IV
The Second Grade of Individuality and Its Attainment
Question: “What’s one and one and one and one and one and one and one and one and one and one?”
Alice through the Looking-Glass.
Answer (sometimes):
“Each one almost a Whole, yet all but Parts
They have lost self to form a Greater Whole
Far nobler than its sum of single Parts.”
The Green Bayswater.
In Chapter II it was shown that the very existence of the first living individual, the cell, was originally determined by the material properties of living substance. There are large cells and small cells, but, with few exceptions (see pp. 67–68), it is a very limited largeness to which even the largest attain. This limitation, depending as it does upon the surface-volume ratio, is one of the primitive, original attributes of the cell; and to attain size, the cell must in a way do violence to its nature, somehow modifying its surface-tension, overcoming its natural tendency to the spherical, so as to keep its absorptive organ, the surface layer, large enough to supply the demands of the inner mass.
Why, however, should the cell not be content to stay small—what is it to gain from size that it should strive after it? One is apt to think of size as a rather unimportant element in life. With the example of the field-mouse and the elephant, both built so closely on the same type, the wren and the albatross, one comes to think of a model of organization which can be fitted at will on to whatever bulk of living matter is desired. Within wide limits this is true, no doubt, but limits none the less there are.
Many Neo-Darwinians, too, argue that adaptation is the great reality gained by organisms through natural selection, and that, therefore, no one species now alive has preference over any other—for to be alive both must be adapted to their surroundings. But to exist and nothing more, to vegetate merely, is not the fate of all organisms. There is a higher and a lower, for some are more independent, more powerful than others.
It is now that the importance of size is seen; for increase of size means increase of independence. Most of the forces of the outside world act only on the surface of the organism; but its own forces spring from the whole mass of its substance. The energy necessary for action is let loose by the chemical breaking-down of the molecules of protoplasm and by their combination with oxygen. This, in a primitive cell, is a function of all the molecules, and so of its total bulk. In the higher animals, where locomotive power is delegated to the muscles, the relation still holds good; the three dimensions and so the shape of the thigh-muscles of a jerboa and a kangaroo are approximately the same, and so the surface-volume ratio will hold accurately. If the length of the kangaroo is ten times that of the jerboa, then the surface of his thigh-muscles will be a hundred, the bulk a thousand times as great. Of the outside forces (all antagonistic or at best passively resistant to the organism) that of gravity only is proportional to its mass. If it alone were to be considered, size would make no difference to the animal’s movements—the weight to be moved would increase proportionately with the forces that were to move it, since both are proportional to the mass of the whole.
In reality, even if we consider locomotion alone, the resistance of the medium—air, water, or earth—in which the creature moves, is equally important with gravity. Everyone knows how much harder it is for a thin, loose-built man than for a close-knit, compact one of equal weight, to make headway in a gale of wind. That is because the pressure of the wind is proportional to the surface exposed, and the thin man, with relatively more surface exposed, has less muscle with which to drive his body onward.
The home of all primitive life was the water; and the resistance of water is immensely greater than that of air. The disproportion between inner and outer force is here so great that it is as impossible to think of any single-celled animal swimming against the most sluggish river as it is to imagine a butterfly poised steady in a twenty-knot gale.
Once more we see the importance of the surface-volume ratio: but what it preaches now is for the organism in direct contradiction to its earlier lesson. Then it said, “thus far and no further, on peril of starvation.” Now it warns, “stay thus small, and be condemned to continue the sport of the elements.”
How is life to escape from this quandary? She may be content to remain small, like all the present-day Protozoa. Many of these have attained the most amazing complexity for their size; but there are physical limits to the amount of structures that can be contained in a small fraction of a cubic millimetre of substance, and this way has led up a blind alley. One tribe of plants, the Siphoneae, has made a brave attempt to gain size while still remaining a single cell.1 A plant of Caulerpa, for instance (one of these sea-weeds), may have several square feet of surface, and in spite of being one continuous piece of protoplasm with a wall of cellulose round it, is differentiated into organs resembling in appearance and no doubt in function the stem, leaves, and roots of higher plants. Needless to say, it has only been able to attain this relatively huge bulk by restricting itself to growth in two dimensions only, and is quite thin and plate-like throughout.
What possibilities of development lay along this line we cannot say; all we know is that actually it has not led far. The real advance has been made in a quite different way; by keeping the cell’s original form and plan, but joining up a number of them together so that each preserves a considerable measure of independence, and is yet subordinated to the good of the whole. This resulted in the metazoan type of structure, where the individual is built up out of a number of cells instead of one.
As an example of a simple metazoan, we had better take a primitive sponge. Among sponges, Clathrina blanca is one of the most primitive. A graceful vase-like creature, pure white, with a long stalk of attachment, and a mere fraction of an inch in length, it obtains its food, like the majority of sedentary aquatic animals, by producing a current. A stream of water can easily be demonstrated passing out of the circular mouth of the vase, and, with a little more trouble, can be seen to enter by a number of quite small holes scattered over the walls. The current is produced by the cells lining the central cavity (Fig. 6): these stand side by side like sacks in a granary, their free upper ends tapering very slightly, and then truncated at the top. The flat top of each is surrounded by the most remarkable transparent cylinder, a mere film of protoplasm, yet beautifully round, and capable of being drawn in at will, or protruded till it equals the cell in length. This is called the collar, and in the centre of it there springs from the cell a long vibrating lash or flagellum, of uniform thickness throughout, and also capable of retraction within the body of the cell. Usually however it is very much in evidence, beating several times a second, and so producing the current, from which food is taken up and digested by the collar-cells. The bases of these cells rest upon a thin layer of jelly—dead stuff secreted by the living cells, and serving, like the somewhat similar gelatinous tissue we shall see in Volvox, for the common support of the separate cells. On the outside of the jelly is the dermal layer of flat polygonal cells, fitting together like a mosaic of tiles. The pores through which the current enters are perforations in the bodies of cells, of a third kind large and contractile, each of which stretches drainpipe-wise from the outer world to the central cavity. Embedded in the jelly itself are other supporting structures—three-rayed spicules of carbonate of lime, and through it wander at will a number of amoeboid cells, having much the same appearance and functions as our own white blood-corpuscles, except that from their ranks are recruited the germ-cells, male and female; here, therefore, we have the unusual2 spectacle of the germ-cells being pressed into the service of the individual.

Figure 6
Clathrina coriacea, Mont. Two sections of the body-wall. E, not quite fully expanded; the collar-cells line the cavity of the sponge, and show collar and flagellum. F, very much contracted. The collar-cells have withdrawn collar and flagellum, and are lying in irregular masses behind the layer of immigrated pore-cells. am.c, amoebocytes; c, collars of choanocytes (ch.c); d.ep, dermal epithelium; fl, flagella; p.c, pore-cells; sp.c, spicule-cells. (Highly magnified.) (From Minchin.)
Here is obviously a unity, an individual of a higher order than the cell. Its forms and its functions both depend as much upon the way the component cells are arranged as upon their structure; from an examination of a single one of its cells, or even one of every kind of cell, you could deduce very little about the properties of the ordered whole. That whole is greater than the sum of its parts; for the problem is one of combination, not of mere addition. In spite of this the cells have preserved a very large amount of independence, and indeed do most forcibly present themselves to the mind as bands of beings like ourselves that have voluntarily enlisted under some beneficent tyrant of a general. That analogy, between cells and men, body and state, has been too often and too far pressed; its incompleteness is at once grasped with the realization that no such general does or can exist for the cell-battalions to obey.
What the bond is that keeps them together, what the force that orders them—this is still one of the most mysterious problems of life. We must first grasp the extent to which minor individualities can persist within the major—see how that centralized empire, the body of one of the higher animals, was in its origin a federation, not a tyranny.
In Clathrina, the cells’ independence is largely realized by mere inspection. The collar-cells only touch each other with the lower part of their bodies, and when the sponge contracts, as it does in unfavourable conditions, they—after drawing in their collars and flagella out of harm’s way—are actually forced over each other, so that instead of a single unbroken layer there is an irregular collection of cells filling up almost the whole of the central cavity. Whether when the sponge expands again they always fit themselves in between their former neighbours cannot well be proved or disproved, but seems at least unlikely.
The amoeboid cells wander as they please, and the outer or dermal cells, though to be of use to the sponge as protective and contractile tissue they must constitute a single continuous sheet, and so seem merged and lost in the one dermal layer they form, yet show themselves still independent in performing their further function, the secretion of the calcareous spicules. As these are required, single dermal cells break loose from association with their fellows, wander off into the gelatinous ground-substance, and there take up position where the new skeleton is required. Thus, though what they do only has meaning in regard to the whole, the way they do it proclaims them as partially independent beings.
Experiment reveals further lengths of independence—shows the cells capable of veritable insubordination. By means of experiment it has been possible to study the behaviour of the unit parts after the individuality of the whole has been totally destroyed. By chopping the sponge up small, wrapping the bits in the finest silk gauze, and squeezing them, the cells are wrenched from their attachments, and pass through the meshes either singly or at most by twos and threes.3 By varying the method, one can procure, instead of a mixture of all the sorts of cells, a quantity of collar-cells free from all the rest, and it is their behaviour that concerns us now.
No properly conducted cell, one would have thought, could wish to survive this forcible severance from the whole, the body which we are accustomed to think of as constituting the basis of the only real individuality in an animal. These cells, however, are scarcely inconvenienced. After a short period of shock during which collar and flagellum are withdrawn, they begin joining up one with another, forming irregular solid lumps which, gradually hollowing their central parts, are soon transformed into hollow perfect spheres, their walls a single sheet of cells, and the flagella, now active, beating on the outside. The general resemblance to Volvox (p. 79) is striking, and is made more remarkable by the existence of group of Protozoa—the collared flagellates or Choano-flagellata—whose essential structure is identical with that of the collar-cells; if one of these artificially-produced spheres were found in nature, it would certainly be taken for a colony of Choano-flagellates.
Many of these spheres were kept alive for over a month, and there is little doubt that if the right food were found, they could exist indefinitely, though what would happen with the multiplication of the cells and the consequent growth of the spheres it is hard to prophesy. This remains to be tried; but the facts as they stand are interesting enough. For untold generations no collared cells of a sponge have ever existed except as a subordinate part of a whole sponge-body; and yet, if artificially freed from that “harmonious constellation,” they can act independently, can unite into new societies unlike anything known to exist in free nature, and can there subsist for no inconsiderable time.
So much for the independence of the cells: now for their subordination. If, in the experiment narrated above, all the kinds of cells are allowed to remain mixed after their mutual attachments have been broken, we get a result very different from that obtained with the pure collar-cells. First of all, the cells, many of which are still actively amoeboid, and can be seen crawling over the bottom, unite with each other into small lumps and balls. These balls are unlike any organisms known to exist: for, although all their constituent parts are alive, they are without any arrangement and cannot execute any concerted function, Now comes the strange part: this higgledy-piggledy of cells joined up at random is able to reorganize itself, to produce order out of chaos. First of all the collar-cells sort themselves out and form a central solid mass, the dermal cells migrate to the exterior and join up into a single dermal layer. By so doing (though they still resemble no known organism), they have laid down the ground-plan of the sponge, for it is of the essence of sponges to consist of these two layers in this position. The subsequent changes are changes of detail; cells of the outer layer detach themselves and form the spicules between the two layer. Then the inner mass hollows itself out, and the collar-cells (till now quiescent, with collar and flagellum withdrawn) arrange themselves in a single layer round the cavity, and become active once more. Finally an osculum and pores are developed and the random collection of cells (though by processes not seen in normal development) has become an actual sponge, living and functioning, similar in every way to one that has grown up from the egg.
Of the two experiments, the first is the more surprising, the second the more mysterious. In the first, a new form of life is produced—something capable of living, that is, and yet in its structure unlike any known animal: but, given the large degree of independence possessed by the cells, the rest follows naturally. In the second, however, there seems to be a strange organizing power superior in kind to the powers of the cells themselves—an idea of the whole, informing the parts. Again the image of a general directing his army, even of an architect arranging his materials, springs to the mind: but again, where is the general, where the architect? There is no possibility of anything thus extraneous existing in the normal sponge, still less in the little balls, composed as they are of random cells in random grouping. However, the nature of this directive power we must leave for later consideration (pp. 111–112). Here it suffices to have shown that it exists.
So far the analysis of the simple sponge individual has shown it to be composed of definite, separate cells. These in the normal animal have considerable freedom and independence, both structurally and functionally. Under the artificial conditions of experiment, this independence is shown to be very large, inasmuch as one kind of cell at least can live alone, leading a strange new life, when separated from the rest of the body. Though the whole sponge is a true individual, composed of harmonious parts, yet those parts can themselves behave as harmonious wholes. So far, their independence is merely stated and proved; by their history it can be more or less explained, for various converging testimonies all point to one conclusion, that Sponges are descended from a particular group of Protozoa, and that therefore every cell now forming part of a sponge’s body is derived by an unbroken chain of cell-division (interspersed of course throughout with sexual cell-fusion) from cells which existed as free-living and independent individuals.
On the other hand there does exist a sponge-individuality higher than that of the cells: to start with, in the normal sponge all these cells are working together for a common end, so that every part helps every other part; and in the second place, the plan of this higher individuality somehow permeates all the cells, so that from any group of all the kinds of them taken at random a whole new individual will organize itself.
After this examination of such a compound individual, we must now turn and trace the method by which this second grade of individuality has been built up, the method by which the Metazoa have evolved from Protozoa. The step from first to second grade is one of the two or three most important in the whole history of life; yet it has taken place successfully on several different lines, and unsuccessful attempts are many.
Among the Protozoa, as among almost all other groups of animals, many species live in colonies—using the word colony to mean a collection of organisms all similar to each other, and all united either by living substance or by some framework that the living substance has secreted.
Such colonies are not higher individuals in any sense of the word, but it cannot be denied that they already possess certain properties on which the higher individuality can be grounded. A colony, besides possessing a characteristic shape, forms a single whole, separate from all other similar wholes; this separateness, as has been seen, is a necessary basis for the exclusively or almost exclusively physical individuality of the lower organisms. As regards function, however, the members of the colony often retain as perfect an independence as they would have if living solitary. Colonial life in such species (which are always sedentary), appears to be merely a device for marking the fullest use of a place with good food-supply. Such spots are few and far between, and are discovered by rare individuals only; thus it is of advantage to retain the descendants of these favoured few bound together there in colonies rather than send them off at once into the world with more chances of failure than of success.
In other colonies, function is not so diffuse, and there is a function of the whole which is more than, and sometimes quite different from, the sum of the separate functions of the parts. Even in sedentary species this can sometimes be seen; in Zoothamnium, a colonial bell-animalcule, for instance, a touch on a single one of the animals composing the colony causes the whole colony to retract out of harm’s way. This general contraction, common to a number of individuals, though by no means a necessary result of colonial life, could obviously not occur if the individuals were living separately, however closely they were crowded side by side. But it is in free-swimming colonies that the unity of common function is most pronounced. To take the simplest possible example, imagine two actively-swimming protozoa of the same species joined together by whatever means you please. If free, each would have a similar motion to the other, but both would be independent. When they are joined, however, the motion of the couple is no longer similar to the motion of its two components. Mathematically it is the resultant of their two motions, and as such depends on the way in which the two individuals are attached to each other. If the action of their locomotor organs is not fixed and invariable, it will also depend on the way in which these are used by the two individuals.
Hence for the couple to move, it is essential that the motions of its two parts shall not neutralize each other, but that they shall be co-ordinated to give a resultant motion useful to the whole couple.
Then there is the resistance of the water to be considered, so that before a colonial organism can move effectively its parts will have to acquire a shape, an arrangement, and a mode of action, differing from those which had served them perfectly when they were independent beings.
The further step necessary before the colony can with full right be called an individual is the differentiation of its members so that they perform different functions. As with the primitive homogeneous lump of protoplasm (p. 43) so with the “homogeneous” colony of similar members; both are on the way to acquiring an individuality for themselves, both exhibit features which are the necessary foundations of that individuality, but neither can with justice be said to possess it.
Illustrating these theoretical points, there exist for us, among various other examples, the members of the family Volvocidae,—an old but well-tried object-lesson. These organisms, claimed by botanist and zoologist alike, are members of the Flagellata, unicellular organisms marked off by possessing long whip-lashes or flagella with which they swim. The Volvocidae seem to be a perfectly natural family. They are all free-swimming; they are all colonial, with a framework of transparent jelly common to the colony; they all possess chlorophyll, nourishing themselves after the fashion of plants; and they all have two flagella, a single “eye-spot” and other morphological characters. There can thus be little doubt that they are all descended from a single ancestor who combined these common characters in his person.
The different forms vary very much, however, in the shape and size of the colonies, in the specialization of the sexual elements, and in the degree of individuation of the colonies.
At the base of the series stands Gonium—sixteen precisely similar flagellate cells embedded in firm transparent jelly, joined in definite arrangement to form a flat disc (Fig. 7). The colony thus constituted lives and prospers, nourishes itself, and grows till comes the time for reproduction. Then each cell of the sixteen divides—once, twice, thrice, and four times—into sixteen little ones. Each of the sixteen groups of sixteen breaks away from the rest, arranges its parts in the familiar way, and constitutes itself a minute but perfect new colony.4

Figure 7
Gonium. A, a species containing 16 cells embedded in a flat plate of gelatinous substance. B—F, another species, containing 4 cells. B, C, adult colonies, seen from the top and side respectively. In D one, in E two, and in F all four cells have divided into four. The four groups of four cells in F will shortly separate and become independent daughter-colonies. (Highly magnified.) (From West.)
Among all the other members of the family except Volvox, the asexual reproduction (with which alone we need here be concerned) is accomplished in a similar way—each cell takes upon itself to reproduce a whole new colony. They are colonies and nothing more—their members have united together because of certain benefits resulting from mere aggregation, but are not in any way interdependent, so that the wholes are scarcely more than the sum of their parts.
Though, as we have said, Volvox is obviously related to Gonium and the others, it is separated from them by somewhat of a gap.
In the first place, it contains, instead of sixteen or even sixty-four cells, a vast number, mounting up in some species to twenty thousand (see frontispiece). All these cells are inter-connected by fine strands of protoplasm passing through their party-walls5 and they are arranged in a single layer on the outside of a sphere whose inner parts are filled with a very fluid jelly, so that the Volvox-colony has what we may call an internal medium of its own. Finally, and this is where Volvox has made the great advance, the cells are not all alike. Most are of the type already seen in Gonium and characteristic of the family; these row the colony through the water, steer it, and feed it. Amongst them, in the hinder half of the sphere, are larger cells, lacking flagella and eye-spot, and connected by very numerous strands with their neighbours,
“Their oarsmen-brothers, by whose toil, safe fed
And guarded safe, they live a charmèd life
Within their latticed crystal, peaceably.”
And what do they do in return? Now is discovered the skeleton in the flagellated cells’ cupboard—they cannot reproduce the colony. They are sterile, and must leave reproduction to the big lazy-seeming cells who are only lazy, however, because they must store up food-materials to start the new colony fairly on its way. They grow and grow, bulge inwards, and finally come to float free in the centre space, where they still grow, meanwhile dividing up into a number of cells. In the end, they become perfect miniature colonies, burst out of their parent and swim happily away.
Volvox is thus a real individual; of the two kinds of cells each has given up something the better to fulfil its own special duty. There is division of labour, and, from the point of view of the species, each kind is meaningless apart from the other.
The division of labour in Volvox is that usually first seen in compound individuals—between the reproductive functions on the one side and all the rest on the other. In other words, one sort of cell is concerned entirely with the species, the other entirely with the separate individuals of which the species consists; to use the current phraseology, the one sort is germinal, the other somatic. The word somatic opens up another view: Volvox is the first organism which, in the ordinary sense of the word, has a mortal body. Its substance is not passed on unimpaired from individual to individual, but with each act of generation the major part must die, sacrificed for the greater efficiency of the race.
In Volvox, this body consists of but one sort of cell: in all the organisms usually known as Metazoa there are at least two sorts, if not more. Besides the division of labour between germ and soma, there is developed another in the soma itself, at the first between protective and nutritive cells, the one forming an outer covering round the other, which in its turn surrounds an internal cavity. But even if Volvox only possesses species-individuality, the individuality is none the less real; and the fact that in the family Volvocidae we can positively affirm that the step from an aggregate to a higher individual has actually taken place, is one of the most important in biology.
This, however, is not the only way in which the second grade can or has been reached. It is quite possible that division of labour should set in at the very beginning, and that no such thing as a colony, using the word in its usual sense as a number of equivalent individuals all derived from a single parent and still connected together, should ever have existed.
The best examples of animals with such a history are the Catenata, a small group, all parasites of certain marine worms, discovered by Dogiel (6) only four years ago, and containing but one known genus, Haplozoon. The structure of the most primitive member of the group is simplicity itself (Fig. 9, e). It is a single row of cells, one end fixed to the wall of the worm’s gut, the other sticking out into the gut cavity. The cells, however, are by no means similar among themselves. The first one takes over all the business of attachment, and most of the nutrition. Actively movable, it possesses at its anterior end a piercing spine and a bundle of delicate protoplasmic threads or pseudopodia, which insinuate themselves far up between the cells lining the host’s digestive tube, and serve the double purpose of holding the parasite firm and of sucking up the juices of the neighbouring tissue. From its posterior end this head-cell is continually dividing off new cells, which remain attached to each other in series, up to some seven or eight. The hinder cells of the series gradually become filled with particles of reserve food, analogous to the yolk granules in an egg, and finally lose their connection with the rest, dropping off into the digestive cavity and passing thence to the outer world. Attempts to rear them further have not succeeded, but there can be no doubt that their function is reproductive, designed to spread the race to other hosts.
That is the simplest form: thence to the most complex an interesting series may be traced, through species where a few of the hinder cells divide in such a way that the animal’s posterior end is a plate, not a mere row, of cells, then up to others where this state of things begins much earlier, so that the plate is broadly wedge-shaped, and finally to forms where the hinder cells divide in all three directions of space, and the posterior end is large and club-shaped, several layers in thickness (Fig. 8). In the front half of the body, little openings exist between cell and cell, which serve to pass food-substances down from the “head” into the other cells. When these are full-fed, they close themselves off from their neighbours and prepare themselves for their reproductive destiny.
The ancestry of these curious creatures is almost certainly to be sought in another group of plant-like unicellular flagellates, the Peridineae. These are two forms which serve to bridge the gap—a large one—between the active free Peridineae and the parasitic multicellular Haplozoon.
The first, Gymnodinium pulvisculus (Fig. 9, a), is also a parasite but an external one: it is found attached to the skin of various pelagic creatures by a stalk or bundle of sucking pseudopodia like those of Haplozoon. So it thrives till it is full grown: then, breaking off from its stalk, it divides up into a large number of little cells each of which develops two flagella, takes on the form characteristic of the free-swimming Peridineae, and is off to infest new hosts. Here, it will be seen, the same cell devotes itself at one period to nutrition and at another to dispersal.
In Blastodinium mycetoides (Fig. 9, b), these two functions are carried on by different structural units: the full-fed cell does not break off from the stalk that nourishes it, but divides transversely into two halves which become separated by a membrane. The one that is no longer attached to the stalk at once begins dividing up to form little flagellates, while the other goes on feeding, grows to full size again, and cuts off a second reproductive cell.
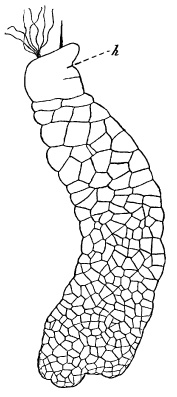
Figure 8
Haplozoon macrostylum, ×300, showing the greatest complexity reached by the Catenata. Only the cell-outlines are drawn. h, head-cell with stylet and pseudopodia. A body-cell is being divided off from it posteriorly. (After Dogiel, slightly modified.)
Now imagine the reproductive cells to remain organically connected with the stalk-cell and to be nourished by it for some time after they have been divided off, and you have in essentials a simple species of Haplozoon (Fig. 9, c–e).
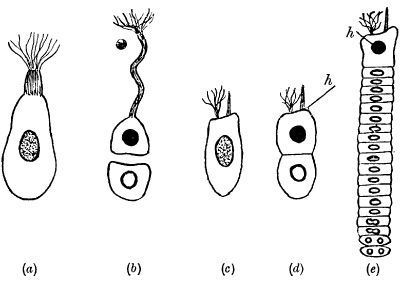
Figure 9
Diagram to show the probable evolution of the Catenata.
(a) Gymnodinium pulvisculus, during its nutritive phase.
(b) Blastodinium mycetoides. A nutritive cell remains permanently attached to the host, and repeatedly divides off reproductive cells from itself.
(c)–(e) Haplozoon lineare. h, head-cell.
(c) One-cell stage, resembling (a).
(d) Two-cell stage, resembling (b) except that the two cells adhere to each other.
(e) Adult, with reproductive cells about to be detached posteriorly.
[Somatic nuclei black; germinal nuclei white; mixed nuclei stippled.] (Modified from Dogiel.)
The Catenata and Volvox are thus similar in being multicellular organisms with unified function and with division of labour among their parts; but their origin is very different.
In the making of Volvox, community-life—mere aggregation—came first, division of labour last. In Haplozoon’s history, division of labour existed before any trace of communal existence, and only later was one cell built up upon another into an individual of a higher order.
To take parallel cases in a different grade of individuality, the simpler Volvocidae closely resemble many low human races among which every family exercises all the ordinary arts and crafts and where society, in spite often of strong communal life, can therefore not rise above the dead level conditioned by the impossibility of doing all things at once and doing them well. Curious and interesting it is that these same peoples if taught, can generally learn and learn quickly and well, many arts and industries before undreamt of among them. The capability was there, but they had not learnt how to use it: only by sacrificing some of their multifarious functions is it humanly possible to advance in the rest, and so to raise society. As with men, so with cells—a jack-of-all-trades cannot advance in any, and the same lesson of sacrifice has to be learnt before the colony can become an individual organism.
A human illustration for the methods of Haplozoon may also be found, or at least imagined. Imagine then a man inflamed with the desire to spread among a benighted race some gospel of good tidings. Poor, he prints the books himself; then comes the question of sending them forth. It is obviously impossible for a single man to do one and the other simultaneously. If he goes out to distribute them himself, the printing will be at a standstill while he is away. If, however, he can obtain volunteers to distribute the books, he himself can stay behind and pull off impressions all the time while a new man goes off with each consignment. Suppose further that while printing he can instruct the distributors in such a way that they will later be able to do their work more soundly, then there will be collected a crowd of embryo distributors at headquarters, from which the fully-trained ones will from time to time depart.
In the first stage the business is like Gymnodinium pulvisculus: then like Blastodinium, and at the last like Haplozoon: the division of labour has come as the first forerunner of the higher development, and this it has done because in both cases there are two special functions to perform which cannot be performed simultaneously by a single individual.
The existence of two mutually exclusive necessities is thus the origin of this type of higher individual: at first the single cell performs them both, but at the expense of not feeding while it is reproducing, not reproducing while it is feeding. Again the sacrifice by a part leads to improvement for the whole; the great fact once discovered that of two cells one can feed for both, the other reproduce for both and the later steps follow almost as a matter of course.
It is to be remarked that for the two functions of nutrition and reproduction thus to clash with one another, it must needs be that the organism can only thrive in a very special and a very limited environment. An individuality like that of the Catenata is therefore found chiefly among parasites, which exist in just such an environment; but in the outer world the conditions are rarely narrow and rigorous enough to call forth such adaptations.6
It was the other method, aggregation of similar units and subsequent division of labour among them, that opened to life the full resources of the second grade of individuality. In some colony like Volvox there once lay hidden the secret of the body and mind of man.