 CHAPTER 3
CHAPTER 3  CHAPTER 3
CHAPTER 3 Size and shape are inextricably connected. One of the most elementary ways we can show that size is the supreme arbiter and has a profound effect on shape is to consider the relation between weight and strength. Galileo first demonstrated the principle elegantly in his book Dialogues concerning Two New Sciences, which he wrote after he was tried and condemned by the court of the Inquisition in Rome for having committed heresy in supporting Copernicus’s heliocentric view of the heavens—that our world was not the center of everything. He wrote the book while in house arrest in the palace of the archbishop of Sienna, but he was forbidden to publish anything in Italy. An opportunity arose when the Dutch publisher Louis Elzevir came to Italy in 1636 and managed to take with him, on his return home, the then completed part of the manuscript. The subjects, mechanics and motions, were ones that Galileo had been working and lecturing on for many years, and now it was brought together in what he (and others) considered his most important work.
The book was written in Italian, and there is an excellent translation in English.4 The science is presented in the form of a dialogue between three men: Salvati, Sagredo, and Simplicio. Today it is so unlike the way we present science that it seems rather contrived, but the main points are made with the utmost clarity, often illustrated by diagrams and drawings. There are four books, or Days, and in Day Two Galileo expounds the basic relation between size and shape. His main point is that the weight of any body is a cubic function, that is, the cube of its linear dimension, be it height or width. Strength, on the other hand, is a function of the cross-section area, that is, the square of the linear dimensions. Let me give you the consequences of this difference in his own words, which he relays through Salvati:
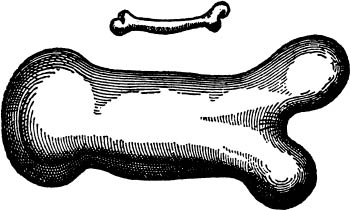
From what has been already demonstrated, you can plainly see the impossibility of increasing the size of structures to vast dimensions either in art or in nature; likewise the impossibility of building ships, palaces, or temples of enormous size in such a way that their oars, yards, beams, iron-bolts, and, in short, all their other parts will hold together; nor can nature produce trees of extraordinary size because the branches would break under their own weight; so also it would be impossible to build up the bony structures of men, horses, or other animals so as to hold together and perform their normal functions if these animals were to be increased enormously in height; for this increase in height can be accomplished only be employing a material which is harder and stronger than usual, or by enlarging the size of the bones, thus changing their shape until the form and appearance of the animal suggests a monstrosity. (Fig. 7.)
Salvati then goes on to say that because of this weight- strength relationship, a smaller animal will have a greater relative strength, which he illustrates by pointing out that “a small dog could probably carry on his back two or three dogs of his own size, but I believe that a horse could not carry even one of his own size.”
Figure 7. A drawing From Galileo’s Dialogues concerning Two New Sciences, showing how a normal bone (top) would need to be increased in thickness (bottom) if the weight of the animal were much greater.
Simplicio then says he doubts this statement is true because of “the enormous size of certain fish, such as the whale.” He was a bit off calling a whale a fish, but Salvati has a ready answer to this welcome question: by being submerged in the water, a whale is “making their bones and muscles not merely light but entirely devoid of weight.”
The consequences of the weight-strength ratio being influenced by size are large, and here I will stress the living world rather than that of the engineer, although both are equally affected. Beginning with the animal world, the fact that a small gazelle has slender, graceful legs, and an elephant is endowed with great stumps is precisely what Galileo would have expected. If there were elephants on the moon, which is smaller than the earth and therefore the gravity is less, one could imagine that they would have evolved into gigantic gazelle-like beasts. Yet another way to look at the same point may be found in that splendid chapter in D’Arcy Thompson’s book On Growth and Form. He points out that the size of a terrestrial vertebrate is reflected in the relative weight of its bones, and consequently, “bones make up some 8 per cent of the body of a mouse or wren, 13 or 14 per cent of goose or dog, and 17 or 18 per cent of the body of a man.”
As Galileo makes clear, the shape of trees is equally affected by size, and we expect a giant sequoia to have, like the elephant’s foot, a huge wide and solid base compared to a smaller tree. The difference can even be seen in the growth of a single tree: a maple starts off as a slender, willowy sapling that, with increasing size, becomes disproportionately thick in its trunk and limbs. The same principle can be seen in fruit of different sizes. The strength of the stem of each fruit will depend on its cross-section area, so the stem of an apple will be disproportionately thick compared to that of a cherry. The stems of coconuts are very thick and sturdy, while watermelons have solved the problem by lying on the ground; their solution is somewhat analogous to that of whales, for in different ways both have largely circumvented the pull of gravity. The relationship between strength and weight can be expressed in a simple equation (to which we will return):
(1) Strength ∝ Weight2/3
(∝= varies as).
To return to Gulliver’s Travels, let us reexamine Jonathan Swift’s work in the light of our discussion; it is a good way of illustrating the principles laid out here.
Let us assume that Lemuel Gulliver was 5 feet and 8 inches tall. The Lilliputians were one-twelfth his height, which would make them just shy of 6 inches tall—roughly the size of a mouse. This would mean that their legs and their bones could be relatively slender compared to normal human beings; they could have stick legs like a sandpiper that runs along the edge of the water on the beach. After all, if Gulliver weighed about 150 pounds, then the similarly constructed Lilliputian would weigh less than a pound. The result would be that he would not be similarly proportioned to Gulliver, for legs like match sticks would be quite sufficient to support him.
The speed of running of Lilliputians, while no doubt slower than that of Gulliver, would not be that far behind because their legs will move much faster (like the sandpiper), and for one of Gulliver’s normal steps they would have taken many. All the movements of the smaller homunculus would be more rapid than our movements, just as the wing beat of a small hummingbird is far quicker than that of a swan.
We know that rats and shore birds have lungs, a heart, gut, and circulatory system as we do. The only difference is that the proportions of these parts of their anatomy will differ from ours, for they have relatively less work to do because of their smaller size: the heart pumps the blood into a smaller set of blood vessels and therefore can be smaller. Because the Lilliputians have smaller vocal chords their voices will be high and squeaky, and they would talk with great rapidity, so much so that it might be difficult for Gulliver to hear and understand them.
The reverse of these arguments would apply to the huge Brobdingnagians, who, to begin with, would have extremely deep and slow voices. Swift describes the first of the giants he encountered this way:
He appeared as Tall as an ordinary Spire-steeple; and took about ten Yards in each Stride, as near as I could guess. I was struck with the utmost Fear and Astonishment, and ran to hide my self in the Corn, from whence I saw him at the Top of the stile, looking back to the next Field on the right Hand; and heard him in a Voice many Degrees louder than a speaking Trumpet; but the Noise was so High in the Air, that at first I certainly thought it was Thunder.
If their proportions are those of a normal human being and they are twelve times taller than we are, which would make them 68 feet tall, they would weigh between 12 and 13 tons. This makes them lighter than the larger dinosaurs, but about ten times heavier than a large elephant. Like these large beasts, the Brobdingnagians would require greatly thickened legs and leg bones, so much so that they would no longer appear like normal human beings, but more like victims of advanced elephantiasis of the legs.
Strength is not the only feature that affects the shape of an organism as its size changes. Another most important one is diffusion, which, as we shall see, has a powerful effect on shape. Diffusion is always through surfaces, and since the surface is a square of the linear dimensions and the weight is the cube, we can again write a simple equation to express the relation of surface to size:
(2) Surface ∝ Weight2/3.
Most organisms, including ourselves, depend on oxygen for their existence; every cell of our body needs to have oxygen diffuse into it in order for it to function and stay alive. Diffusion is the random movement of molecules from regions of high concentration to low, so that ultimately, at equilibrium, they are evenly spread out. Molecules zip around at great speed when they collide with one another. In regions where they are numerous, they collide often and have more luck spreading when there are fewer ones to bump into. A way to look at this might be to imagine a tea bag placed in a glass of cold water and all currents, convection and otherwise, are eliminated. The brown color released from the tea bag is first concentrated in and around the bag itself, but eventually the brown color will be evenly distributed; it has diffused from the bag.
If oxygen has to get into an animal or to its cells it must diffuse through a surface that, like strength, is a function of the square of the linear dimensions. This can be illustrated in a simple fashion. Consider two hypothetical spherical organisms, one the size of a sand grain a millimeter in diameter, and the other the size of a golf ball. Oxygen is a small molecule that diffuses rapidly, and in a matter of seconds it can penetrate a millimeter into a living body. The middle of the tiny sphere is close enough to the surface so that as soon as it uses up its oxygen it will be replenished by a steady diffusion of more from the surface. But in the golf-ball beast it would take an hour or so to reach the interior, and we all know that would not do—if we are deprived of oxygen we die. In fact, we are so tuned in to this problem we cannot even hold our breath for more than a few minutes; we have a built-in mechanism that forces us to start breathing again.
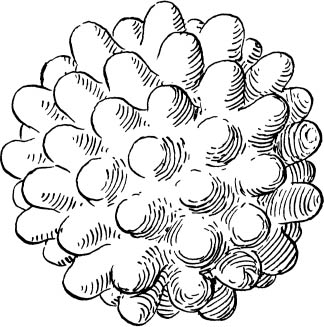
The only way an organism larger than a millimeter can survive is to bring its interior near the surface everywhere so that no part is farther away than one millimeter. This can be managed by radically increasing the amount of surface, making many convolutions and folds so that every point in the interior is close to the surface. Instead of a perfect, smooth sphere, we now have a golf ball with an incredibly wrinkled surface (fig. 8). Or to put it another way: because the enlarged spherical organism cannot exist in it original form, it must grow more skin, and the result will be a radical change in shape.
Shape changes caused by diffusion problems can be far more revolutionary than this transformation. Much of the shape of human beings is molded by the diffusion of oxygen and the problem of getting it to our inner cells. To achieve this end, we have a marvelously complex set of devices. We have lungs that bring the oxygen close to our blood vessels where the hemoglobin can pick it up; we have a circulatory system with a heart, a most ingenious and durable pump that pushes the oxygen-containing blood into the minute vessels, or capillaries. They permeate all parts of our bodies and lie close—well within the one millimeter limit—to each of our deep-seated cells. Note that the lungs and the blood vessels have vast surfaces; like our wrinkled golf ball, they keep the surface-volume ratio constant despite our large size. More than that, they have set up a system of motion so that the lungs move the air in and out, and the beating heart keeps the blood circulating. The movements supplement diffusion so that fresh oxygen can be rapidly picked up in the lungs and delivered to the cells.
Figure 8. The shape of a hypothetical golf-ball-size organism that requires every internal cell to be near the surface. (Drawing by Hannah Bonner)
Oxygen is not the only bit of fuel needed to keep a large animal going. Food to burn for energy is needed as well, and it also must be taken in through a surface. Our food canal, from our mouth to our anus, is a tube, a surface through which the broken-down, dissolved food has to pass. The greater the surface of our gut, the greater our ability to absorb food. Because surface increases as the square, the larger the size of an animal (which increases as the cube), the greater must be the surface of the gut to keep pace in order to maintain a constant surface-volume ratio. This change can be seen clearly if we compare ourselves to minute nematode worms that are little bigger than an eyelash. Their gut is a straight tube, but ours is a tortuous, highly convoluted tube many times our length. Furthermore, our small intestine is not a simple tube, but has a forest of tiny papillae that stick up into its inside, each one greatly increasing the gut surface to absorb more food. We have also developed glands: the salivary glands, the pancreas, and the liver that produce batteries of enzymes to break down the food into small molecules such as simple sugars and amino acids, so that they can diffuse through the gut wall more easily.
All these devices found in a large animal are present to get energy from food by combusting it with oxygen in all the cells of the body. This job calls for lungs, a circulatory system, and a highly elaborate gut and its associated glands. From this necessity it is clear that size increase requires quite extraordinary changes in shape. In no way do we resemble a smoothly spherical organism, nor even a wrinkled golf ball; we have evolved into a far more complex shape largely because of the increase in our size.
Viewed through the eyes of Lemuel Gulliver, the Lilliputians need only a comparatively small number of blood vessels to reach all the tissues of their bodies. The Brobdingnagians, on the other hand, need a vast number, and their guts will have to be disproportionately extended with an immense increase in their surface area to absorb sufficient food to accommodate their giant size.
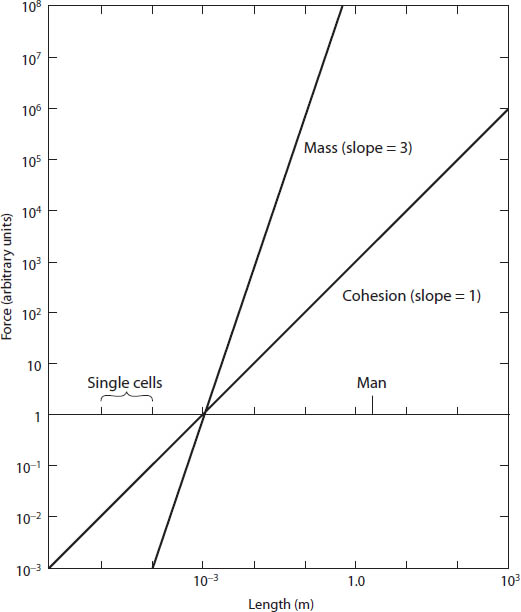
It is possible to put large and minute organisms in quite separate baskets; there is a fundamental rift between the living worlds of the large and very small, even though it is obviously a continuum if one goes from large animals and plants to small ones; organisms come in all sizes in between. The reason for arguing for such an artificial separation of two size worlds has to do with the forces affecting the totally different size levels. The effect of gravity becomes progressively insignificant the smaller an organism, while molecular forces come to the fore and play an increasingly important role. Even though these forces decline and rise smoothly as the size changes, one can artificially designate a critical point where one jumps from one world into the other. Presently I will give an example, but first let us look at a simple graph (which I have taken from a splendid paper by F. W. Went) in which the force of gravity and the force of molecular cohesion are plotted on a size scale (fig. 9).5 In terrestrial organisms, gravity varies with the weight, and therefore with the cube of the linear dimensions, while the force of attraction between surface molecules varies roughly as the linear dimension. This approximation takes into account the forces of attraction between molecules and the distance between the molecules on the two surfaces. The important point is that the cohesion curve is less steep than the curve for gravity. The figure is drawn in such a way that the lines for gravity and cohesion arbitrarily intersect for an organism roughly a millimeter in length; above that point, the force of gravity increases more rapidly than that of cohesion, while below that point smaller organisms see a declining effect of gravity compared to that of cohesion. In other words, minute beasts hardly feel the effects of gravity at all; rather, the forces of cohesion affect every aspect of their existence.
Figure 9 (facing page). A synthetic graph comparing how different size terrestrial organisms are affected by gravity and cohesive forces. Forces due to weight vary as the length cubed, while the forces due to molecular cohesion average out very roughly to be proportional to the length. The lines are arbitrarily placed so that they cross for organisms 1 mm in length, and it can be seen that in larger organisms the forces of gravity (which vary with the mass) play a far more significant role than the forces of cohesion, while for smaller organisms cohesion forces dominate. (This figure is based on one in F. W. Went, American Scientist 56 [1968]: 400–413)
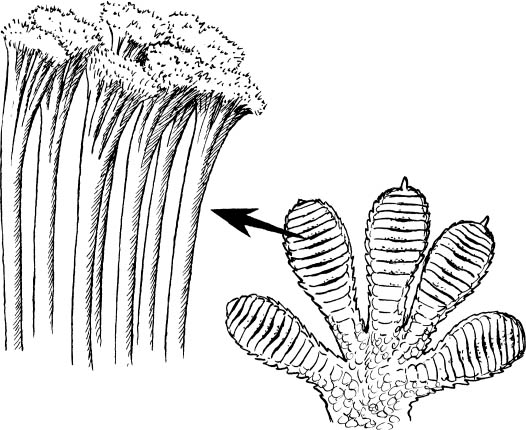
A fly or a gecko can easily walk up a wall, something we humans cannot even begin to manage. The reason is that these creatures’ weight is negligible and the bottoms of their feet are specially constructed with many hairs so that they can have very intimate contact with all the minute nooks and crannies of the rough surface of the wall. This feature can be seen especially clearly in the foot of a small gecko (fig. 10), indicating that the molecules in the feet and those in the wall become very close to one another and the force of that molecular attraction increases the closer the molecules of the two surfaces are. The smaller the beast, the greater the effect of cohesion compared to that of gravity.
From this we see that small organisms live in a very different world than large ones; it is physically different. Besides the increase in the importance of molecular forces, it is also the world of low Reynolds numbers, a world that does not exist for larger forms.
The Reynolds number is a convenient way of describing the immediate environment of a moving body. It is a way of showing that a swimming bacterium is in a very different environment than a swimming whale even though both are swimming through water. The vast difference in their size, for purely physical reasons, affects their locomotion. A whale has a far greater inertial force, and if it suddenly stops swimming it will glide on for some time. Because of its great mass it is endowed with a large amount of momentum. On the other hand, when a minute bacterium stops swimming, its mass is so small that it stops in a split second.
Figure 10. The foot of a gecko and a high-power microscopic view showing the mass of very fine hairs that can make intimate contact with a solid surface. (Drawing by Hannah Bonner)
In the latter part of the nineteenth century, Osborne Reynolds, a distinguished engineer, came to understand this problem in a deep way. But here I will describe his sophisticated principle in a very informal way that will no doubt make a physicist cringe.6 Nevertheless, its essence can be expressed as follows:
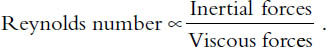
Both the whale and the bacterium swim in water so the denominator in the equation above will be the same for both. However, the inertial forces for the whale will be huge compared to that of the bacterium, with the result that the whale will have a correspondingly large Reynolds number while the bacterium’s will be small. Microscopic organisms live in a world of low Reynolds numbers.
To give one a feel of what this means for minuscule organisms, the noted physicist Edward Purcell painted the following picture.7 Imagine a man swimming in a liquid that produces the same low Reynolds number as would exist for his own sperm. It would be like trying to swim in thick molasses in which one was not allowed to move one’s arms or legs faster than the hands of a clock. Under these rules, if one moved a couple of meters in a couple of weeks, one would qualify as a low Reynolds number swimmer. This vivid picture gives one an idea of how the changes in the inertial forces affect a bacterium; it is indeed a different world. Purcell says, “Motion at low Reynolds numbers is very slow, majestic and regular.”
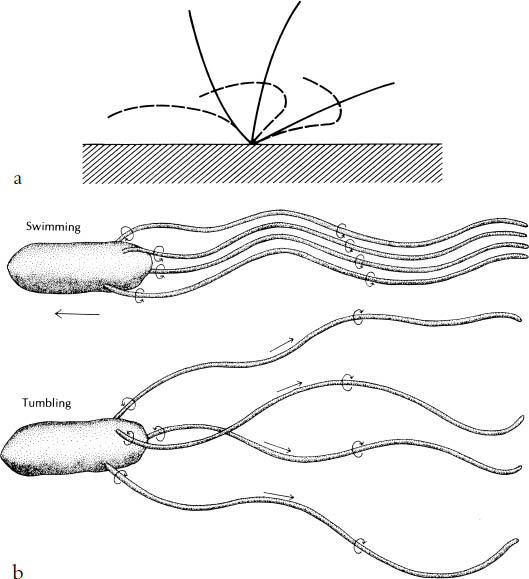
Swimming microbes have whiplike structures called cilia or flagella that propel the organisms through the water. In single-cell protozoa, cilia move the organism by behaving like sophisticated oars; they are flexible and bend on the back stroke, but are stiff when pushing forward and provide the cell with a powerful thrust (fig. 11a).
Let me interject an aside here. When I was a student and first saw, through the microscope, these little whiplike structures move one-cell organisms across the field of vision, I marveled at how fast they traveled. Later I realized that they were in fact moving very slowly and the magnification of the microscope had fooled me. After all, speed is distance divided by time (as in miles per hour), and the high power of the microscope made the minute distance appear very large. For me it was part of growing up: I had not realized that what I was seeing—those protozoa zipping around at an incredible rate—is an optical illusion created by the magnification of the microscope. It was rather disappointing to learn that the microworld was not dazzlingly fast but rather sedate.
Bacteria are very much smaller and for that reason have a different set of problems. Not only are they in an environment of even lower Reynolds numbers, but also they are so small they cannot construct the relatively huge and complex oarlike cilia or flagella of larger microbes. They are in the nanoworld, having very thin flagella of a simple construction that produces locomotion in quite a different way. Each flagellum is attached to a rotor that goes around and around, causing the flagella themselves to rotate like a ship’s propeller, and this action thrusts the bacterium forward (fig. 11b). There are many remarkable aspects of this ingenious method of dealing with the difficulty of moving a minute body in conditions of very low Reynolds number.
Figure 11. Microbe propulsion. (a) The flexible oar of ciliate. The solid curves show successive positions of the cilium beating downwards for forward movement; the broken curves show the cilium on the reverse stroke (from T. A. McMahon and J. T. Bonner, On Size and Life, 1983). (b) A bacterium (E. coli) swimming forward (top); and how a sudden reversal of rotation causes the flagella to splay out and the cell to tumble (bottom). (After J. Adler and H. C. Berg; from Bonner, The Evolution of Culture in Animals, 1980)
The discovery that bacteria move this way was made not that many years ago. It was first predicted on theoretical arguments based exactly on what we have just seen: the special problems connected with low Reynolds numbers. This prediction was followed by a very clever experiment in which the flagella were trapped on to a specially prepared sticky microscope slide with the amazing result that the bodies of the bacteria could be seen, through the microscope, to rotate continuously. They possessed a true rotor, something not previously known to exist in the living world.
We often congratulate ourselves on our inventiveness that places us above all other animals. We not only discovered how to control fire and make use of it, but we also invented the wheel, that clever device that allows us to move heavy loads. It is usually claimed that no other animal can manage this feat, but as we now realize, it was in fact devised billions of years before Homo sapiens appeared on the globe. As we have just seen, bacteria invented the wheel, and exploited theirs in a most sophisticated manner.
To illustrate this sophistication, let me briefly explain how bacteria can orient themselves in their aqueous environment. It is well known that swimming bacteria will go towards food and away from noxious substances; now we know how they do it. They have molecules on their surface that can adhere to specific external chemicals, and if they happen to be swimming in the direction towards food, then there is a molecular signaling mechanism that tells the rotor to keep going. If, on the other hand, the food molecules become increasingly less abundant because they are swimming away from the food, then the rotor gets a new message and abruptly reverses its direction. This action causes the whole cell to stop, to tumble, and when it starts up again there is a good chance it will be pointed in a new direction, which, with luck, might be the direction of the energy-rich food. So bacteria not only can move, but they are blessed with the ability to move in a direction that is to their advantage. And they have managed all this without a brain!
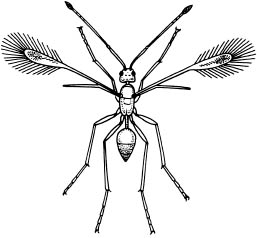
A low Reynolds number environment also influences very small flying insects. Their force of inertia is insignificant, with the result that air is also like molasses. As a consequence in more than one group of insects, their wings have become transformed into what look like minute feather dusters (fig. 12). These so-called fairy flies (which are in fact minute parasitic wasps) use their wings the way ciliate protozoa use their cilia; they swim through the air.
Figure 12. Among the smallest insects are fairy flies, which are actually tiny wasps. Their body lengths can be less than half a millimeter. The wings are slender stalks, whose margins are fringed with hairs. (From T. A. McMahon and J. T. Bonner, On Size and Life, 1983)
It is clear that size, from the pure point of view of physics, has an enormous effect on living organisms. It constrains, or even controls, the shape of an organism, including its internal structure. It determines what is possible and what is impossible. And as we have also seen, it can affect the behavior of animals as well as their locomotion. In the next chapter we will see that size is also directly related to the degree of division of labor.
Note that if one compares small and large animals or plants, one aspect of their relative shapes is a change in proportions. This is not surprising because, as we have seen, in order to exist those properties that are related to the square of the linear dimensions, such as strength and the diffusion processes related to energy utilization, must, with increased size, rise more rapidly to keep pace with an increase in volume or mass. It shows itself by a disproportionate increase in the thickness of limbs and a more convoluted gut in larger animals.
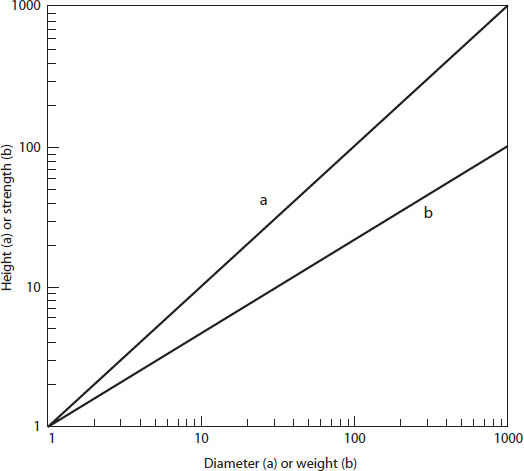
There is a simple and elegant way to describe the changes in proportion to size, known as allometry.8 Consider our earlier example of the bones of small gazelles and large elephants. If they were similar in all respects, then an elephant would have legs that looked like a photographic enlargement of a gazelle. If this were so and we plotted the height of animals of different sizes against the diameter of their bones, we would get a straight line with a slope of 1 (fig. 13). That is, the diameter varies as (∝) the height:
Diameter ∝ Height.
As we learned from Galileo, they are not similar because the overall size (weight) of the animal rises as the cube of the linear dimensions, while the strength of the bones rises as the square of the linear dimensions. That is,
(2) Strength ∝ Weight2/3.
This equation neatly expresses how the strength-weight affects the proportions. It is convenient to plot this relationship on the graph as logarithms, for the exponent of 2/3 will produce a straight line whose slope is 2/3 (fig. 13).
Figure 13. A log-log graph showing allometric relations. (Line a) If diameter is plotted against height in geometrically similar organisms of different sizes, then the slope of the line equals 1. (Line b) If strength (which is diameter squared) is plotted against weight, which is the cube of the linear dimensions, the slope of the line equals 2/3.
There are many ways to illustrate the usefulness of allometry, and I shall pick two: size changes during human growth, and relative brain sizes.
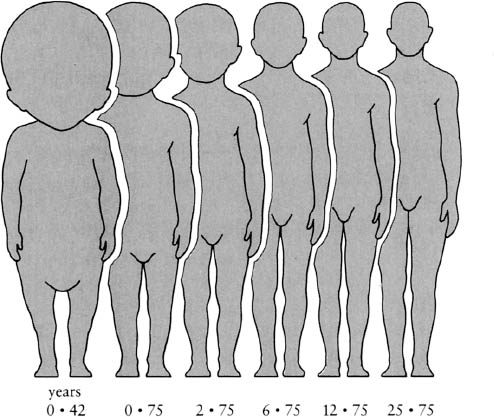
Proportions change as animals and plants grow. For instance, a human baby is quite different in its shape from an adult—there is a continual change in proportions as we grow. The best way to show this is to draw the various stages of human growth adjusted to the same height (fig. 14). If one looks at a small baby its head seems enormous compared to that of a grown-up. In fact, a baby’s head accounts for 34 percent of its height, but for a mature person the head is only 13 percent of its height. The body grows in length about twice as fast as the head in the growth of a human being.
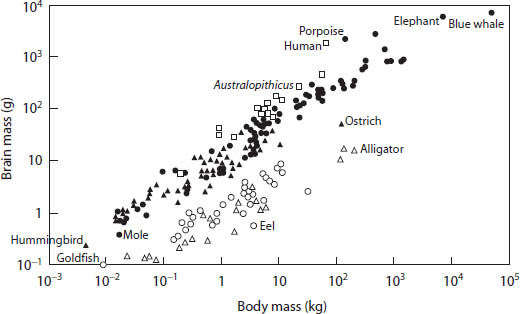
Brain size is a subject especially dear to us human beings because we are so pleased and self-satisfied with our very large brains. If one plots the weight of the brain against the body weight of various vertebrates, one again finds a 2/3 relationship (fig. 15). The bigger the animal, the bigger the brain, although it is not directly proportional, for as the body size increases the brain size increases less (roughly 2/3 of the increase in body size). It is not clear why the brain of large animals should be relatively smaller than those of little animals.
Bigger brains generated an interesting idea by a distinguished German evolutionary biologist named Bernard Rensch some years ago.9 Rensch argued that being bigger meant having a larger brain, which meant that larger animals must be smarter. Increased size automatically conferred such an advantage, and this was one of the reasons there are often evolutionary trends towards larger size. Rensch used the ability to learn, and retention of what is learned in the form of memory, as his measure of intelligence. To examine his idea he compared similar birds of different sizes—bantam chickens, normal-sized ones, and giant breeds. He gave them a simple learning procedure followed by periodic testing to see how long they remembered what they had learned. He found that the larger birds could learn more and retained the newly acquired information for a longer period of time. Having a big brain did seem to provide an advantage. He realized that all of this fits in with the old adage that an elephant never forgets, and proceeded to make some interesting observations on circus elephants, and indeed they had a remarkably long memory for some card puzzles he taught them. I am reminded of a story my father used to tell us children about the circus elephant that recognized his old trainer in the audience and lifted him up with his trunk and moved him from the fifty-cent seats to the dollar seats.
Figure 14. Changes in body proportions in a human being with increasing age. (After Wells, Huxley, and Wells, 1931; from T. A. McMahon and J. T. Bonner, On Size and Life, 1983)
Figure 15. Brain size of 200 species of vertebrates plotted against body size on a log-log graph. Primates are open squares; other mammals are solid dots; birds are solid triangles; bony fishes are open circles; and reptiles are open triangles. (After H. J. Jerison, The Evolution of the Brain and Intelligence, 1973)
Rensch once came to our university to lecture on all this, and in the discussion period I asked him if perhaps it was not intelligence, but because the animal’s metabolism is slower the larger the animal, the elephant was simply slow to forget. Professor Rensch got quite irritated with this fresh question from a young man, but to this day I wonder if I did not have a point. Later I plan to talk about size and metabolic rate in some detail. I cannot believe that Rensch’s intriguing idea has been an important factor in evolution. There are too many small animals of remarkable intelligence, such as parrots and crows among numerous others. One cannot imagine that the size of elephants increased because of a selection advantage of being better at learning and remembering things.
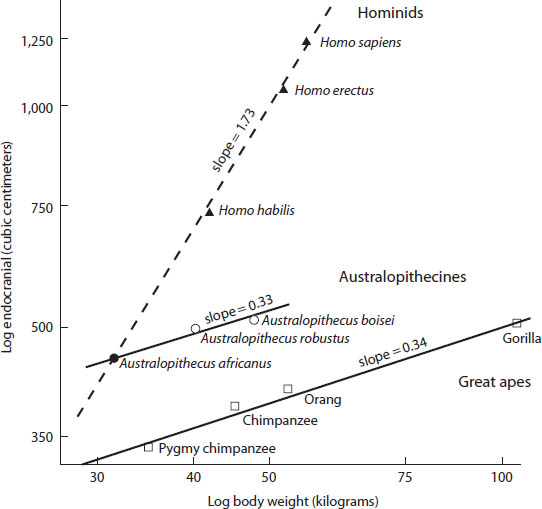
If we turn to primates, we see a similar relationship: the larger the monkey or ape, the larger the brain. However, if we look to the evolution of human beings, from Australopithicus to Homo, then we see an amazingly steep rise of brain size relative to body size. Instead of the brain enlarging at a slower rate (2/3 or less) than the body size, it rises at a much greater rate—almost two times that of the increase in the body size (fig. 16). Note that if we compare Australopithicus species of different sizes, the allometric relation of their brain size increase is modest, mirroring that of apes. In other words, in our evolution from a common ancestor with apes, one great difference from them is our remarkable increase in brain size. We did indeed become the brainy animal. The only other animals that had a similar increase in relative brain size are porpoises and whales; we are only beginning to understand something of their complex behavior.
Figure 16. The endocranial volume plotted against the body weight of great apes, Australopithecines, and the Homo line on a logarithmic scale. For the fossil forms the weight is, of course, only an estimate. (Redrawn from D. Pilbeam and S. J. Gould, Science 186 [1974]: 892–901)
As one might expect, there has been a considerable amount of speculation as to why brain size might have made these sudden spurts. One of the earlier hypotheses centered around the idea that it had something to do with needed extra brain power to make and use tools. For various reasons this notion has fallen in disfavor, largely because tool making has come to light not only in great apes but in many animals. For instance, Caledonian crows will not only use a piece of wire to extract food from a long, narrow glass vessel, but after a few unsuccessful tries they will carefully bend the tip of the wire to hook the food at the bottom.10 It seems unlikely that devising and using tools would drive brain evolution to any great degree; those Caledonian crows have the same size brain as other crows. Another idea is that the brain increase is associated with the rise of human language, which implies that in terms of Darwinian natural selection, being able to speak more effectively conferred a selective advantage, and the better communicators had more offspring to whom they passed on those skills. The difficulty with this suggestion is that an increase in brain size along with communication skills is merely a correlation; it is hard to see what comes first—what caused what.
A related and more general argument comes from the modern study of primate social groups, from lemurs to chimpanzees. Clearly, reproductive success in those groups does not just depend on brute strength, but on careful manipulation of one’s position within the group. This in return requires managing alliances with others in the group to strengthen one’s social position; it is what Frans de Waal has called, based on his fascinating studies, “chimpanzee politics.”11 This whole idea has now taken hold and has even been given a name: “Machiavellian intelligence.”12 According to this hypothesis, it is the social interactions that are key, and the individuals who can influence others to help them in their power struggle are the ones blessed with reproductive success. It is of special interest that recent work on the large-brain dolphins shows some strikingly elaborate social behavior; maybe they too have become big brained due to Machiavellian intelligence.
Because a large brain is such a striking character of Homo sapiens, great efforts have been put into attempts to tell us something about differences among humans. Using head or cranium size as a measure of brain size, there is great variation in our species. Those who have some sort of dubious agenda have especially loved this: those who scrounge for evidence to prove that a race with bigger heads is superior in intelligence to another with smaller ones. It has even been used to argue that men are superior to women because they have, on average, larger bodies and therefore larger heads!
Over and above the fact that this approach is distasteful, for many reasons it has proved to be a fruitless and muddled enterprise. To begin with, variation in brain size, as measured by volume within the cranium, very likely is too rough an approximation of the number of neurons in the brain and their pattern of interconnections. Variation in how closely packed the nerve cells are in different parts of the brain, and the degree of folding of the brain surface, will not be reflected in head size. In a splendid play about the famous mathematician Alan Turing, who did so much to launch computers and artificial intelligence, there is a scene where he goes back to his old school to give a lecture about the brain, which he describes to the boys as looking like a lump of porridge. In this chaos we know little about how the millions of neurons, and even more importantly how their interconnections, lead to intelligence. To make the point in a forceful way: thanks to the pioneering work of Karl von Frisch, we know that honeybees have an extraordinary ability to memorize information and even communicate that information to their fellow workers, yet the brain of a bee is minute with a correspondingly minute fraction of the neurons found in any mammal, let alone human beings. This is a fascinating area of inquiry, and despite recent considerable advances, we have yet to scratch the surface.
Furthermore, our crude methods of measuring human intelligence are woefully inadequate. IQ tests tell us something about thinking, but one never quite knows what. There is a strong suspicion among many, including myself, that they do not say anything about what we would normally recognize as intelligence, but merely tell us how good we are at taking that kind of a test. I have already pointed out that brain size varies with body size and therefore it is to be expected that women, who are on the average smaller than men, should have smaller brains, but there is no evidence for any significant difference in their mental abilities, even their ability in taking IQ tests.
As one might imagine, there is a great literature on this subject, often polemical. To my mind it can be brushed aside in a simple way. There are many well-known savants who had small skulls containing great minds. An example of such a small-headed person is Anatole France, the French author who is famous for his novels and for having exposed the anti-Semitic injustice of putting Dreyfus in the penal colony on Devil’s Island at the turn of the twentieth century. In looking this matter up, I was delighted to discover that Jonathan Swift of Gulliver’s Travels had an exceptionally large head—twice the size of Anatole France’s!
Let us pause for a moment to comment on the two rules that have just been discussed. Both strength and surfaces vary with the weight in the same way:
(1) Strength ∝ Weight2/3
(2) Surface ∝ Weight2/3.
Later we will see that other important properties of living organisms vary in a similar way, although the exponents, and therefore the slopes of the lines when plotted logarithmically, vary. The three examples to come are
(3) Complexity ∝ Weighta
(4) Abundance ∝ Weight−b
(5) Metabolic rate ∝ Weightc.
All five reflect the fact that size differences, as indicated by their weight, affect different properties: for a given weight, an appropriate strength is required for support and movement; an appropriate surface, so that an adequate diffusion of oxygen and food substances can reach the inner tissues; an appropriate division of labor, so that the body of the animal or plant can function; and the appropriate rate of metabolism for that functioning. Finally, size affects the abundance of animal and plants in nature: the bigger the organism, the more space it needs. These five rules are central to the argument of this book; they are the pillars that underpin the effects of size on living organisms.