book The Enigma of Reason, the French cognitive scientists Hugo Mercier and Dan Sperber argue that although philosophers and others have tended to see reasoning as a paradigm of self-contained, individual thought, its evolutionary origins are probably more social. These origins lie in projects of coordinating social life and, especially, trying to convince others of what to think and do. For Mercier and Sperber, we don’t reason to find conclusions, but to get others to agree with our conclusions. They allow that our brains have many processes of “inference” going on inside them that we use to work out what to do and think. These processes are usually unconscious. Reasoning, as they see it, is something more specific. This is the process where one idea is explicitly brought in as support for another: you should think this because of that. Language is then pretty much essential, as it enables you to bring ideas together in the right form, putting one idea in place as the justification for another.
Mercier and Sperber argue that the essentially social role of reasoning shows up in our strengths and weaknesses, including the logical mistakes we tend to make. These “errors” don’t show that reasoning is a bad tool for its purpose; instead, we’ve mistaken where reason came from and what that purpose is. An example of an error we all tend to make is confirmation bias—looking for things that support our present view, and not seeing evidence that pushes the other way. In a project of trying to convince others to follow us, this is no error at all.
Mercier and Sperber’s idea about origins seems pretty reasonable (!) to me. A difference between their picture and mine is the extent to which these origins hang over the activity today. Even if reasoning has its origin in trying to persuade, this skill can operate in a new way inside an individual. There, it can become a means to find new conclusions, to probe ahead and uncover what is hidden.
A category that tends to recede in their story is deliberation—by an individual, rather than a group such as a jury. For Mercier and Sperber, there are inferences, which are mostly unconscious, and there’s reasoning in the sense of reason-giving—“here is why this is the right conclusion …” Deliberation is not the same: “Something has gone wrong; what should I do next?” This activity is akin to reasoning in the role that language has, at least much of the time, but it’s more open, more exploratory. And this feeling of “openness” need not be a mere rationalizing illusion. Old tools can be given new roles. We can step back and reflect, using this socially derived tool. We can ask, “What sort of food should I really be eating, when I factor in everything I can think of?”
A related way in which language and culture affect our minds involves the nature of the self. This theme takes us back to the ideas about narrative that were emphasized a few pages back. Part of what gives us a sense of having a definite self is the feeling that we are a subject in a coherent story that extends over time. Some aspects of selfhood do not depend on any of this. There is a selfhood that comes from the physical unity that animals of our kind have as a result of evolution, and also from the memories we have of past events. But as well as the memories themselves, there’s the role of our narrative urge. We tie old memories into a story and try to make sense of continuity through change. This probably depends on language, our narrative tool. When we do this, some of what we arrive at is “cooked up,” to various degrees—it introduces fictions, and smooths over rough patches. But the inner narrator is not just a commentator who comes along after the fact; it can also shape how one handles the next choices and challenges that arise. This is who I am. As you interpret your past, you affect your present and what you might do and become in the future.
Let’s follow these paths a little further. When I’ve talked about “language” so far in this section, I’ve meant spoken language. Literacy has its own role, though. As we saw in the previous chapter, reading and writing are relatively recent cultural inventions, and most humans have not been literate. But when literacy comes on board, it has effects on the brain. As I said in the previous chapter, it increases the size of the corpus callosum and changes patterns of activity seen in brain scans, with a left-brain area becoming very active when recognizing words. This appears to reorganize our face recognition skill, pushing it more to the right than before. I said some pages back that emotion recognition is stronger in the right hemisphere, and that was written as a claim about people—everyone. But we should wonder: Were all the people who were tested on emotion recognition able to read? Might their brains have been rewired by reading in a way that makes the result less applicable, or not applicable at all, to non-readers? We might also give some thought to the language they were reading. Over thirty years ago, Jyotsna Vaid and Maharaj Singh wondered whether the usual results showing left visual field processing of facial emotions might be due in part to the left-to-right reading habits of the subjects; this might make a left-to-right scan “come naturally.” They compared how those artificial composite faces I mentioned earlier were interpreted by readers of several different languages—Hindi, Arabic, Urdu—including some people who read both left-to-right (Hindi) and right-to-left (Urdu) scripts. It was quite hard to interpret the results they found, but there did seem to be an effect of reading background, where the left visual field bias for emotion processing was more marked in people who read left-to-right. On the other hand, a leftward bias in looking at faces has been seen in some other animals—rhesus monkeys and domestic dogs—though the dogs had it when looking at us, not at other dogs. Human-like responses to chimeric faces (with emotion on the left seen as stronger) have also been found in chimps. The situation seems to be a real tangle, with a role for features of our brains that are shared across other animals, and a role for the details of culture as well.
Giorgio Vallortigara, writing about animals with deeper divides between the left and right sides, such as fish, reptiles, and birds, wondered where the felt experience of these animals ends up. The question is especially acute when an animal’s eyes are on the sides of its head, as in fish, but not only then. Might such an animal have two different sensory perspectives on the world, each with a different feel? Or might the animal retain a single point of view, but find the two sides of the world have a different “coloring,” with the experienced-left side more socially loaded and more relationally connected?
That second option would be an extreme case of something that seems to be present also in us, with our right hemispheres picking up emotions better than the left. Those artificial faces with emotion on one side and not the other are very striking. I have become so curious about this that I keep trying to attend to the left of my visual field, and that doesn’t work, as it just brings the “left” into the center. (Closing an eye doesn’t work, as each of our eyes sends information to both left and right hemispheres.) This point about movement is an important one, too, because one way animals deal with the quirks of their left-right separation is by continually moving their head—you see a lot of this in birds. A unity of subjectivity is achieved in part through action.
Do filmmakers make use of any of this? In film, motion from left to right is often seen as having a different feel from right to left. Left to right movement is seen as progressive and natural. Exactly the same sequence reversed by flipping each frame, so the motion starts on the right, can look disconcerting. This has gone from being semi-implicit lore, and perhaps something that early filmmakers made use of without realizing it, to being a topic of scientific work. A famous essay by the critic Roger Ebert, written before the scientific work started, listed a left-to-right preference for movement among a number of other generalizations, including the idea that “right is more positive, left more negative.” People have tended to explain left-right phenomena in movies in terms of explicit cultural ideas in the West about left and right (an old word for left being “sinister”). But all this might also have to do with perceptual effects that stem from lateralization of the brain.
If that is so, the causal relationships here might form quite a thicket. I said a page or so back that, when looking again at how we process faces for signs of emotion, language differences seem to have a role—people who read a right-to-left language are different from those who read a left-to-right one. A similar effect has been found with action sequences in video clips. In a study of how soccer goals were interpreted (as stronger, faster, more beautiful), native speakers of Italian and Arabic differed quite a lot in how the direction of the action affected their judgment. Arabic, unlike Italian, is a right-to-left language on the page, and the Arabic speakers interpreted right-to-left goals as stronger, faster, and more beautiful than left-to-right ones. Some of those ideas about filmmaking I just mentioned could be more specific to particular cultures than they initially seemed.
Here is one more thought about the psychological role of language. Our minds have a side that is intensely private—mind-wandering, reminiscence, disconnected free reverie. Until the watchers in a dystopian future start scanning us all the time, no one need know what you are imagining or remembering. Some of this is present in nonhuman animals; mental exploration of space is visible in the rat’s brain, as a result of the inner maps mentioned earlier. A rat can be quite physically still, asleep or awake, while activating a series of “place cells” in its brain that normally register its actual location. Whether this mental exploration is felt or not by the rat, it is a kind of offline rehearsal of action. This is related to dreaming, for which the evidence is now strong in quite a number of nonhuman animals.
This side of the mind, full of quirks and the unrepeatable, is the home of a pure kind of individuality. But the social tool of language, once internalized, gives special power and reach to what we do here. Again, this side of the mind is not wholly dependent on language, as seen in the mentally meandering rats. But with language on board, it flowers. It makes possible a form of private creative thought—or private at first, made public if we choose—that is immensely rich. Reverie can be steered and magnified by letting the narrative side of the mind run on, calling on traces of images and sounds as it does so. You can, as the philosopher John Dewey said a century ago, tell tales to yourself that no one else would believe, and do so with good reason, to your benefit. Language expands the power of free combination, of contemplation of the merely possible and impossible. An enlargement of the private is made possible by the internalization of a social tool. This distinctively human combination of the private (offline, whimsical) side of the mind and its basis in a social and cultural set of skills seems incongruous, almost ironic. It is a great gift from the public to the private.
Hyperscan
My sketch of the biological basis of felt experience at the start of this chapter had two elements. The first involves the role of sensing and acting in the animal way of being—the origin of subjectivity—and the other makes much of some physical peculiarities of brains and nervous systems, with their mixture of cell-to-cell influences and large-scale dynamic patterns. The ideas we were looking at just now, concerning language, the different hemispheres, and the role of culture, relate pretty clearly to the first of those two—to sensing, point of view, and so on. If we turn to the other part of my sketch, it might seem unlikely that much will be added to this story about consciousness and human life. The electrical rhythms that I discussed earlier are found across many animals, including species with brains very unlike ours. If we’re interested in consciousness and culture, it initially seems that those features shouldn’t make a lot of difference … Not so fast.
The last few decades have seen advances in brain-scanning techniques. I sketched some basics earlier in this chapter. The EEG method dates from around 1929, and a lot has happened since then. A 2002 paper set a lot of work in motion; it was called “Hyperscanning: Simultaneous fMRI During Linked Social Interactions,” by a team led by Read Montague. The paper explores the idea of scanning the brains of two or more people at a time, when they are engaged in social interactions.
They used the fMRI method (functional magnetic resonance imaging). This is a scan of the brain, from outside, that is not looking directly at neurons and their electrical activity, but at changes in the oxygen supply, through the blood, to different parts of the brain. The social interaction used in the study was a simple game of deception, where one person sends a message and the other has to guess whether they lied. The two people were not face-to-face, but in separate scanning machines. The main idea of the paper was to introduce the method, and they don’t go through detailed results, but it showed some correlations between brain activity on each side. The paper introduced the term “hyperscanning,” and the method took off.
That is the conventional opening of the story. In fact, there was an earlier experiment, published in 1965. This one used not fMRI (which had not been invented) but EEG, a more direct pickup of overall electrical activity in the brain. The paper, by two ophthalmologists, T. D. Duane and Thomas Behrendt, came out in Science magazine on a single page. It was an ESP study of “identical” (single-zygote) twins.
This is an appealing replay of history. The EEG method itself was introduced by Hans Berger as part of a search for telepathy. Decades later, the first known study of real-time relationships between two people’s EEG rhythms was another telepathy study. The ophthalmologists were interested in communication across unknown channels. In the test, one twin would close their eyes in a lit room. Closing the eyes tends to initiate a particular kind of rhythm (alpha rhythms) in the brain. Might the other twin, in a separate room, enter the same brain wave pattern also? They reported that in some cases, the answer was yes.
This study was done very informally, with no statistics. A few other studies of this kind followed, including a much more technically sophisticated one, not using twins, led by a Mexican researcher whose career bridged scientific and shamanistic approaches, Jacobo Grinberg-Zylberbaum. This 1992 experiment, using EEG scans, reported success in transmission across nearby but shielded rooms. Grinberg-Zylberbaum disappeared, in a still-unsolved mystery, two years later.
I am not as instantly dismissive of this sort of work as many people are. I’m open to the possibility of unexpected channels, but skeptical on balance. Anyway, as with EEG’s discovery by Berger, the double-scanning method became mainstream, in this case after the 2002 paper that introduced the term “hyperscanning.” A lot of papers about hyperscanning that include a quick history at the start do not mention the adventurous ophthalmologists’ paper at all.
The usual goal of hyperscanning work is to look at the relationship between the brain activity in two different people during a social interaction, and make comparisons to other situations. Several different kinds of scanning are used: fMRI, EEG, and a couple of others. The EEG work is especially relevant for the themes of this chapter, as it looks directly at large-scale electrical patterns in the brain while allowing people in the experiment to interact naturally. The other methods contribute to our picture in different ways.
The picture emerging is a surprising one. In some social settings, especially those featuring cooperation and teamwork, there’s a lot of correlation between the large-scale dynamic patterns in one brain and the patterns in another. This is called “interbrain synchrony.” In measuring the correlations we reach some technical topics that I understand very imperfectly. You are looking at a pair of rhythms, or a series of events over two brains, and seeing if they match up to some extent. This might be done by looking at their frequencies, or at the “power” of the oscillations (are both rhythms strong when one is strong?). You might look at the phase (are the peaks lined up to some degree?), or at combinations of these. You also have to try to work out whether any synchronization is just due to some external cause affecting both people, rather than mutual influence between them. Many studies find that the patterns do show associations of some kind when the people are socially engaged, to a degree that is often surprising, given the physical separateness of the two brains. The brain rhythms don’t fall exactly into sync, and all this is a matter of degree. But it’s still, surely, a surprise.
The synchronization doesn’t just happen when people are looking at the same thing or performing exactly the same actions. Two people playing a guitar duet do synchronize, but so do two people who have to cooperate to perform a task and have different roles—for example, one person might have to locate a target in a scene and use their eye gaze to send information to another person who hits a button, or two pilots might be working together in a flight simulator. When people are in a situation where they can choose to be more cooperative or less so (in a computer game), synchronization is greater when they are being cooperative, and a researcher can even predict cooperative interactions from whether people’s brains are in sync. In a more purely cooperative task that two people had to do together, pairs with higher levels of synchronization performed better than pairs with less. Synchronization of brain rhythms also seems to be part of the explanation for the “cocktail party effect,” where a listener can pick one voice out from several that are going at once, and follow what is being said by that person despite noise and competition.
This whole phenomenon is especially puzzling when the brain rhythms that are matched are very fast. In work using the EEG scan, synchrony of brain rhythms can occur in the “gamma” band. This is a speed of oscillation above twenty-five cycles per second, sometimes much faster. That seems too fast for the synchronization to be controlled by anything visible in a person’s behavior.
I’ll come back to those questions about how the contact is achieved, but let’s first ask something else. What might all this do, when it is real? What difference might be made by the syncing, whether it’s those surprising cases with very fast cycles or something else? Reports made in these studies sometimes include a feeling of contact between people, a sense of being—and now the old metaphors take on a new meaning—attuned, in sync, with someone. This is seen as one of the things that synchronization might explain. How could this be possible, though? How could you tell that someone’s invisible super-fast rhythms are in sync with yours, or are not? What difference could it make?
The most radical option (outside of the telepathic camp) that I know of is a view outlined by the cognitive scientists Ana Lucía Valencia and Tom Froese. They suggest that there is a partial merging of the physical basis for the experience in one person and the experience in another. Part of the physical basis for “your” experience is activity in the other person’s brain, and part of the basis of their experience is in your brain. The sense of sharing and contact with another that we sometimes find and often value might have a more substantial physical reality than we had supposed. As Froese put it in another paper, “when we become aware that ‘we’ are sharing a moment with someone else, it is no longer necessarily the case that we are fundamentally separated by our distinct heads—we could really be two distinct individuals sharing in one and the same unfolding experience.”
This goes against the usual view, one that otherwise seems entirely adequate, that each of us is an individual subject, perhaps having similar experiences to someone else, but each having our own, with no literal merging between them. If that is wrong, it is truly a massive change.
I want to be open-minded about these possibilities. I accept a materialist view of the mind. This means that there are some physical processes in nature that are, themselves, also mental processes, and this includes sensations. It’s not that these physical processes cause the mental ones; they are them. Every sensation has a physical basis of some kind, a basis that exists in some particular place. In addition, other physical goings-on might cause the sensation while not being part of it. The thing that I am calling (and that Valencia and Froese also call) the “basis” of the sensation or experience might always be entirely inside one person’s head, or not always so. Are there reasons to expect things to be one way or the other?
If minds and brains are for controlling actions, and those actions animate individual animal bodies, this gives us some initial reason to think that the physical basis for experience will always be localized inside an animal. Valencia and Froese also discuss an argument that was given by the cognitive scientist Andy Clark, where he says that conscious experience will require very fast and intricate activity in whatever physical basis it has, and this probably can’t take place across two different brains because the interface between them, their separate bodies, would be too slow. The bodies of the two people would act as a kind of filter (a “low pass filter”) between them, so that all the rapid and high-frequency activity that consciousness relies on would have to take place separately in one person and the other, not in a unit that is spread across them. Valencia and Froese respond that although this argument about the body as a filter initially looks completely reasonable, the evidence from hyperscanning suggests it might not be right. Somehow, two brains can become coupled together in surprising ways, and that opens up the question of whether the physical basis of an experience, and hence the experience itself, might be literally shared across them.
An alternative view is that the hyperscanning experiments are picking up not a merging, but a connection between two minds that stay distinct from each other. If this is offered as an explanation of the feeling of togetherness or contact across people, it is less directly an explanation than the more radical view. In the radical view, a feeling of contact comes from a partial unity between minds, a partial blending. In the alternative view, it’s a meeting without being a merging; the feelings on each side are separate, but are affected in subtle ways by what is happening on the other, and that shows up as an experience of contact. What would those effects be, and how might they arise?
The main possibility to look at seems to be that subtle cues in the behaviors on each side are picked up and lead to a degree of synchronization in some internal processes. This feeling of alignment might have effects on the two people’s experience, as is more overtly seen in the case of dancing.
A problem to grapple with is how this could work in the case of the faster neural rhythms. It’s not hard to see how similar rhythms across two people could be switched on at roughly the same time. But it would be hard for this to be very exact, I assume, and it wouldn’t explain ongoing synchronization across the two people, in any case. What might bring about the tight connections? I will offer a few ideas (making use of other discussions).
In an old experiment done first by chance in the seventeenth century, the Dutch physicist and inventor Christiaan Huygens noted that two pendulum clocks near each other will become synchronized (though they end up swinging in opposing directions—out of phase) if their rhythms start out fairly similar and the clocks are also physically similar. Their motions come into alignment. This also works with metronomes. The effect is pretty puzzling, and it has now been studied quite a lot. The picture we have is that the effect does work by physical influences that go through whatever the clocks or metronomes are fixed onto (no metronome ESP), and the nature of the influence—I think!—is generally like this. Each device as it moves tends to nudge the other—gives it a little “kick,” as some writers say—that perturbs it from its current pattern, whatever that might be. The mutual nudges or kicks go on and on, but if the two clocks happen to fall into a state where they are closely synchronized, then the kicks cancel and neither affects the other. They are in an equilibrium.
In the physics cases, with clocks and metronomes, this is all pretty exact. In the brain, nothing will be so neat. But there might be a situation in which tiny visible movements and other cues in a social setting act to nudge and perturb two people’s brain activity, and when this happens, some arrangements across the two people are more stable than others—they don’t get pushed away from, or not as much. Then the two people’s brain patterns might be brought closer and closer together, even with respect to those very fast patterns of activity. Perhaps what people can track in each other is a complicated combination of movements spread across the whole body. The way a person produces many tiny actions together might have, without them realizing it, a subtle rhythmic signature. These rhythmic signatures in small ordinary motions might play a role in explaining the feelings of contact between people that I mentioned earlier.
All this is very much on the edge—it is scientifically controversial, attracts critical papers with titles like “Beyond the Hype,” and so on, and its importance is hard to interpret. But I am struck by this work. The hidden lining-up of activity seems to me remarkable, even without any argument for a literal merging of minds. This work has already changed my intuitive sense of how some everyday interactions go. An actual cocktail party now seems different; it is, among other things, a sea of interacting brain rhythms, going in and out of sync. Feelings of attunement or contact between people, whether stable or fleeting, might be taken more seriously as physical realities than they have been before. They are not just unconscious judgments (hmm, this person is okay), but reflect something deeper in the links between us.
I’ll mention one other idea relating to the social role of interbrain synchrony, linking to the previous chapter. This idea is usually discussed independently of any exotic neural ideas. The idea is that a distinctive part of human life is the formation of shared intentions, which are sometimes called “we-intentions.” In some cooperative situations, it’s not just that I want to get something done and I know that you might be inclined to help me; more than this, we get to a stage where the intention is something that exists across both of us, a we-intention. We intend to move this big log over there. I’ve not seen this idea as pivotal in the way some others do. In part, that is because I’ve thought that we can understand collaboration quite well in a more individualist way. I have beliefs (or at least assumptions) about what you are up to, and you have beliefs about what I am up to, and so on. Valencia and Froese, as you might expect, think there might be more reality to these special mental states, we-intentions, and think that interbrain coupling might be part of how they are achieved. They even wonder whether the we-perspective might be primary for us in some sense, fundamental in human life, so that I-thinking comes later. I doubt that. The acting I is more basic than the acting we, potent though the latter may be.
Brothers
That is human consciousness as I see it, a combination of features of felt experience that come from the animal way of being and a powerful cultural overlay. That is what happened to a group of primates, initially just another skinny branch of the evolutionary tree, over a few million years.
Alongside us all through this book and the two that preceded it, Other Minds and Metazoa, has been the genealogical tree of animals, the animal part of the tree of life. Our branch is the mammals. In Africa and Australia, one gets a strong sense of this part of the tree. In Australia, we see traces of the lower branchings heading off to a few distant cousins—echidnas and platypuses, and then marsupials. In Africa, we encounter the great thicket of branchings within eutherian mammals, the group on the other side of the split from marsupials.
That evolutionary thicket has proved hard to fully resolve, but it includes one early split leading to elephants and a few others, including hyraxes (like outsized, scrambling guinea pigs) and aardvarks. In the branch on the other side of that split, we find a divide that shapes two big collections of familiar mammals. On one side are horses, deer, dogs; on the other, primates, with a collection of rodents and rabbits. (The previous page has one more “tree of life” drawing, simplified as usual, that illustrates some of this.) If we look across from our home with the rodents, we see giraffes walking, as Karen Blixen said, like long-stemmed flowers, along with antelope, jackals, and whales.

Dolphins and other whales are in that group. Manatees, on the other hand, are with the elephants. A return to the water happened several times in different groups of mammals.
In that big cluster of mammals along with the deer and the whales are also cats. In the Maasai Mara, Kenya, I saw cheetahs in the wild. Out on grasslands in the sun, two of similar size were jogging along together. They were radiant, almost glowing, it seemed. I thought, right away and unexpectedly, that these were the most beautiful animals I had ever seen.
Afterward, I asked one of our guides, Collins, about them. One was male, I saw. The other? He said that both were male, young brothers. They had formed a coalition, a bond, and would stay within it permanently.
One turned his head back, glancing at the other, to check without slowing. They seemed a miracle of animal evolution, but more fundamentally, a miracle of self-organization, a miracle on the part of matter itself, that it had come together into that form.
The combination of wild beauty and companionship was overwhelming. I expect never to forget them. Those radiant animals, with their bond, jogging together through my dreams.