If your vision is normal, imagining a world without the ability to perceive depth and colour is difficult. But for people who can’t see these things, the problem can be highly debilitating.
How the human brain processes these aspects of the visual experience is quite remarkable. (To read about the basic biology of human perception and some theories about how people see the world around them, turn to Chapter 4.) As well as describing here how you see depth and perceive colours, we also include real-world examples and some fascinating illusions that fool your visual system. The world isn’t what it appears to be!
Seeing the Third Dimension
Depth perception is vital – without it you wouldn’t be able to cross the road, pick up things or even identify what things are. For cognitive psychologists, establishing how this awesome skill works is really important. For one reason, it highlights how the human mind influences how people see. For another, people need depth cues to know about things (and knowing is very cognitive). In this section, we introduce depth perception and review the various cues to depth.
Introducing depth perception
Being able to see in three dimensions (3-D) is critical. Without depth perception, you’d walk into lampposts, knock things over and not realise how far away a speeding car is. Just think of this example: you’re sitting at a bar and you need to pick up your pint. Without depth perception, doing so would be hit or miss and you may end up spilling it – a tragedy!
People use depth perception to work out how far away things are from them – called egocentric distance. For example, you know that your laptop is a couple of feet in front of you when you’re on Facebook, er, we mean writing a research paper. You also use depth perception to work out how far things are away from each other – called object-relative distance.
You can also use depth perception to help identify objects. For example, look at Figure 5-1. You can see a gorgeous chinchilla behind the wires of her cage, because depth perception allows you to tell the difference between the animal’s grey fur and the metal of her cage.
Figure 5-1: Can you see where the chinchilla ends and her cage begins?
The eye of the tiger: Monocular cues
A variety of cues signal depth. A common fallacy is that you need two eyes for depth perception, but as you can see from Figure 5-2, only two cues are based on binocular (two-eyed) vision. In fact, you can determine many depth cues using only one eye. These monocular cues are also known as pictorial depth cues, because you can obtain them from a simple 2-D image. When you look at a picture, having one eye is just as good as two eyes.
The simplest cue to depth is size. Generally, the bigger something is, the closer it is to you. Simple … or maybe not. Consider looking at a cube in isolation. You can’t tell how big it is without some other reference point. The cube can be any size, so you need something to compare it to.
You need a reference point only for ambiguous or unfamiliar objects. For example, if a rabbit appears 2 centimetres tall, you know that it’s quite far away, because rabbits are larger than that. Therefore, size cues require familiarity with the object or a reference point.
Linear perspective is related to size cues. Think about walking down a straight road at night. As you look down the road, it appears to narrow, and the lampposts get closer together. Although you see this, you perceive that the road ahead is farther away from you. (Chapter 4 covers why seeing and perceiving aren’t always the same thing in more detail.)
Figure 5-3 shows a variant of the Ponzo illusion (named after its creator, the Italian psychologist Mario Ponzo). In the figure, you perceive the top horizontal bar as larger than the bottom one, even though they’re exactly the same size; linear perspective tells you that the two converging lines are in fact parallel. Therefore, the upper line must be bigger because it almost touches the perceived-to-be parallel lines whereas the bottom line doesn’t.
Figure 5-4 shows the Müller-Lyer illusion (the German sociologist Franz Carl Müller-Lyer came up with it) – this illusion is similar to the Ponzo illusion and is potentially a result of implied depth perception. The line with the outward pointing arrows seems shorter than the arrow with the inward pointing arrows despite the fact that the lines are the same length. One hypothesis for this effect is that people are so used to seeing inward pointing arrows in the corners of rooms and outward pointing arrows on the exterior corners of buildings. Evidence shows that Western peoples are more susceptible to this illusion than pre-Stone age forest-dwelling peoples.
In the real world, you can experience size depth illusions simply by looking at the moon (appropriately enough called the moon illusion). When it’s high in the sky, it appears smaller than when it’s close to the horizon.
Very similar to linear perspective are texture gradients. If you’re on a pebbly beach and you look along it, the pebbles appear larger near your feet but smaller farther away from your feet. Again, that’s the sensation – it’s what you see – but the perception is that all the pebbles are the same size: the ones that look smaller are simply farther away.
Another important cue is aerial perspective: objects appear blurrier and more faded in the distance. If you look out across the countryside, distant hills are slightly blurry. So, you know that the blurrier an object, the farther away it probably is.
Covering objects partially
Occlusion (also known as interposition) is another simple cue to depth. People naturally believe that objects will be complete.
Figure 5-5a appears to show an illusory bar positioned over three circles. You don’t perceive three incomplete circles though; you perceive a bar over the top. In fact, your brain responds in the same way to the illusory bar as to the real bar. These shapes are similar to the figures in Figure 5-5b, where a square appears to be placed over the top of the four circles.
Figure 5-5: Are the circles complete or incomplete? a) you perceive a bar; b) you perceive a square.
The Necker cube (see Figure 5-6a) highlights occlusion nicely. When presented with a hexagonal shape with extra lines, people tend to perceive a cube: that is, 3-D is so important that it pops out from a 2-D image. However, the basic Necker cube is ambiguous: is the far-left vertical the back or front of the cube? Only when some of the sides are occluded (see Figures 5-6b and 5-6c) can you clearly work out the depth.
Figure 5-6: The Necker cube: a) ambiguous depth; b) and c) obvious depths.
Transparency is a special case of occlusion, when one object covers another object but you can still see through it.
Occlusion is such a powerful cue to depth that it can override other cues. If you can manipulate an image so that the depth cues aren’t consistent, occlusion is the one cue that people appear to follow most readily. Occlusion may even be processed by a special place in the brain, because some people can’t see occlusion but can tell depth by other means.
The brain takes only 100 to 200 milliseconds (that’s one-tenth to one-fifth of a second) to process occlusion. In other words, when presented with one of those part circles in the earlier Figure 5-5, your brain registers the bar over a circle in 100 milliseconds. Prior to this, however, your brain thinks of the shape as a single object.
Shading the world
People know that light comes from above, so when they see a shadow they assume that it’s underneath something. Look at the pattern of dots and dimples in Figure 5-7. Those with light patches on top appear to be dots, whereas those with light sides on the bottom appear to be dimples.
Turn this book upside-down and watch the dots and dimples swap over. Now turn the book on its side – the dots and dimples may spontaneously swap and swap back. If you must, stand on your head to be sure. Whatever way you look at it, you know that light comes from above, which gives you an extra depth cue.
Flexing the eye muscles: Physiological cues
Six muscles control how each of your eyes move, and two muscles control the shape of the lens in the eye. These muscles change in tension depending on how far away an object is. Muscular movement of the eyes produces tiny electrical impulses that the brain can record and use to establish depth.
Accommodation happens when the muscles that control the lens by stretching the ligaments attached to the lens (which focuses incoming light) bend it sufficiently to deflect the light onto the back of the retina. When these muscles are relaxed, the lens is flat. Objects are about 3 metres away at this point. To bring a closer object into focus, the muscles have to relax, stretching the ligaments and allowing the lens to form its natural curved shape. These muscles can bend the lens sufficiently to bring an object 20 centimetres away into focus, but no nearer. So, accommodation is useful only for objects located between 20 centimetres and 3 metres from you.
Hold your finger directly in front of your face and slowly bring it closer to your nose: try to keep looking at it the whole time. To keep the image in your fovea (the middle part of each eye with the best resolution), you have to rotate your eyes (called vergence movement). You can probably feel your eyes begin to cross when your finger is about 10 centimetres away. Most people feel their eye muscles tense up. (Don’t hold it for too long – otherwise, it hurts!)
The brain can use this tension as a measure of depth perception. Vergence movement can provide accurate depth perception up to about 6 metres:
Convergence: Occurs when you have to roll your eyes inwards to see near objects.
Divergence: Occurs when you have to roll your eyes outwards to see distant objects.
Using both eyes together: Binocular cues
Here we consider the depth cues involving two eyes. Your two eyes are in slightly different positions and so what they see is slightly different.
You can demonstrate that you have different images on each retina (like double vision) simply by holding a finger out in front of you and noting what object your finger is in front of. Close one eye and your finger appears to shift positions; close the other eye and the finger appears to shift again. This effect is called retinal disparity. People move their eyes inwards and outwards so that corresponding points in the scene fall onto the two foveas:
Horopter: Region where no retinal disparity is present and you experience the singleness of vision. Points farther away or closer than the horopter fall onto non-corresponding parts of the two retinas, generating disparity. This disparity becomes greater, the farther in front or behind the horopter an object is.
Crossed disparity: Images closer to you than the horopter.
Uncrossed disparity: Images farther away than the horopter.
Normally your brain manages to form the two disparity images into a single image (called fusion). But if you suffer diplopia or are very drunk (we wouldn’t know personally, of course), you have double vision.
The human brain can calculate the amount of disparity between the images of each retina. This ability is called stereopsis, and it gives people a measure of depth – the more crossed disparity, the closer the object is; the more uncrossed disparity, the farther away the object is.
Humans have cells in the visual cortex that respond when some form of retinal disparity exists. About 5–10 per cent of the population (including one of your authors) lack this ability. These people are stereoblind (unable to detect depth based on retinal disparity) and have to make use of all the other depth cues for accurate depth perception. This condition is often the result of a strabismus, a squint or lazy-eye, at an early age that prevents the development of these disparity cells.
You can induce depth perception experimentally with random-dot stereograms – created by neuroscientist and psychologist Béla Julesz in 1971. Random-dot stereograms are patterns of black and white dots, in which one dot is presented to the left eye and another dot is presented to the right eye. These patterns are identical, except that a portion of one of the dots is shifted in one direction. This set-up creates disparity and is perceived as being nearer or farther away than the rest of the stereogram.
When you have no other cues available, you need to use two eyes for depth perception. To demonstrate, stand in front of a plain wall and close one eye. Hold your hands out with your forefingers pointing towards each other. Now try to make them touch. Most people find this quite difficult (not your stereoblind author!). Doing it with both eyes open is far easier.
On the move: Motion cues
Motion parallax is another form of depth perception based on a sort of retinal disparity (refer to the preceding section). Motion parallax is based on the fact that you have disparity due to movement.
Hold out your hand and raise two fingers – one behind the other. Staring straight on, you can’t see the finger that’s behind. However, if you move your head to the side, the back finger comes into view. The finger nearer to you should appear to move quicker than the finger that’s farther away, because of retinal disparity.
Similarly, if you’re on a train looking out the window and you keep your eyes fixed at a certain distance, objects near to you seem to move quickly in the direction opposite to your movement, whereas objects just beyond the fixation point appear to move in the same direction as you, slowly.
The human brain can calculate the relative speeds of the movement and use this information to work out how far away something is. This form of depth perception is incredibly powerful and widely used by animals for hunting.
Combining the depth cues
Given all the different cues to depth, you may be wondering how people use them to form an accurate representation of how far away something is from them. Psychologists haven’t established precisely how this happens, but experts do know that the more cues to depth perception that exist, the more accurate the judgement of depth is going to be.
Evidence suggests that some depth cues are processed in specific parts of the brain. Some patients with a brain injury can’t work out depth based on a particular cue but can use others, such as occlusion and retinal disparity.
Another way to view how the depth cues interact is to see what happens when the depth cues give conflicting information: for example, a huge mouse, partially occluded by a tiny elephant. Generally speaking, in this situation estimates of depth perception are very poor. The brain decides which cue is likely to be more reliable (how it does this is up to the individual brain) and uses that one cue at the expense of the others.
Living Life in Colour
Accurate colour perception can be the difference between life and death. Consider trying to find fruit nestled in trees if you can’t see colour. Animals (including humans) also use colour to convey psychological states: certain frogs turn bright yellow or red to signal danger, humans give away more than they mean to by going red with anger (or embarrassment) and some monkeys change colour to indicate a readiness to mate. Without colour, poor Curious George would miss out terribly.
Colour perception has massive effects on other forms of perception too, such as affecting a human’s sense of taste. Tests show that people dislike yellow strawberry milkshake even if the only difference between it and pink strawberry milkshake is the colour (people couldn’t tell them apart with their eyes shut). Which is why food manufacturers add colouring to many foods – mushy peas would be yellow without the food colouring.
In this section, we describe colour and show that although your eyes are designed to process only three colours, you can see a whole myriad of them. We discuss two theories that explain how people see the range of colours in the colour spectrum: trichromacy theory and opponent processing. We also show how the perception of colour isn’t as simple as identifying what the brain detects: colour perception is affected by knowledge as well.
Defining colour
Of course, colour makes the world look prettier, but what is it? Well, colour is simply the brain’s response to light of different wavelengths. Light is a form of electromagnetic radiation, which includes gamma rays, X-rays, ultraviolet light, infrared light, microwaves and radio waves. Although some animals can perceive other wavelengths, the bit that humans see lies between ultraviolet and infrared – often called the visible part of the spectrum.
This visible part ranges from 400 to 740 nanometres. Light waves in the 400–500 nanometres range are purples; those in the 580–740 nanometres range are reds; and the rest of the rainbow colours lie in between.
Light waves don’t contain colour, just a wavelength and intensity (or luminance). Colour is a response from the human brain and physiology (refer to Chapter 4).
Counting the colours: Trichromacy theory
Try to name as many colours as you possibly can in a minute. Perhaps ask a friend to do it, too. You probably name the 11 basic colours that anthropologist Brent Berlin and linguist Paul Kay identified in 1961, including red, green, blue, yellow, grey, brown and pink. If you’re an artist, you may also list colours such as navy, indigo and violet. If you’re a computer scientist, you may mention cyan and magenta. If you work for a paint company, you can probably come up with an infinite number of other colour names (such as applewood green or warming sunshine yellow!).
Three is the magic number
You may be surprised to discover that people have only three types of colour receptors (cones) in their eyes.
Think about how many colours you can see in dimly lit conditions. In the dark, all colours tend to look pretty much the same. People don’t have colour vision in the dark, because the cones detect colour and they need more light to respond (refer to Chapter 4).
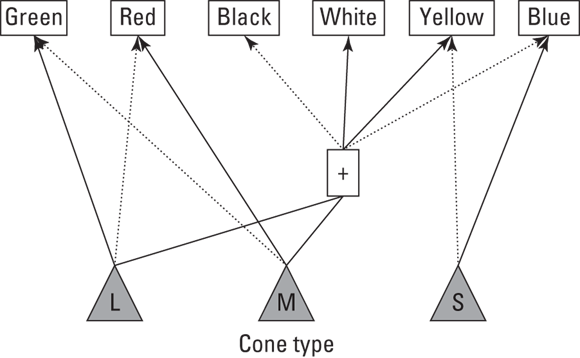
The three types of cones (S for short, M for medium and L for long) respond to different wavelengths of light. Microspectrophotometry (in which a small pinprick of light is shone on each photoreceptor and its electrical response is measured) shows to what wavelengths of light each cone type maximally responds. The peak sensitivity (the wavelength that causes the most amount of response) is as follows:
S-cones (420 nanometres): Approximately blue-purple.
M-cones (530 nanometres): Approximately yellow-green.
L-cones (560 nanometres): Red.
In other words, when a light of wavelength 500 nanometres is shone on the eye, all three types of cones respond. The response is greater, however, for the M-cones than the S-cones and L-cones. So, you interpret the colour to be green. From three cones, you can see all colours.
Added to the direct physiological evidence for this trichromacy theory of colour vision is a lot of behavioural evidence. People can make every colour from mixing these three colours. This theory is often referred to as the Young–Helmholtz theory, after Thomas Young and Hermann von Helmholtz, the key researchers who developed it.
It’s all in the genes
Recent evidence pinpoints the exact DNA responsible for these cones. The genes that control the cones are located on the X chromosome and other research has determined the cause for certain types of colour-blindness. People with colour-blindness (or, to be more accurate, colour deficiencies) usually have either a missing type of cone or an abnormal type of cone. Usually, they miss only one type of cone. Table 5-1 describes the types of colour deficiencies that experts have discovered.
Dichromacies (only two types of functioning colour receptors)
Protanopia
Missing L pigment
Confuses 520–700 nanometres (green to red)
Deuteranopia
Missing M pigment
Confuses 530–700 nanometres (yellow to red)
Tritanopia
Missing S pigment
Confuses 445–480 nanometres (blue)
Anomalous trichromacies (colour matches differ from normal)
Protanomaly
Abnormal L pigment
Abnormal matches; poor discrimination
Deuteranomaly
Missing M pigment
Confuses 530–700 nanometres (yellow to red)
The cone types aren’t evenly distributed in the retina, which means that people can detect certain colours more easily in the periphery than others. Specifically, no S-cones are in the fovea (the middle part of the eye) and so people are partially colour deficient in the fovea (because the fovea is so small, you never really notice). Also, many more M- and L-cones exist than S-cones. In fact, only 10 per cent of cones are S-cones; the rest are divided among the M-cones (30 per cent) and L-cones (60 per cent).
Colours in opposition: Adding more colours to the colour wheel
Although three cone types exist that respond to light of a particular wavelength (see the preceding section), people are clearly able to see many more colours. How is this possible? Well, experts aren’t sure. One suggestion is that certain colours are processed in some form of opposition. We look at some evidence for this idea here. If you remain unconvinced after reading this section, check out the later one ‘Categorical perception: Keeping colours straight’ for more on the theory.
In 1878, Ewald Hering, a German physiologist, observed the existence of four (rather than three) primary colours that people only ever perceive in opposition (the opponent-processing theory). He suggested that blue and yellow are mutually exclusive and red and green are mutually exclusive in a similar way that black and white are mutually exclusive – that is, they’re opposite sides of some sort of colour wheel or colour space.
Hering observed specifically that when describing colours, people never used the term ‘bluey-yellow’ but they did sometimes say ‘yellowy-green’ (the colour of a tennis ball). Likewise, they never said ‘reddy-green’.
Enjoying the after-effect
More compelling evidence for the opponent-processing theory comes from something called the colour after-effect. Before reading on, look at the demonstration in Figure 1 in the Part II online article (at www.dummies.com/extras/cognitivepsychology). Stare at the black dot for 30 seconds and then the image swaps to a plain white screen. Most people report seeing something instead of the plain white.
Usually, during the prolonged exposure to the colour, you become adapted to it (similar to the adaptation experiments we describe in Chapter 4). When you look at a blank screen, you see the opposite colours where the colours used to be: when you adapt to red, the after-image appears green; when you stare at yellow, the after-image appears blue; and when you stare at black, the after-image appears white. Thus, the colours are processed in opposition.
You can prove that this effect isn’t a computer trick, by adapting to the stimulus and then looking at a white wall: again you see the after-image. In fact, wherever you look, you see it. The after-image lasts for only a few seconds (though it returns when you blink), and it’s usually weaker than the original image seen.
The explanation for the colour after-effect is virtually the same as the explanation for all after-effects: processing a particular colour for a period of time fatigues the cells that respond to that particular colour, but has no effect on cells for other colours. Thus, when you look at a plain white image, the ‘opposite’ colour is relatively more active than the actual colour.
Joining opposites
With this knowledge in mind, how do these opposite colours combine to form the plethora of colours that people can see? Figure 5-8 offers the coding system based on the opponent-processing theory. Basically, the system works out the ratio of activation coming from the S-cones relative to the other cones. Solid lines represent that maximum response signals the colour; dotted lines represent that minimum response signals the colour.
When you know how the cones combine to form the colour signals that get transferred to the brain, assessing how the brain then copes with this colour information becomes important. Until fairly recently, researchers assumed that the cortical area known as V4 processed colour information – because some patients with achromatopsia (an inability to perceive any colour) had damage typically only to area V4.
Researchers discovered new patients, however, with damage only to this particular brain region and yet who could still perceive colour. This discovery led to an investigation into what other brain regions may be involved in the perception of colour.
Research focused on an area called V8 (which happens to be right next to V4). V8 seems to process colour, but no patients with damage exclusively to V8 have been reported yet. One study showed that when people are looking at colour during an adaptation experiment, both areas V4 and V8 are activated. However, during the after-image (in which only illusory colour is perceived), only area V8 is activated.
So these brain regions seem to process colour differently, but exactly how isn’t fully understood, yet …
Colour constancy: How colours remain the same
The light coming into your eye isn’t always the same as the colour you perceive, and yet it often seems so. Colours appear the same under all light conditions, even though the sensory input is different. This effect, called colour constancy, was described by Edwin Land in 1977.
Imagine that you’re getting ready to go out clubbing. You put on your favourite purple top and yellow trousers (yes, assume just for a moment that you have no dress sense!) and then look at yourself in the mirror under artificial light in your house. The colours seem clear. When you go out into the street and the orange glow of the street lights up your clothes, you still seem to be wearing purple and yellow. Then in the club, with a blue strobe light flashing, your clothes are the same: purple and yellow.
When you’re outside, the orange glow means that everything entering your eye is some shade of orange. But you seem to be wearing colour filters over your eyes that change every colour by the same amount. So, your brain somehow filters out the glow and determines that the colour of your clothes hasn’t changed.
How your brain does this is quite remarkable and not entirely understood. Here are three theories devised to explain colour constancy:
Adaptation: When in an environment that has one particular colour, people adapt to it, which removes it from their perception.
Anchoring (retinex): People find something that’s supposed to be white, and then label everything from there (like creating a baseline).
Computations: People identify one colour and then work through every colour boundary and compute what the colour must be from that boundary.
How does the brain represent colour constancy? Well, the higher visual areas are more responsive to context than the lower visual areas. So given the presence of cortical cells in the brain that respond to particular colours (for example, a blue cell that fires when S-cones are active), when all light is blue the S-cones are active even if the object would normally be yellow. The V1 area of the brain seems to respond as if the colour perceived were the colour sensed (no sign of colour constancy), but area V4 seems to respond as if the colour were the colour it would be under sunlight (following colour constancy principles). Hey, you didn’t expect a definitive answer, did you?
Categorical perception: Keeping colours divided
Categorical perception is when two items can’t be confused with each other – because people form a category, one thing is perceived as belonging to one category and can’t belong to another category.
In the earlier section ‘Colours in opposition: Adding more colours to the colour wheel’, we describe the opponent-process theory of colour perception, which suggests only three categorical boundaries: between red and green, between blue and yellow, and between black and white. Blue and yellow, say, are therefore mutually exclusive – a yellow can never be confused with a blue. But this isn’t what happens in the real world.
If you ask participants to name a series of colour stimuli, you tend to find more categories that aren’t readily confused. Participants see a series of yellows that gradually become more and more green. They normally describe the colours as yellow, and then suddenly at a particular amount of green (a particular wavelength), they say the colours are green.
Think of a tennis ball. What colour is it? Most people say yellow or green. (Obviously, those of you who say yellow are right.) Only a few people automatically say yellowy-green. That is, a sharp distinction exists between what’s considered yellow and what’s considered green.
If you use equiluminant stimuli (ones that have the same level of brightness) and the whole colour spectrum, English-speaking people tend to describe the colours as being one of the following: purple, blue, green, yellow, orange or red. These are the focal colours that have distinct categories.
Using these stimuli, researchers presented new English-speaking participants with two colours (one after the other). The colours need to be very similar, and differ only by a few nanometres in wavelength. If they cross a category boundary (where the name changes), participants can tell them apart: known as cross-category discrimination. If the colours are from the same category (so that people would always describe them as one colour), participants have difficulty telling them apart: known as within-category discrimination.
In 1987, Stevan Harnad found that the hallmark of categorical perception is that cross-category discriminations are relatively easy and within-category discriminations are hard. In experiments, participants find the discrimination between a colour patch of 550 nanometres (yellow) and one of 555 nanometres (yellow) harder than the one between 550 nanometres (yellow) and 545 (green), even though the physical difference is the same.
 People use depth perception to work out how far away things are from them – called egocentric distance. For example, you know that your laptop is a couple of feet in front of you when you’re on Facebook, er, we mean writing a research paper. You also use depth perception to work out how far things are away from each other – called object-relative distance.
People use depth perception to work out how far away things are from them – called egocentric distance. For example, you know that your laptop is a couple of feet in front of you when you’re on Facebook, er, we mean writing a research paper. You also use depth perception to work out how far things are away from each other – called object-relative distance.
 You need a reference point only for ambiguous or unfamiliar objects. For example, if a rabbit appears 2 centimetres tall, you know that it’s quite far away, because rabbits are larger than that. Therefore, size cues require familiarity with the object or a reference point.
You need a reference point only for ambiguous or unfamiliar objects. For example, if a rabbit appears 2 centimetres tall, you know that it’s quite far away, because rabbits are larger than that. Therefore, size cues require familiarity with the object or a reference point.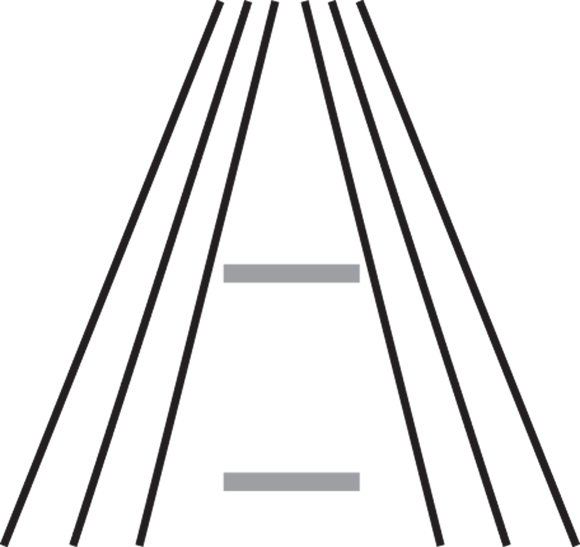

 In the real world, you can experience size depth illusions simply by looking at the moon (appropriately enough called the moon illusion). When it’s high in the sky, it appears smaller than when it’s close to the horizon.
In the real world, you can experience size depth illusions simply by looking at the moon (appropriately enough called the moon illusion). When it’s high in the sky, it appears smaller than when it’s close to the horizon.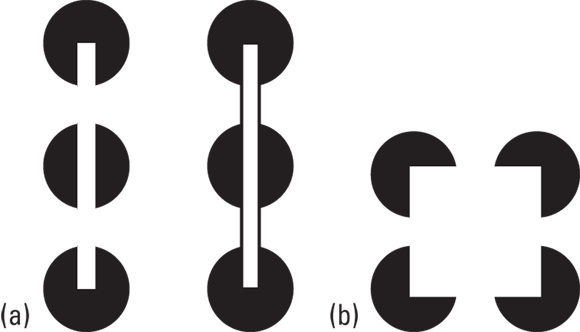

 Occlusion is such a powerful cue to depth that it can override other cues. If you can manipulate an image so that the depth cues aren’t consistent, occlusion is the one cue that people appear to follow most readily. Occlusion may even be processed by a special place in the brain, because some people can’t see occlusion but can tell depth by other means.
Occlusion is such a powerful cue to depth that it can override other cues. If you can manipulate an image so that the depth cues aren’t consistent, occlusion is the one cue that people appear to follow most readily. Occlusion may even be processed by a special place in the brain, because some people can’t see occlusion but can tell depth by other means.
 Turn this book upside-down and watch the dots and dimples swap over. Now turn the book on its side – the dots and dimples may spontaneously swap and swap back. If you must, stand on your head to be sure. Whatever way you look at it, you know that light comes from above, which gives you an extra depth cue.
Turn this book upside-down and watch the dots and dimples swap over. Now turn the book on its side – the dots and dimples may spontaneously swap and swap back. If you must, stand on your head to be sure. Whatever way you look at it, you know that light comes from above, which gives you an extra depth cue. This visible part ranges from 400 to 740 nanometres. Light waves in the 400–500 nanometres range are purples; those in the 580–740 nanometres range are reds; and the rest of the rainbow colours lie in between.
This visible part ranges from 400 to 740 nanometres. Light waves in the 400–500 nanometres range are purples; those in the 580–740 nanometres range are reds; and the rest of the rainbow colours lie in between.