Kreationisterna är djupt förälskade i den fossila dokumentationen, för de har lärt varandra att gång på gång upprepa mantrat att den är full av ”luckor”. ”Fram med era ’mellanformer’!” De inbillar sig med stor förtjusning att dessa ”luckor” är pinsamma för oss evolutionister. I själva verket har vi tur som har några fossil överhuvudtaget, för att inte tala om de väldiga kvantiteter som vi verkligen har att dokumentera evolutionshistorien med – stora mängder som utgör perfekta ”mellanformer”, hur man än definierar dem. I kapitel 9 och 10 understryker jag att vi inte behöver fossil för att bevisa att evolutionen är ett faktum. Beviset för evolutionen skulle vara fullständigt vattentätt även om inte en enda död kropp någonsin hade fossiliserats. Det är en extra fördel att vi faktiskt ha rika fossilådror att gräva ut, och fler upptäcks varje dag. De fossila bevisen för evolution i många stora djurgrupper är underbart starka. Men det finns förstås ändå luckor, och kreationisterna är besatta av dem.
Vi ska än en gång ta upp vår analogi med brottsutredaren som kommer till en brottsplats där inga ögonvittnen finns. Sir Charles uppe på herrgården har blivit skjuten. Fingeravtryck, fotavtryck, DNA från en svettfläck på pistolen och ett starkt motiv: allt pekar på butlern James. Saken är så gott som klar och juryn och alla i domstolen är övertygade om att James gjorde det. Men så hittar man ett bevis i sista minuten, just innan juryn ska dra sig tillbaka och överväga vad som nyss såg ut att vara det ofrånkomliga avgörandet: att James är skyldig. Någon kommer plötsligt ihåg att sir Charles hade installerat övervakningskameror mot inbrottstjuvar. Spänningen är på topp medan filmerna visas. På en ser man James dra ut lådan i serveringsrummet, ta upp en pistol, ladda den och smyga ut med ondskefull blick. Man skulle tro att det bekräftar misstankarna mot James ännu mer. Men James försvarsadvokat påpekar skarpsinnigt att det inte fanns någon kamera i biblioteket där mordet skedde och inte heller någon i korridoren från serveringsrummet. Han viftar med pekfingret på det betvingande sätt som advokaterna har gjort till sitt. ”Det finns en lucka i filmen! Vi vet inte vad som hände när min klient hade lämnat serveringsrummet. Bevisen mot min klient räcker inte för en fällande dom.”
Åklagaren framhåller förgäves att det fanns en andra kamera i biljardrummet, och på den syns James genom den öppna dörren med pistolen skjutfärdig där han smyger på tå genom passagen till biblioteket. Visst täpper detta till luckan i filmen? Visst är det nu alldeles uppenbart att James är skyldig? Men nej. Försvarsadvokaten spelar triumferande ut sitt ess. ”Vi vet inte vad som hände innan eller efter det att min klient passerade den öppna dörren till biljardrummet. Nu finns det två luckor i filmen. Ärade jurymedlemmar, saken är klar. Nu finns det ännu mindre bevis mot min klient än tidigare.”
Liksom övervakningskameran i deckaren är den fossila dokumentationen en extra fördel, något vi inte hade någon rätt att förvänta oss som en självklarhet. Det fanns redan mer än nog med bevis för att döma butlern utan övervakningskameran, och juryn stod just i begrepp att fälla honom innan kameran upptäcktes. På samma sätt finns det mer än nog med bevis för evolutionen i det komparativa studiet av moderna arter (kapitel 10) och deras geografiska fördelning (kapitel 9). Vi behöver inte fossilen – beviset för evolutionen är vattentätt utan dem, så att använda luckor i den fossila dokumentationen som om de motbevisade evolutionen är paradoxalt.
Vad som skulle motbevisa evolutionen, och det mycket övertygande, vore om ett enda fossil påträffades i fel geologiskt lager. När J. B. S. Haldane blev ombedd att nämna en observation som skulle motbevisa evolutionsteorin kom han med det berömda svaret: ”Fossila kaniner i det prekambriska lagret!” Inga sådana kaniner, inga autentiskt anakronistiska fossil av något slag, har någonsin hittats. Alla fossil vi har, och de är överväldigande många, förekommer utan ett enda verifierat undantag i rätt tidsföljd. Inte ett enda enstaka fossil har någonsin påträffats i något lager som bildats innan just den organismen kunde ha utvecklats. Detta är ett mycket avslöjande faktum. Evolutionen kunde så lätt motbevisas om ett enda fossil påträffades i tidsmässigt fel geologiskt lager. Den har bestått detta prov med glans. Evolutionsskeptikerna som vill bevisa sin teori borde ägna all sin tid åt att klättra omkring i bergen och desperat leta efter anakronistiska fossil. De hittar inget, hur de än ligger i.
Den största luckan, och den som kreationisterna tycker allra mest om, är den som föregick den så kallade kambriska explosionen. För lite drygt en halv miljard år sedan, under den kambriska perioden, dyker större delen av phyla – stammarna i det biologiska hierarkiska systemet – ”plötsligt” upp i den fossila dokumentationen. Det vill säga plötsligt i bemärkelsen att inga fossil av dessa djurgrupper är kända i berg äldre än de kambriska, inte plötsligt i bemärkelsen ”på ett ögonblick”. Den period vi talar om täcker omkring 20 miljoner år. Tjugo miljoner år ter sig som en kort tidrymd när de ligger en halv miljard bakåt i tiden, men de utgör naturligtvis exakt samma mängd tid för evolutionen som 20 miljoner år i dag. Hur som helst är det fortfarande plötsligt, och som jag skrev i en tidigare bok har vi ett avsevärt antal stora djurgrupper från den kambriska perioden:
djur som redan befinner sig i ett framskridet utvecklingsskede allra första gången de uppträder. Det är som om de hade planterats där, utan någon evolutionär historia. Detta intryck av att de plötsligt har planterats har kreationisterna självfallet anammat med liv och lust.
Den sista meningen visar att jag hade förstått att kreationisterna skulle gilla den kambriska explosionen. Då jag skrev den, 1986, hade jag inte vett att inse att de skadeglatt skulle citera mina rader mot mig gång på gång som bevis för sin egen tes och nogsamt utelämna mina omsorgsfullt valda förklarande ord. Av en nyck letade jag just på nätet efter ”Det är som om de hade planterats där, utan någon evolutionär historia” och fick inte mindre än 1 250 träffar. Som ett enkelt kontrolltest av hypotesen att de flesta träffarna representerar kreationistiskt citatletande sökte jag för jämförelsens skull efter den mening som följer omedelbart på citatet ovan ur Den blinde urmakaren: ”Evolutionister av alla kulörer är emellertid övertygade om att detta verkligen representerar en mycket stor lucka i den fossila dokumentationen.” Jag fick sammanlagt 63 träffar, jämfört med de 1 250 för den föregående meningen.
Jag har behandlat den kambriska explosionen utförligt, särskilt i Unweaving the Rainbow. Här ska jag bara lägga till en enda ny synpunkt, illustrerad av plattmaskarna, Platyhelminthes. Denna stora maskgrupp omfattar de parasitiska levermaskarna och binnikemaskarna, som har stor medicinsk betydelse. Mina favoriter är ändå virvelmaskarna, turbellaria, det finns mer än fyratusen arter av, ungefär lika många som alla däggdjursarter sammanlagda. Några av dessa virvelmaskar är utomordentligt vackra, något som bilderna visar. De är vanliga, både i vatten och på land, och har antagligen varit vanliga under mycket lång tid. Därför skulle man vänta sig att få se en rik fossilhistoria. Tyvärr finns det nästan ingenting. Frånsett en handfull osäkra spårfossil har inte en enda fossil plattmask någonsin påträffats. Alla exemplar av Platyhelminthes befinner sig redan ”i ett framskridet utvecklingsskede allra första gången de uppträder. Det är som om de hade planterats där, utan någon evolutionär historia”. Men i det här fallet är ”allra första gången de uppträder” inte den kambriska perioden utan i dag.
Här har kreationisterna något att bita i. De tror att plattmaskarna skapades i samma vecka som alla andra varelser och därmed haft exakt lika lång tid på sig att fossiliseras som alla andra djur. Under alla de sekel då alla dessa djur med ben eller skal avsatte sina fossil i tusental måste plattmaskarna ha levt lyckliga sida vid sida med dem, men utan att lämna några signifikanta spår av sin närvaro i berget. Vad är det då som är så märkvärdigt med luckor i registret över de djur som verkligen fossiliseras, med tanke på att plattmaskarnas historia bildar en enda stor lucka, trots att de enligt vad kreationisterna själva säger har levt lika länge? Om luckan före den kambriska explosionen används som bevis för att de flesta djur plötsligt kom till under den kambriska perioden, bör exakt samma ”logik” tillämpas för att bevisa att plattmaskarna plötsligt kom till i går. Men detta motsäger kreationisternas övertygelse att plattmaskarna skapades under samma kreativa vecka som allt annat. Man kan inte både äta kakan och ha den kvar. Detta argument utraderar totalt och i ett enda slag kreationisternas påstående att den prekambriska luckan i den fossila dokumentationen försvagar beviset för evolutionen.
Varför finns det från evolutionär synpunkt så få fossil före den kambriska perioden? Samma faktorer som verkade på plattmaskarna under geologisk tid fram till våra dagar verkade troligen på resten av djurriket före den kambriska perioden. De flesta djur före den kambriska perioden hade förmodligen mjuka kroppar liksom moderna plattmaskar, och de var antagligen också ganska små liksom moderna virvelmaskar. De var helt enkelt inte lämpade för fossilisering. Men så hände något för en halv miljard år sedan som tilllät djur att fossiliseras fritt – till exempel utvecklingen av hårda skelett med mineralinnehåll.



Virvelmaskar – inga fossil finns men de måste ha funnits där hela tiden
En tidigare beteckning för ”lucka i den fossila dokumentationen” var ”den felande länken”. Uttrycket var på modet i det senviktorianska England och dröjde sig kvar in på 1900-talet. Det gick tillbaka på en missuppfattning av Darwins teori och användes som en förolämpning på ungefär samma sätt som ”neandertalare” (orättvist) används i dag. I listan på representativa citat i Oxford English Dictionary förekommer ett från 1930 där D. H. Lawrence berättar om en kvinna som skrev till honom att hans namn ”stank” och fortsatte: ”Ni som är en blandning av den felande länken och schimpansen.”
Den ursprungliga innebörden var oklar, vilket jag ska visa, och antydde att Darwins teori saknade en avgörande länk mellan människan och andra primater. Ett annat av ordbokens illustrativa citat, detta från den viktorianska tiden, använder ordet på följande sätt: ”Jag har hört talas om någon felande länk, mellan människan och aporna.” Än i denna dag älskar historieförnekarna att säga med vad de tror är ett retsamt tonfall: ”Men ni har fortfarande inte hittat den felande länken”, och så kommer de med ett nålstick om Piltdownmannen som påbröd. Ingen vet vem som hittade på lurendrejeriet med Piltdownmannen, men den personen har ett tungt ansvar. Att en av de första kandidaterna till ett människo-apfossil som påträffades var en bluff gav historieförnekarna en ursäkt för att nonchalera de mycket talrika fossil som inte är falska, och de har fortfarande inte slutat gå på om det. Om de bara ville studera fakta skulle de snart se att vi nu har ett rikt förråd av mellanliggande fossil som kopplar moderna människor till den förfader som vi delar med schimpanserna. Det vill säga på den mänskliga sidan om skiljelinjen. Intressant nog har man ännu inte funnit några fossil som knyter denne förfader (som varken var schimpans eller människa) till den moderna schimpansen. Det kanske beror på att schimpanser lever i skogar, som inte ger goda förutsättningar för fossilisering. Om något är det schimpanser, inte människor, som nu för tiden har rätt att klaga över felande länkar!
Detta är alltså en av betydelserna av ”den felande länken”, den påstådda luckan mellan människan och resten av djurriket. Den felande länken i den bemärkelsen felar milt uttryckt inte längre. Jag återkommer till detta i nästa kapitel, som handlar just om mänskliga fossil.
En annan betydelse avser den påstådda bristen på så kallade ”övergångsformer” mellan större grupper, till exempel mellan reptiler och fåglar eller mellan fiskar och amfibier. ”Fram med era mellanformer!” Evolutionisternas svar på denna utmaning från historieförnekarna är att kasta till dem benen från Archaeopteryx, den berömda ”mellanformen” mellan ”reptiler” och fåglar. Det är ett misstag, vilket jag ska visa. Archaeopteryx är inget svar på någon utmaning, för det finns ingen utmaning som är värd att svara på. Att föra fram ett enda berömt fossil som Archaeopteryx underblåser en felaktighet. Man kan på goda grunder hävda i fråga om mängder av fossil att vartenda ett är en mellanform mellan något och något annat. Den förmenta utmaning som ser ut att besvaras av Archaeopteryx bygger på ett föråldrat begrepp, det som förr kallades ”varandets kedja”, och det är under det namnet som jag kommer att behandla den längre fram i det här kapitlet.
Den enfaldigaste av dessa ”felande länk”-utmaningar är de båda följande, eller varianter på dem, som det finns gott om. Den första lyder: ”Om människan kom från apor genom grodor och fiskar, varför innehåller då inte den fossila dokumentationen en ’grodapa’?” Jag har sett en islamisk kreationist stridslystet fråga varför det inte finns några krokoankor. Den andra lyder: ”Jag lovar att jag ska tro på evolutionen när jag ser en apa föda ett människobarn.” Den sista begår samma misstag som alla de andra, plus det extra att tro att stora evolutionära förändringar sker på en gång.
Två av dessa felaktigheter råkade dyka upp intill varandra på den långa lista med kommentarer som följde på en artikel i den engelska tidningen Sunday Times om en TV-dokumentär om Darwin som jag hade presenterat:
Dawkins syn på religionen är orimlig eftersom evolutionen själv inte är något annat än en religion – man måste tro på att vi alla uppstod ur en enda cell … och att en snigel kan bli en apa osv. Haha – det är den mest skrattretande religionen hittills i historien!
Joyce, Warwickshire, Storbritannien
Dawkins borde förklara varför vetenskapen har misslyckats med att hitta de felande länkarna. Tro på ogrundad vetenskap har mer sagokaraktär än tro på Gud.
Bob, Las Vegas, USA
Det här kapitlet tar upp alla dessa besläktade vanföreställningar och börjar med den löjligaste av allesammans, eftersom svaret på den blir en inledning till de andra.
”Varför finns det inte en grodapa bland alla fossil?” Apor härstammar naturligtvis inte från grodor. Ingen evolutionist med förståndet i behåll har någonsin sagt att de gör det, eller att ankor är avkomlingar till krokodiler eller tvärtom. Apor och grodor delar en förfader, som sannerligen inte liknade en groda och inte heller en apa. Den såg kanske ut lite som en salamander, och vi har verkligen salamanderliknande fossil som härrör från rätt tidsperiod. Men det är inte poängen. Varenda en av alla våra miljoner djurarter delar en förfader med alla de andra. Om ens begrepp om evolutionen är så snedvridet att man tror att vi bör vänta oss att hitta en grodapa och en krokoanka, uttalar man sig säkert sarkastiskt också om avsaknaden av en flodhund och en elefimpans. Och varför dra gränsen vid däggdjuren? Varför inte en kängulacka (mellanform mellan känguru och kackerlacka) eller en bläckpard (mellanform mellan bläckfisk och leopard)? Det finns oändligt många djurnamn som man kan sätta ihop på det sättet.* Flodhästar härstammar självklart inte från hundar eller tvärtom. Schimpanser härstammar inte från elefanter eller tvärtom, precis som apor inte härstammar från grodor. Ingen modern art härstammar från någon annan modern art (om vi utelämnar klyvningar som har förekommit helt nyligen). Alldeles som man kan hitta fossil som närmar sig en grodas och en apas gemensamme förfader kan man hitta fossil som närmar sig elefantens och schimpansens gemensamme förfader. Här har vi en som heter Eomaia, som levde under den tidiga kritperioden, för lite mer än 100 miljoner år sedan.
Eomaia liknade inte alls en schimpans och inte alls en elefant. Den påminner lite om en näbbmus och var antagligen ganska lik deras gemensamme förfader, som den var ungefär samtida med, och en stor evolutionär förändring har alltså ägt rum på båda vägarna från en Eomaia-liknande förfader till en elefant och från samma Eomaia-liknande förfader till en schimpans. Men den är inte i någon mening en elefimpans. Om den vore det skulle den också nödvändigtvis vara en hundko, för den gemensamme förfadern till en schimpans och en elefant är också gemensam förfader till en hund och en sjöko. Den skulle också ha varit en jordhäst, för samme förfader är också den gemensamme förfadern till ett jordsvin och en flodhäst. Blotta föreställningen om en hundko (eller en elefimpans eller en jordhäst eller en känguhörning eller en lejonbuff) är djupt o-evolutionär och löjlig. Detsamma gäller en grodapa, och det är en skam att upphovsmannen till detta fåniga och enfaldiga ord, den australiske kringresande predikanten John Mackay, turnerade i brittiska skolor under 2008 och 2009 där han poserade som ”geolog” och lärde oskyldiga barn att den fossila dokumentationen skulle innehålla ”grodapor” om det låg någon sanning i utvecklingsläran.
Eomaia
Ett lika löjeväckande exempel förekommer i den muslimske kreationisten Harun Yahyas jättelika, överdådigt producerade, glassigt illustrerade och dumdrygt okunniga bok Atlas of Creation. Den har uppenbarligen kostat en förmögenhet att producera, och med tanke på det blir det ännu mer förbluffande att den skickades gratis till tiotusentals lärare i naturvetenskap, bland annat till mig. Trots de enorma pengar som ödslats på boken innehåller den fel som har blivit legendariska. För att belysa den falska föreställningen att de flesta gamla fossil är omöjliga att skilja från sina moderna motsvarigheter visar den en havsorm under artistnamnet ål (två djur så olika sinsemellan att de har förts till olika klasser av ryggradsdjur), en sjöstjärna som en ormstjärna (olika klasser av echinodermer, tagghudingar), en sabellid- (annelid)-mask som en liljeformig sjölilja (en tagghuding; detta par kommer inte bara från olika phyla utan från olika underriken, så de kunde knappast ligga längre från varandra även om de försökte, samtidigt som båda är djur) och – allra bäst – en fiskefluga som en nattslända. Utöver dessa pärlor av enögd okunnighet, som lyfter läsarens munterhet till oanade höjder, innehåller boken ett avsnitt om felande länkar. En bild påstås visa att det inte finns någon mellanform mellan en fisk och en sjöstjärna. Författaren kan inte på fullt allvar tro att evolutionisterna väntar sig att träffa på en övergångsform mellan två så olika djur som en sjöstjärna och en fisk. Jag misstänker ofrånkomligen att han känner sin läsekrets alldeles för väl och avsiktligt och cyniskt exploaterar dess okunnighet.
Låt mig än en gång understryka att människor inte härstammar från apor. Vi delar en förfader med apor. Denne gemensamme förfader råkade vara mycket mer lik en apa än en människa, och vi skulle troligen ha kallat den för en apa om vi hade mött den för ungefär 25 miljoner år sedan. Men trots att människan utvecklades från en anfader som vi på goda grunder skulle kalla en apa, föder inget djur en ny art i en handvändning, eller i varje fall inte en som skiljer sig lika mycket från djuret självt som en människa skiljer sig från till exempel en babian, eller ens från en schimpans. Det är inte vad evolutionen handlar om. Evolutionen är en successiv process, som måste vara successiv om den ska kunna förklara något. Väldiga språng på en enda generation – vilket är vad en apa som föder ett människobarn skulle vara – är nästan lika osannolika som en gudomlig skapelse och uteslutna av samma anledning: statistiskt alltför osannolika. Det vore så roligt om de som vägrar att acceptera evolutionen gjorde sig det minsta lilla besvär att lära sig de enklaste elementa i det som de käbblar mot.
Bakom det ogrundade kravet på ”felande länkar” ligger bland annat en medeltida myt som fyllde människors tankevärld ända fram till Darwins tid och fortsatte att sprida förvirring bland dem därefter. Det är myten om varandets kedja, som säger att allt i universum satt på en stege, med Gud överst, sedan ärkeänglarna, så änglar av olika rang, så människor, djur, växter och ner till stenar och andra skapade ting utan liv. Med tanke på att detta går långt tillbaka till en tid när rasismen var en självklarhet behöver jag inte tilllägga att alla människor inte satt på samma trappsteg. Långt därifrån. Och de manliga medlemmarna av respektive släkten höll sig förstås ett snäpp högre än de kvinnliga, precis som det skulle vara. Men det var den förmenta hierarkin i djurriket som hade störst kapacitet att grumla vattnen när evolutionens idé stormade in på scenen. Det föreföll naturligt att tro att ”lägre” djur utvecklades till ”högre” djur. Och i så fall borde vi kunna se ”länkar” mellan dem, hela vägen upp och ner på ”trappan” eller ”stegen”. En stege där flera pinnar saknas är inte pålitlig. Det är denna föreställning om stegen utan pinnar som lurar bakom mycket av skepticismen kring felande länkar. Men hela stegmyten är grundligt feltänkt och strider mot evolutionen. Det ska jag visa här.
Uttrycken ”högre djur” och ”lägre djur” halkar så lätt av tungan att man får en chock när man inser att de ingalunda obehindrat passar in i det evolutionära tänkandet, som man kanske skulle kunna tro. Tvärtom var de – och är – raka motsatsen till det. Vi tror oss veta att schimpanser är högre djur och daggmaskar lägre, vi tror att vi alltid har vetat vad det betyder och vi tror att evolutionen klargör det ännu mer. Men det gör den inte. Det är ingalunda klart att indelningen i ”högre” och ”lägre” betyder något alls. Eller om den betyder något är det så många olika saker att det är missvisande, rentav farligt.
Här följer en lista på förvillande betydelser, mer eller mindre distinkta, till exempel när man säger att en apa står ”högre” än en daggmask.
1. ”Aporna utvecklades ur daggmaskar.” Detta är fel, precis som det är fel att människor har utvecklats ur schimpanser. Apor och daggmaskar har en gemensam förfader.
2. ”Apornas och daggmaskarnas gemensamme förfader var mer lik en daggmask än en apa.” Det kan låta rimligare. Man kan till och med använda ordet ”primitiv” på ett halvprecist sätt om man definierar det som ”bär likhet med förfäder”, och det är självfallet sant att vissa moderna djur är mer primitiva i denna bemärkelse än andra. När man tänker efter betyder det att den mer primitiva av ett par arter har förändrats mindre efter den gemensamme förfaderns tid (alla arter utan undantag har en gemensam förfader om man går tillräckligt långt bakåt). Om ingendera arten har förändrats dramatiskt mer än den andra, bör ordet ”primitiv” inte användas när man jämför dem.
Här lönar det sig att stanna upp och lägga fram ett besläktat resonemang. Det är svårt att mäta grader av likhet. Och hur som helst finns det ingen nödvändig anledning till att den gemensamme förfadern till två moderna djur ska vara mer lik det ena eller det andra. Av två djur, till exempel en sill och en bläckfisk, är det möjligt att det ena är mer likt den gemensamme förfadern än det andra, men därför behöver det inte vara så. Båda har haft exakt lika lång tid på sig att avvika från förfadern, så evolutionistens första tanke är, om något, att inget modernt djur bör vara mer primitivt än något annat. Vi väntar oss gärna att båda ska ha förändrats i samma grad men åt olika håll efter den gemensamme förfaderns tid. Det inträffar långt ifrån alltid (till exempel i fallet med apa och daggmask), och det finns inget tvingande skäl för oss att vänta oss det. Dessutom behöver djurs olika kroppsdelar inte utvecklas i samma takt. Ett djur kan vara primitivt från midjan och nedåt men högt utvecklat från midjan och uppåt. För att uttrycka saken lite mer seriöst kan det ena ha ett mer primitivt nervsystem, det andra ett mer primitivt skelett. Lägg särskilt märke till att ”primitiv” i betydelsen ”bär likhet med förfäder” inte behöver vara detsamma som ”enkel” (det vill säga mindre komplicerad). En hästfot är enklare än en människofot (den har till exempel en enda tå i stället för fem), men människofoten är mer primitiv (den förfader som vi delar med hästarna hade fem tår, som vi, så hästen har förändrats mer). Detta för oss till nästa punkt på vår lista.
3. ”Apor är intelligentare [eller vackrare, har större genom, mer komplicerad kroppsbyggnad och så vidare och så vidare] än daggmaskar.” Sådant zoologiskt snobberi ställer bara till oreda när man försöker tillämpa det vetenskapligt. Jag nämner det enbart därför att det så lätt förväxlas med de andra betydelserna, och det bästa sättet att bringa klarhet i förvirringen är att exponera den. Man kan föreställa sig en lång rad skalor att rangordna djur efter, inte bara de fyra jag har nämnt. Djur som sitter högt upp på en av dessa stegar kanske inte sitter högt upp på en annan. Däggdjur har onekligen större hjärnor än salamandrar, men de har mindre genom än vissa salamandrar.
4. ”Apor är mer lika människor än daggmaskar är.” Detta är oförnekligt för det speciella exemplet apor och daggmaskar. Men än sen då? Varför skulle vi välja människor som den standard vi bedömer andra organismer efter? En indignerad blodigel skulle kunna framhålla att daggmaskar har den stora fördelen att vara mer lika blodiglar än människor är. Trots den traditionella inplaceringen av människor på varandets kedja mellan djur och änglar finns det ingen evolutionär grund för den vanliga idén att evolutionen på något sätt är ”inriktad” mot människan eller att människan är ”evolutionens sista ord”. Det är märkligt hur ofta denna inbilska föreställning tränger sig fram. På dess grövsta nivå möter man den i den överallt förekommande grälsjuka frågan: ”Om schimpanser utvecklades till oss, hur kommer det sig då att det fortfarande finns schimpanser?” Detta har jag redan nämnt, och jag skämtar inte. Jag får den frågan gång på gång på gång, ibland från till synes högutbildade personer.
5. ”Apor [och andra ’högre’ djur] är duktigare på att överleva än daggmaskar [och andra ’lägre’ djur].” Här finns inte det ringaste spår av förnuft, eller ens sanning. Alla levande varelser har överlevt minst in i vår tid. Vissa apor, till exempel den utsökta guldtamarinen, löper risk att dö ut. De är mycket mindre duktiga på att överleva än daggmaskar. Råttor och kackerlackor frodas trots att många människor anser att de står ”lägre” än gorillor och orangutanger, som är farligt nära att dö ut.
Jag hoppas att jag har sagt tillräckligt för att klargöra hur enfaldigt det är att rangordna moderna arter på en stege, som om det vore uppenbart vad man menar med ”högre” och ”lägre”, och för att visa hur fullständigt det strider mot det evolutionära tänkandet. Man kan föreställa sig massor av stegar. Ibland kan det vara rimligt att rangordna djur eller i varje fall några stegar separat, men stegarna är inte väl koordinerade med varandra och ingen av dem har rätt att kallas en ”evolutionär trappa”. Men det farliga arvet efter varandets kedja ger också näring åt utmaningen ”Var är mellanformerna mellan stora djurgrupper?” och ligger bakom den nästan lika vanhedrande tendensen bland evolutionisterna att anta en sådan utmaning med att föra fram speciella fossil, till exempel Archaeopteryx. Ändå pågår något annat, dolt under vanföreställningen om Archaeopteryx, och det är av generell betydelse så jag ska ägna det ett par stycken där jag använder Archaeopteryx som ett speciellt exempel på ett generellt fall.
Zoologerna har av tradition delat in ryggradsdjuren i klasser, större grupper med namn som däggdjur, fåglar, reptiler och amfibier. Vissa zoologer, de så kallade ”kladisterna”,* hävdar att en klass rätteligen består av djur som alla har en gemensam förfader i den klassen och att denne förfader inte har avkomlingar utanför den.** Alla fåglar är avkomlingar till en enda förfader som också skulle ha kallats för fågel och med moderna fåglar delat de avgörande kännetecknen – fjädrar, vingar, en näbb och så vidare. De djur som brukar kallas reptiler är inte en riktig klass i denna bemärkelse. Det beror på att kategorin åtminstone i konventionella taxonomier uttryckligen utesluter fåglar (som bildar en egen klass), men trots det är vissa konventionellt erkända ”reptiler” (till exempel krokodiler och dinosaurier) närmare släkt med fåglar än med andra ”reptiler” (till exempel ödlor och sköldpaddor). Somliga dinosaurier är närmare släkt med fåglar än med andra dinosaurier. I strikt mening skulle vi behöva ta med fåglar om vi skulle göra reptilerna till en äkta naturlig klass. Zoologer med benägenhet för kladism undviker ordet ”reptiler” helt och håller och delar upp dem i archosaurier (krokodiler, dinosaurier och fåglar), lepidosaurier (ormar, ödlor och den sällsynta nyzeeländska Sphenodon) och testudiner (havsoch landsköldpaddor). Zoologer som inte ansluter sig till kladistskolan tycker att ett ord som ”reptil” är bra därför att det gör deskriptiv nytta, även om det artificiellt utesluter fåglarna.
Vad är det då hos fåglarna som föranleder oss att skilja dem från reptilerna? Vad är det för något som tycks rättfärdiga att vi tilldelar fåglarna hedersnamnet ”klass” när de evolutionärt talat bara är en gren inom reptilklassen? Det är det faktum att de reptiler som sluter sig tätt omkring dem, fåglarnas nära grannar på livets träd, råkar vara utdöda, medan fåglarna, ensamma i sitt slag, marscherade vidare. Fåglarnas närmaste släktingar finns allesammans bland de sedan länge utdöda dinosaurierna. Om en lång rad dinosauriesläkter hade överlevt skulle fåglarna inte sticka av. De skulle inte ha fått status som en egen klass av ryggradsdjur och vi skulle inte ställa frågor som ”Var finns de felande länkarna mellan reptiler och fåglar?” Archaeopteryx skulle fortfarande vara ett fint fossil att ha på museet men den skulle inte spela sin nuvarande stjärnroll som standardsvaret på (vad vi nu kan se är) en meningslös utmaning: ”Fram med era mellanformer.” Om korten i utdöendets kortlek hade fallit på ett annat sätt skulle vi ha massor av dinosaurier i farten omkring oss, bland annat några befjädrade, flygande dinosaurier med näbb som kallades fåglar. Och fossiliserade befjädrade dinosaurier upptäcks nu allt oftare, så det börjar bli glasklart att Archeopteryx inte är svaret på frågan om mellanformerna.
Nu ska vi fortsätta till några av evolutionens större övergångar, där ”länkar” har påståtts ”fela”.
Frånsett att skjutas upp i rymden är det svårt att tänka sig ett djärvare eller mer livsförändrande steg än att lämna vattnet och gå upp på torra land. De båda livszonerna skiljer sig åt på så många sätt att det kräver en radikal omställning i nästan alla kroppens delar att röra sig från den ena till den andra. Gälar är bra på att hämta syre ur vatten men praktiskt taget värdelösa i luften, och lungor är värdelösa i vatten. Metoder för att ta sig fram som är snabba, smidiga och effektiva i vatten är riskabelt klumpiga på land och tvärtom. Inte att undra på att ett uttryck som ”fisk på torra land” har blivit ett ordspråk. Och inte att undra på att ”felande länkar” i denna del av den fossila dokumentationen är intressantare än det mesta.
Om man går tillräckligt långt tillbaka levde allting i havet – allt livs blöta, salthaltiga hulda moder. Vid olika punkter i evolutionens historia gick företagsamma individer ur många olika djurgrupper upp på land, ibland slutligen till de torraste öknar, och tog med sig sitt eget privata havsvatten i blod och cellvätskor. Utöver de reptiler, fåglar, däggdjur och insekter som vi ser runt omkring oss finner man skorpioner, mångfotingar och tusenfotingar, spindlar och deras släktingar och minst tre phyla maskar bland de grupper som har lyckats med den långa och mödosamma vandringen ut ur livets vattenfyllda livmoder. Och vi får inte glömma växterna, de enda som åstadkommer användbart kol. Om de inte hade invaderat land dessförinnan skulle ingen av de andra migrationerna kunnat ske.
Lyckligtvis är övergångsformerna under vår utvandring, då fiskarna tog sig upp på land, utsökt dokumenterade i rader av fossilfynd. Detsamma gäller övergångsformerna åt andra hållet mycket senare, när valarnas och dugongernas förfäder övergav sitt mödosamt förvärvade hem på torra land och återvände till haven. I båda fallen finns det gott om länkar som tidigare felade men nu pryder våra museer.
När vi säger att ”fiskar” tog sig upp på land måste vi komma håg att ”fiskar” liksom ”reptiler” inte bildar en naturlig grupp. Fiskar definieras med uteslutningsmetoden. Alla ryggradsdjur (utom däggdjuren) som inte lämnade havet och gick upp på land kallas fiskar. Eftersom ryggradsdjurens tidiga utveckling i sin helhet ägde rum i vatten är det inte förvånande att de flesta överlevande grenar på ryggradsdjurens träd fortfarande håller till i havet. Släktskapsförhållandena skiljer sig kraftigt åt i många fall. Öring och tonfisk är närmare släkt med människan än med hajar. Och lungfisk och kvastfeningar är närmare släkt med människan än med öring och tonfisk (och givetvis hajar). Till och med hajar är närmare släkt med människan än med nejonögon och pirålar, de enda moderna arter som finns kvar av den en gång blomstrande och mångskiftande gruppen rundmunnar, men vi kallar dem allesammans för fiskar. Ryggradsdjur vilkas förfäder aldrig vågade sig upp på land ser allihop ut som ”fiskar”, de simmar som fiskar (till skillnad från delfiner, som simmar genom att böja ryggraden uppåt och nedåt, inte från sida till sida som fiskar) och jag tror att allihop smakar som fisk.
Som vi just såg i exemplet med reptiler och fåglar ser en evolutionist en ”naturlig” grupp djur som en där alla medlemmar är närmare släkt med varandra än de är med alla utanför gruppen. ”Fåglar” är en naturlig grupp eftersom de närmast bakåt har en gemensam förfader som någon icke-fågel inte härstammar från. Alla ”fiskars” närmaste gemensamma förfader delas också av många arter utanför gruppen. Om vi lämnar våra avlägsna släktingar hajarna åt sidan tillhör vi däggdjur en naturlig grupp som omfattar alla moderna fiskar med skelett av ben (i motsats till hajarna som har brosk). Om vi sedan utelämnar fiskarna med benskelett och strålfenor (lax, öring, tonfisk, havsängel, det vill säga ungefär alla fiskar man brukar se som inte är hajar) omfattar den naturliga grupp som vi tillhör samtliga ryggradsdjur som lever på land plus de så kallade lobfeniga fiskarna. Det är i dessa sistnämnda som vi har vårt ursprung, och nu ska vi ägna särskild uppmärksamhet åt dem.
I dag är gruppen lobfeniga fiskar reducerad till lungfiskar och kvastfeningar (”reducerad” som ”fiskar”, men de har expanderat kraftigt på land – vi ryggradsdjur som lever på land är avvikande lungfiskar). Fenorna liknar mer ben än strålfenorna på vanliga fiskar. Old Fourlegs var titeln på en populär bok om kvastfeningar skriven av den sydafrikanske biologen J. L. B. Smith, som fäste världens blickar på dem sedan det första levande exemplaret upptäcktes under dramatiska former 1938 i en sydafrikansk trålares fångst: ”Jag skulle inte ha blivit mer häpen om jag hade sett en dinosaurie komma gående på gatan.” Man hade känt till kvastfeningar tidigare, som fossil, men trott att de dog ut samtidigt med dinosaurierna. Smith beskriver målande det ögonblick när han för första gången såg detta enastående fenomen. Margaret Latimer, som hade upptäckt det, hade kallat honom till sig för att han skulle avge sin bedömning som expert. (Han gav kvastfeningen namnet Latimeria.)
Vi gick raka vägen till museet. Miss Latimer var ute för ögonblicket så vaktmästaren förde in oss i det inre rummet och där var den – Kvastfeningen, ja, Gud! Jag var förberedd, men den första anblicken slog mig som en pust ur en vitglödgad smedja och jag kände mig alldeles skakig och konstig, det pirrade i hela kroppen. Jag stod som förstenad. Ja, det fanns inte skuggan av ett tvivel, fjäll för fjäll, ben för ben, fena för fena var det en äkta kvastfening. Den kunde ha varit en av de där varelserna för 200 miljoner år sedan som hade väckts till liv igen. Jag glömde allt annat och bara stirrade och stirrade, och sedan gick jag fram till den, nästan rädd, och rörde vid den och strök över den, medan min hustru tyst såg på. Miss Latimer kom in och välkomnade oss varmt. Det var först då som jag fick målföret tillbaka, jag har glömt hur orden föll men jag försäkrade dem att det var sant, det var faktiskt sant, det var utan tvekan en kvastfening. Inte ens jag kunde tvivla längre.
Kvastfeningar är närmare släkt med oss än med de flesta fiskar. De har förändrats något sedan vår gemensamme förfaders tid, men inte tillräckligt för att flyttas ut ur den kategori djur som i dagligt tal och av yrkesfiskarna skulle klassificeras som fisk. Men de och lungfiskarna är avgjort närmare släkt med oss än de är med öring, lax, tonfisk och det stora flertalet fiskar. Kvastfeningar och lungfiskar är exempel på ”levande fossil”.
Vi härstammar dock inte från lungfisken eller kvastfeningen. Vi delar förfader med lungfiskarna, och den såg mer ut som en lungfisk än som vi. Men den var inte särskilt lik någondera. Lungfiskar må vara levande fossil men de är ändå inte särskilt lika våra förfäder. När vi söker efter förfäderna måste vi i stället leta riktiga fossil i berg. Och vi är särskilt intresserade av fossil från devonperioden, som täcker övergången mellan vattenlevande fiskar och de första ryggradsdjur som levde på land. Även bland riktiga fossil skulle vi vara alltför optimistiska om vi hoppades finna våra förfäder. Vi kan dock hoppas att hitta släktingar till våra förfäder som är tillräckligt nära för att vi ska få veta ungefär hur de såg ut.
En av de mest kända luckorna i den fossila dokumentationen – så stor att den har fått ett namn, ”Romers lucka” (A. S. Romer var en berömd amerikansk paleontolog) – sträcker sig från omkring 360 miljoner år, i slutet av devonperioden, till omkring 340 miljoner år, i början av karbonperioden, ”de kolförande bergformationerna”. Efter Romers lucka stöter vi på vad som ovedersägligen är amfibier krälande genom träsken, en ymnig utspridning av salamanderliknande djur, några stora som krokodiler, som de hade en ytlig likhet med. Det tycks ha varit en bjässarnas tid, för det fanns trollsländor med en vingbredd så lång som min arm, de största insekter som någonsin har levat.* Med början för omkring 340 miljoner år sedan skulle vi nästan kunna kalla karbonperioden för amfibiernas motsvarighet till dinosauriernas tidsålder. Före den låg emellertid Romers lucka. Och före sin lucka kunde Romer bara urskilja fiskar, fiskar med lobfenor, som levde i vatten. Var fanns mellanformerna och vad förmådde dem att våga sig upp på land?
I Oxford eldades min studentfantasi av den ofantligt lärde Harold Puseys föreläsningar. Trots sitt torra och omständliga sätt hade han förmåga att se bortom museala ben till de levande djur av kött och blod som hade fått dra sig fram i en annan värld.* Hans beskrivning, som han hade hämtat från Romer själv, av vad som drev en del lobfeniga fiskar att utveckla lungor och ben tedde sig fullt rimlig i mina öron, och det gör den fortfarande även om den är mindre på modet bland moderna paleontologer än den var på Romers tid. Romer och Pusey tänkte sig årliga torkperioder då sjöar och gölar och vattendrag torkade upp, och sedan svämmade de över året därpå. Fiskar som levde i vatten kunde dra nytta av en tillfällig förmåga att överleva på land medan de släpade sig från en grund sjö eller göl, som hotades av överhängande uttorkning, till en djupare där de klarade sig till nästa regntid. Enligt denna uppfattning använde våra förfäder det torra landet som en tillfällig bro som hjälpte dem tillbaka ner i vattnet, så som många moderna djur också gör. Det var inte fråga om att de en gång för alla tog sig upp på torra land.
Det var olyckligt att Romer introducerade sin teori med en inledning som syftade till att visa att devonperioden utmärktes av torka. När färskare bevismaterial undergrävde det antagandet såg det ut att undergräva Romers hela teori. Han skulle ha gjort klokare i att utelämna inledningen, som hur som helst tog i för mycket. Som jag hävdar i The Ancestor’s Tale fungerar teorin fortfarande, även om devonperioden var mindre förhärjad av torka än Romer trodde från början.
Låt oss i alla fall återgå till fossilen själva. De sipprar sparsamt genom yngre devon, den period som omedelbart föregick karbonperioden: frestande spår av ”felande länkar”, djur som delvis fyllde luckan mellan de lobfeniga fiskar som förekom så rikligt i devonhaven och amfibierna som längre fram klafsade omkring i karbonperiodens träsk. På luckans fisksida upptäcktes Eusthenopteron år 1881 i en samling fossil från Kanada. Den tycks ha varit en ytjagande fisk och gick troligen aldrig upp på land, trots en del tidiga fantasifulla rekonstruktioner. Den hade ändå flera anatomiska likheter med amfibierna som kom 50 miljoner år senare, bland annat skallbenen, tänderna och framför allt fenorna. De användes troligen till att simma med och inte att gå på, men benen följde det typiska mönstret för en tetrapod (namnet på alla landvertebrater). I den främre extremiteten var ett överarmsben ledat mot två ben, strålbenet och armbågsbenet, som i sin tur var ledade mot många små ben, som vi tetrapoder skulle kalla handledsben, mellanhandsben och fingrar. Och den bakre extremiteten visar ett liknande tetrapodmönster.
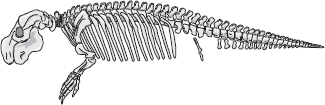
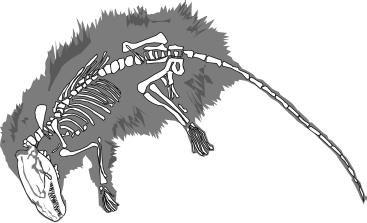
Nära amfibiernas sida av luckan, ungefär 20 miljoner år senare, vid gränsen mellan devon och karbon, levde Ichthyostega. Det blev stor sensation när den påträffades på Grönland 1932. Vi ska inte låta oss missledas av tankar på köld och is. På Ichthyostegas tid låg Grönland vid ekvatorn. Ichthyostega rekonstruerades första gången 1955 av den svenske paleontologen Erik Jarvik, och den bild han tog fram låg närmare ett landlevande djur än moderna experters. Den senaste rekonstruktionen, av Per Ahlberg i Uppsala, Erik Jarviks universitet, placerar Ichthyostega huvudsakligen i vattnet, även om den antagligen gjorde utflykter på land då och då. Ändå liknade den mer en jättesalamander än en fisk, och den hade det platta huvud som är så typiskt för amfibier. Den hade sju tår till skillnad från alla moderna fyrfotingar, som har fem fingrar och tår (åtminstone som embryon, även om det händer att de förlorar några som vuxna). Det verkar som om de första tetrapoderna hade större frihet att ”experimentera” med varierande antal fingrar och tår än vi har i dag. Vid någon punkt bestämde sig de embryonala processerna för fem, och det steg som togs var svårt att vända på. Men inte orimligt svårt. Det finns individuella katter, och för den delen också människor, som har sex tår. Dessa extra tår beror antagligen på ett dupliceringsfel i det embryonala skedet.
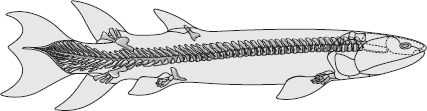
Eusthenopteron
Ichthyostega
En annan spännande upptäckt, också från det tropiska Grönland och också från gränslinjen mellan devon och karbon, var Acanthostega. Den hade också en platt amfibieskalle och tetrapodliknande lemmar, men den avvek ännu mer än Ichthyostega från det vi nu ser som femfingernormen. Den hade åtta tår. De forskare som mer än andra har skänkt oss våra kunskaper om den, Jenny Clack och Michael Coates vid universitetet i Cambridge, tror att Acanthostega liksom Ichthyostega i stort sett levde i vatten, men den hade lungor, och lemmarna tyder starkt på att den kunde klara sig på land likaväl som i vatten om den var tvungen. Återigen såg den ut ungefär som en jättesalamander. Om vi går tillbaka till fiskarnas sida av luckan finner vi Panderichthys, också från yngre devon, som är lite mer amfibielik och lite mindre fisklik än Eusthenopteron. Men om man såg den skulle man säkert kalla den för en fisk och inte för en salamander.
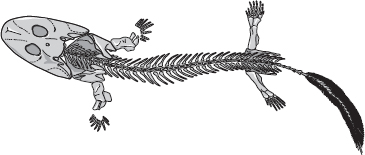
Acanthostega
Panderichthys
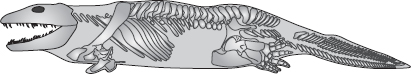
Vi står där alltså med en lucka mellan den amfibieliknande fisken Panderichthys och den fiskliknande amfibien Acanthostega. Var finns den ”felande länken” mellan dem? En expedition från University of Pennsylvania, där Neil Shubin och Edward Daeschler ingick, började leta. Shubin tog deras sökande som underlag för en härlig serie reflektioner om människans utveckling i sin bok Your Inner Fish. Forskarna övervägde noga var de hade störst chanser att finna något och valde med stor omsorg ut en bergig del som hade exakt rätt ålder, yngre devon, i Kanadas arktiska del. Dit for de – och hittade zoologiskt guld. Tiktaalik! Ett namn som aldrig ska glömmas. Det är ett inuitiskt ord för en stor sötvattensfisk. Apropå dess artnamn, roseae, ska jag berätta en varnande historia riktad mot mig själv. När jag först hörde namnet och såg färgfotografierna som det på s. 10 i färgbildarken gick mina tankar omedelbart till devonfärgen, ”den gamla röda sandstenen”, kulören på grevskapet Devon och på Petra i Jordanien. Tyvärr hade jag alldeles fel. Fotot överdriver det rosenröda skimret. Namnet hade valts till ära för en mecenat som bidrog till finansieringen av expeditionen till det arktiska devonområdet. Jag hade den stora förmånen att få en förevisning av Tiktaalik roseae av Edward Daeschler när jag åt lunch tillsammans med honom i Philadelphia kort efter upptäckten, och den livslånge zoologen inom mig – eller kanske min inre fisk – blev mållös av gripenhet.
Om man råkade möta en äkta levande Tiktaalik, nos mot nos, skulle man nog rygga tillbaka som inför en krokodil, för det var så den såg ut. Ett krokodilhuvud på en salamanderkropp, med bakänden och stjärten på en fisk. Till skillnad från alla andra fiskar hade Tiktaalik hals. Den kunde vrida på huvudet. I nästan alla detaljer är Tiktaalik den perfekta felande länken – perfekt därför att den nästan exakt är hälften fisk och hälften amfibie, och perfekt därför att den inte felar längre. Vi har fossilet. Man kan se det, röra vid det, försöka bedöma hur gammalt det är – och misslyckas.
Förflyttningen från vatten till land utlöste en betydande omvandling av varje aspekt av livet, från andning till reproduktion. Det var en väldig vandring genom det biologiska rummet. Med vad som nästan ser ut som överlagd vrånghet vände många etablerade landdjur ändå om längre fram, kastade bort sina mödosamt förvärvade kunskaper om hur man klarar sig på land och vandrade tillbaka ner i vattnet. Sälar och sjölejon har bara tillryggalagt en del av vägen. De visar hur mellanformerna kan ha sett ut, på vägen till extrema fall som valar och dugonger. Valar (även de små valar som vi kallar delfiner) och dugonger med sina nära släktingar sjökorna upphörde helt och hållet att vara landdjur och återgick till sina avlägsna förfäders marina vanor. De går inte ens i land för att para sig. Men de andas fortfarande luft eftersom de aldrig utvecklade något som var likvärdigt med deras äldre marina föregångares gälar. Andra djur som har återvänt från land till vatten, åtminstone delvis, är dammsnäckor, vattenspindlar, vattenbaggar, krokodiler, uttrar, havsormar, vattennäbbmöss, galapagosskarvar som inte kan flyga, Galapagos havsleguaner, yapoker (vattenlevande pungdjur från Sydamerika), näbbdjur, pingviner och sköldpaddor.
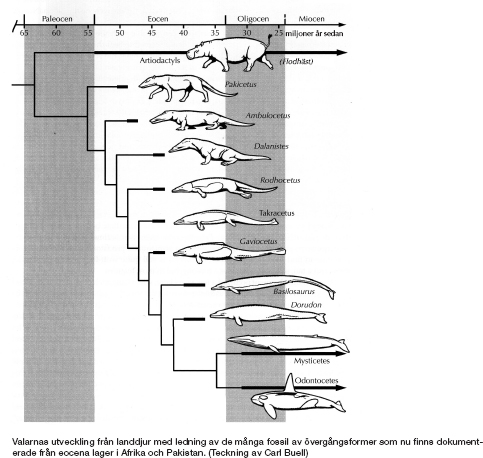
Valarna var länge gåtfulla, men på senare tid har vi fått veta mycket om deras evolution. Molekylärgenetiskt material (se kapitel 10) visar att valarnas närmaste nu levande släktingar är flodhästar, så grisar och sedan idisslare. Ännu mer överraskande visar molekylärbeläggen att flodhästarna är närmare släkt med valar än med djur med klövar (till exempel grisar och idisslare) som är mycket mer lika dem till utseendet. Detta är ännu ett exempel på den diskrepans som ibland uppstår mellan släktskap och fysisk likhet. Vi såg det tidigare i samband med fiskar som är närmare släkt med oss än med andra fiskar. I det fallet uppstod anomalin genom att vår linje lämnade vattnet och gick upp på land och följaktligen rusade iväg i evolutionen, medan våra nära kusiner bland fiskarna, lungfisken och kvastfeningen, behöll sin likhet med mer avlägsna fisksläktingar därför att allesammans stannade kvar i vattnet. Nu stöter vi på samma företeelse igen, men omvänt. Flodhästarna blev åtminstone delvis kvar på land och liknar därför sina mer avlägsna landlevande släktingar idisslarna, medan deras närmare släktingar valarna gav sig ut i havet och förändrades så drastiskt att deras likheter med flodhästarna undgick alla biologer utom molekylärgenetikerna. Liksom när deras avlägsna fiskförfäder från början gick åt andra hållet var det lite som att flyga ut i rymden, eller i varje fall som att släppa upp en ballong, eftersom valarnas förfäder svävade fritt från tyngdlagens belastning och skar av sina förtöjningar med torra land.
Samtidigt har den tidigare ganska torftiga fossila dokumentationen från valarnas evolution kompletterats på ett övertygande sätt, huvudsakligen tack vare en ny skattgömma i Pakistan. Fossilvalarnas historia har emellertid behandlats så väl i andra böcker som har kommit på senaste tiden, till exempel Donald Protheros Evolution: What the Fossils Say and Why it Matters och Jerry Coynes färska Why Evolution is True, att jag har bestämt mig för att inte återge samma detaljer här. I stället begränsar jag mig till ett enda diagram, hämtat ur Protheros bok, som visar en sekvens av fossil ordnade kronologiskt (se nästa sida). Observera hur noga bilden är ritad. Det är frestande – och man brukade se det i äldre böcker – att dra fossilsekvenser med pilar från äldre till yngre. Men ingen vet till exempel om Ambulocetus härstammade från Pakicetus, eller om Basilosaurus härstammade från Rodhocetus. Diagrammet följer den försiktigare linjen och anger möjligheten till exempel att valarna härstammar från en samtida släkting till Ambulocetus som antagligen var ganska lik Ambulocetus (och rentav kan ha varit Ambulocetus). Fossilen i diagrammet är representativa för olika stadier i valens evolution. De bakre extremiteterna som försvinner gradvis, de främre som omvandlas från ben att gå på till fenor att simma med och stjärten som plattas ut till sidoflikar är alla förändringar som skedde i en elegant kaskad.
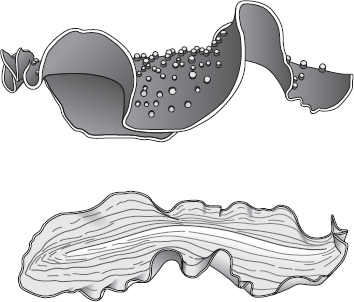
Den andra, mindre talrika och olikartade men lika grundligt vattenlevande gruppen av marina däggdjur, sirendjuren – dugonger och sjökor – är inte så väl etablerad i den fossila dokumentationen, men en utomordentligt vacker ”felande länk” har upptäckts på sistone. Pezosiren, fossilet av ”den vandrande sjökon”, är från Jamaica och ungefär samtida med Ambulocetus, ”den vandrande valen” från den eocena epoken. Den är ganska lik en sjöko eller en dugong så när som på att den har riktiga ben att gå på både fram och bak, där dagens arter har fenor fram och inga bakre extremiteter alls. Bilden på nästa sida föreställer ett modernt dugongskelett (överst), Pezosiren (under).
Fossila valar
Alldeles som valar är besläktade med flodhästen är sirendjuren besläktade med elefanten. Det framgår av en mängd belägg, bland dem det omätligt viktiga molekylära. Men Pezosiren levde antagligen som en flodhäst, tillbringade mycket tid i vatten och använde benen både till att gå på botten med och till att simma. Kraniet är omisskännligt sirenskt. Det är inte klarlagt om Pezosiren verkligen var de moderna sjökornas och dugongernas förfader, men den passar perfekt för rollen.
Modern dugong
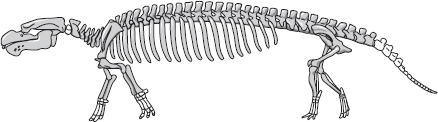
Pezosiren, forntida dugong
Nu till en annan grupp djur som har vänt tillbaka till vatten från land, ett alldeles särskilt spännande exempel därför att en del av dem ändrade sig och tog sig upp på land en gång till! I ett viktigt avseende har havssköldpaddorna behållit sina vanor från livet på land mer än valar eller sirendjur: de lägger fortfarande sina ägg på stränder. I likhet med alla ryggradsdjur som har sökt sig till vattnet igen har sköldpaddorna inte upphört att andas luft, men i den detaljen går några av dem längre än valarna. Dessa sköldpaddor hämtar extra syre ur vattnet genom ett par kammare i stjärtpartiet som är rikligt försedda med blodkärl. En australisk flodsköldpadda får större delen av sitt syre genom ändtarmen.
Det som genast faller i ögonen är sköldpaddornas sköld. Hur utvecklades den och hur såg mellanformerna ut? Var finns de felande länkarna? Vad har de för nytta av en halv sköld (frågar nog en nitisk kreationist)? Märkligt nog har vi just fått en beskrivning av ett nyfunnet fossil som vältaligt besvarar den frågan. Det debuterade i tidskriften Nature i sista ögonblicket innan jag var tvungen att lämna in manuskriptet till den här boken till förlaget. Det var en vattenlevande sköldpadda som påträffades i yngre triassediment i Kina och beräknas ha levt för 220 miljoner år sedan. Den heter Odontochelys semitestacea, ett namn som anger att den till skillnad från moderna sköldpaddor hade tänder och faktiskt en enda sköld. Den hade också en mycket längre stjärt än moderna sköldpaddor. Alla dessa tre detaljer pekar ut den som förstklassigt ”felande länk”-material. Buken var täckt av en sköld, det så kallade plastron, på ungefär samma sätt som på en modern havssköldpadda. Men den saknade nästan helt sköldens ryggdel, carapax. Ryggen var antagligen mjuk, som en ödlas, men längs mitten över ryggbenet hade den hårda, benliknande plattor som på en krokodil, och revbenen var tillplattade som om de ”försökte” bilda den evolutionära begynnelsen till en ryggsköld.
Och här har vi en intressant kontrovers. Författarna till den uppsats som presenterade Odontochelys för världen, Li, Wu, Rieppel, Wang och Zhao (för korthets skull hänvisar jag i fortsättningen till dem som ”de kinesiska författarna” trots att Rieppel inte är kines), anser att deras fynd hade hunnit halvvägs till att skaffa sig en sköld. Andra ifrågasätter dess anspråk på att kunna bevisa att skölden utvecklades i vatten. Nature följer den beundransvärda principen att be andra experter än författarna skriva en kommentar till veckans intressantare artiklar, som publiceras i ett avsnitt med rubriken ”News and Views”. Kommentaren till uppsatsen om Odontochelys är skriven av två kanadensiska biologer, Robert Reisz och Jason Head, och de lägger fram en alternativ tolkning. Hela skölden hade kanske redan utvecklats på land, innan Odontochelys förfäder återvände till vattnet. Och den kanske miste sin sköld efter att ha börjat leva i vatten igen. Reisz och Head påpekar att en del av dagens havssköldpaddor, till exempel den väldiga havslädersköldpaddan, har förlorat eller kraftigt reducerat skölden, så deras teori är fullt trovärdig.
Här måste jag göra en kort utvikning och ta upp frågan vad det är för nytta med en halv sköld, och i synnerhet varför Odontochelys var bepansrad på undersidan men inte på översidan. En förklaring kan ha varit att farorna lurade underifrån, vilket tyder på att dessa varelser för det mesta simmade nära ytan, och för övrigt måste de ju gå upp till ytan för att andas. I dag anfaller hajar ofta underifrån, och hajar bör ha varit ett hot i Odontochelys värld. Det finns ingen anledning att tro att deras jaktvanor var olika på den tiden. Som ett parallellt exempel är en av evolutionens mest överraskande prestationer, det extra paret ögon hos fisken Bathylychnops, antagligen avsedd att upptäcka rovdjursangrepp underifrån. Huvudparet ögon ser åt sidan, som på alla vanliga fiskar. Men vart och ett av dessa båda ögon har ett litet extraöga, komplett med lins och näthinna, instoppat på undersidan. Om Bathylychnops kan göra sig besväret att skaffa sig ett helt extra par ögon, sannolikt för att kunna se upp för attacker underifrån, ter det sig trovärdigt att Odontochelys skaffade sig pansar för att avvärja attacker från samma håll. Att ett plastron är bra att ha säger sig självt. Och om man nu tycker att det väl vore bra med en sköld på ryggen också, bara för att vara på den säkra sidan, är svaret enkelt. Sköldar är tunga och otympliga, de är dyra att bilda och dyra att bära omkring. I evolutionen förekommer alltid kompromisser. För landsköldpaddorna leder kompromissen till tungt, motståndskraftigt pansar både på översidan och på undersidan. För havssköldpaddorna medför kompromissen ett starkt plastron under buken men lättare pansar på ryggen. Och det är en trovärdig hypotes att Odontochelys förde den tendensen lite längre.
Bathylychnops extra öga
Har de kinesiska författarna däremot rätt i att Odontochelys var på väg att utveckla en komplett sköld och att skölden utvecklades i vatten, blir slutsatsen att landsköldpaddorna, som har välutvecklade sköldar, härstammar från vattensköldpaddorna. Som vi ska se är det förmodligen så. Men det är anmärkningsvärt, för det innebär att moderna landsköldpaddor representerar en andra vandring från vatten till land. Ingen har någonsin påstått att valarna eller dugongerna återvände till land efter att ha gett sig ut i vattnet. Den alternativa hypotesen för landsköldpaddorna är att de höll till på land hela tiden och utvecklade skölden oberoende av sina vattenlevande släktingar och parallellt med dem. Detta är ingalunda omöjligt, men vi råkar ha goda skäl att förmoda att havssköldpaddorna verkligen återvände till land för att göra ett nytt försök att bli landsköldpaddor.
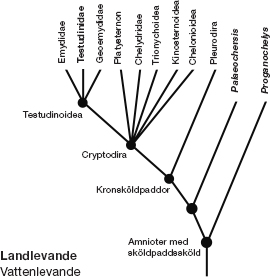
Land- och havssköldpaddors släktträd
Om man ritar upp släktträdet för alla moderna land- och havssköldpaddor med ledning av molekylära och andra jämförelser, är så gott som alla grenarna vattenlevande (mager stil). Landsköldpaddorna anges med halvfet stil, och vi ser att dagens landsköldpaddor utgör en enda gren, Testudinidae, djupt inbäddad i det rika grenverket av i övrigt vattenlevande sköldpaddor. Alla deras nära släktingar är vattenlevande. Moderna landsköldpaddor bildar en enda kvist på busken bestående av vattenlevande sköldpaddor. Deras vattenlevande förfäder begav sig tillbaka upp på land. Detta faktum stämmer med hypotesen att skölden utvecklades i vatten hos en varelse som Odontochelys. Men nu stöter vi på en annan svårighet. Om man studerar släktträdet närmare märker man att det utöver Testudinidae (samtliga moderna landsköldpaddor) finns två fossila släkten av djur med komplett sköld som heter Proganochelys* och Palaeochersis. De återges som landlevande, av orsaker som vi kommer till i nästa stycke. De ligger alldeles bredvid de grenar som representerar vattensköldpaddorna. Det förefaller som om dessa båda släkten långt tillbaka i tiden levde på land.
Innan Odontochelys upptäcktes var dessa båda fossil de äldsta kända exemplaren av Chelonia. Liksom Odontochelys levde de under den yngre triasperioden men ungefär 15 miljoner år senare än Odontochelys. Vissa experter har rekonstruerat dem som sötvattenlevande men forskningsrön från senare tid placerar dem på land. I diagrammet anges deras namn med fetstil. Hur kan man avgöra om fossila djur levde på land eller i vatten, särskilt om man bara har fragment att gå efter? Ibland är det lätt att se. Ichthyosaurierna var reptiler samtida med dinosaurierna, med fenor och strömlinjeformad kropp. Fossilen liknar delfiner och de levde med stor säkerhet som delfiner, i vatten. När det gäller land- och havssköldpaddor är det inte lika lätt att se. Som man kan vänta sig är den mest avslöjande detaljen deras extremiteter. Paddlar skiljer sig ju ganska mycket från ben att gå på. Walter Joyce och Jacques Gauthier vid Yale University tog upp denna förnuftsstyrda iakttagelse och räknade fram de siffror som stödde den. De tog tre viktiga mått i arm- och handbenen på sjuttioen levande arter av Chelonia. Jag ska motstå frestelsen att förklara deras eleganta beräkningar, men deras slutsats var klar. Dessa djur hade ben att gå på, inte paddlar. De levde på land, men de var bara avlägset släkt med moderna landsköldpaddor.
Nu ser vi ut att stå inför ett problem. Om det är riktigt som författarna till uppsatsen om Odontochelys tror, att deras fossil med halv sköld visar att skölden utvecklades i vatten, hur förklarar vi då två släkten landsköldpaddor med komplett sköld 15 miljoner åt senare? Tills Odontochelys upptäcktes skulle jag inte ha tvekat att säga att Proganochelys och Palaeochersis var representativa för den landlevande förfaderstypen innan den återvände till vattnet. Skölden utvecklades på land. Vissa landsköldpaddor med sköldar återvände till havet, liksom sälar, valar och dugonger skulle göra längre fram. Andra stannade på land men dog ut. Och sedan återvände några havssköldpaddor till land och gav upphov till alla moderna landsköldpaddor. Det var vad jag skulle ha sagt, och vad jag också sade i den föregående versionen av det här kapitlet som jag skrev innan Odontochelys steg in på scenen. Men Odontochelys häller tillbaka alla spekulationer i smältdegeln. Nu har vi tre möjligheter, alla lika fascinerande.
1. Proganochelys och Palaeochersis kan vara överlevande från de landlevande djur som tidigare hade skickat en del representanter till havet, däribland förfäderna till Odontochelys. Denna hypotes innebär att skölden utvecklades på land i ett tidigt skede och att Odontochelys förlorade sin carapax i vattnet men behöll bukskölden, plastron.
2. Skölden kan ha utvecklats i vatten, som de kinesiska författarna hävdar, först bukskölden, plastron, och sedan carapax, ryggskölden. Om det var så, hur ska vi då förklara Proganochelys och Palaeochersis, som levde på land efter det att Odontochelys hade levt i vatten med sin halva sköld? Proganochelys och Palaeochersis kan ha utvecklat skölden var för sig. Men det finns en annan möjlighet:
3. Proganochelys och Palaeochersis kan representera ett tidigare, upprepat återvändande från vatten till land. Den tanken tycker jag är fantastiskt spännande.
Vi är redan säkra på att havssköldpaddorna märkligt nog genomförde en evolutionär kretsgång tillbaka till land: en tidig typ av landlevande ”sköldpaddor” återvände till sina ännu tidigare fiskförfäders vatten, blev havssköldpaddor och gick sedan upp på land igen som en ny inkarnation av landsköldpaddor, Testudinidae. Det vet vi, eller är så gott som helt säkra på. Men nu står vi inför tillägget att denna kretsgång inträffade två gånger! Inte bara för att ge upphov till de moderna landsköldpaddorna utan mycket längre tillbaka i tiden, för att ge upphov till Proganochelys och Palaeochersis under triasperioden.
I en annan bok kallar jag DNA ”de dödas genetiska bok”. På grund av det naturliga urvalets sätt att fungera är ett djurs DNA i viss mening en textuell beskrivning av de världar där dess förfäder valdes ut naturligt. I fråga om en fisk beskriver de dödas genetiska bok dess förfäders hav. I fråga om oss själva och de flesta däggdjur utspelar sig de första kapitlen i boken allesammans i havet och de senare enbart på land. Rörande valar, dugonger, havsleguaner, pingviner, sälar, sjölejon, havssköldpaddor och märkligt nog landsköldpaddor innehåller boken en tredje serie kapitel som skildrar deras episka återkomst till sitt avlägsna förflutnas försöksområde, havet. Rörande landsköldpaddorna innehåller boken ett fjärde avsnitt som ägnas åt ett slutgiltigt – eller är det verkligen slutgiltigt? – återvändande till land, kanske två gånger oberoende av varandra vid två vitt skilda tidpunkter. Kan det finnas något annat djur för vilket de dödas genetiska bok är en sådan palimpsest av multipla evolutionära u-svängar? Jag kan inte låta bli att undra över de former som lever i sötvatten och bräckt vatten (sumpsköldpaddor, ”terrapiner”), som är nära släkt med landsköldpaddorna. Flyttade deras förfäder raka vägen från havet in i bräckt och sedan sött vatten? Representerar de ett mellanstadium på vägen från havet tillbaka till land? Eller är det tänkbart att de står för en andra kretsgång till vattnet från förfäder som var moderna landsköldpaddor? Har medlemmarna av ordningen Chelonia gått i skytteltrafik fram och tillbaka i evolutionär tid mellan vatten och land? Kan palimpsesten vara ännu mer intrikat än jag har antytt hittills?
PS
Den 19 maj 2009, då jag läste korrektur på denna bok, tillkännagav den vetenskapliga nättidskriften PLOS One att man hade påträffat en ”felande länk”. Den heter Darwinius masillae och levde för 47 miljoner år sedan i regnskogar i vad som nu är Tyskland. Författarna hävdar att det är det mest kompletta primatfossil som någonsin hittats, med inte bara benen bevarade utan också hud, hår, några inre organ och dess sista måltid. Visst är Darwinius masillae utsökt, men den är omvärvd av överord som fördunklar klara tankeprocesser. Enligt Sky News är den ”världens åttonde underverk” som ”slutgiltigt förklarar Charles Darwins evolutionsteori”. Kära nån då! Den mer eller mindre nonsensartade mystiken kring den ”felande länken” verkar inte ha förlorat sin makt.
* Jag använder ordet ”oändligt” i den vanliga, ofta missbrukade retoriska bemärkelsen ”kolossalt stor”. Det verkliga antalet är antalet parvisa kombinationer av alla arter med alla andra, och det är så nära oändligheten att det inte spelar någon roll.
* Av ordet ”clade”, en grupp organismer som tros omfatta alla en gemensam förfaders evolutionära avkomlingar.
** I varje fall enligt den samstämmiga uppfattningen bland zoologerna, och jag kommer att fortsätta att för resonemangets skull använda fåglarna som ett exempel på en god klass. På senare tid har fossilforskningen visat upp ett antal befjädrade dinosaurier, och någon stiger kanske fram och påstår att en del av de moderna djur vi kallar fåglar härstammar från en annan grupp av befjädrade dinosaurier än andra fåglar. Om alla moderna fåglars närmaste gemensamme förfader visar sig vara ett djur som inte skulle klassificeras som en fågel, skulle jag få revidera mitt uttalande att fåglarna utgör en god klass.
* En hypotes går ut på att denna gigantism blev möjlig tack vare att atmosfären innehöll mer syre på den tiden. Insekter saknar lungor och andas genom små luftrör som för luft genom hela kroppen. Luftrör kan inte bygga upp ett så omfattande och intrikat distributionssystem som blodkärl, och det verkar sannolikt att detta begränsar kroppsstorleken. Den gränsen bör ha varit högre i en atmosfär med 35 procent syre, i stället för de 21 procent som vi andas in i dag. Detta ger en tillfredsställande förklaring till de jättelika trollsländorna, men den behöver inte vara den rätta. Jag är för övrigt förbryllad över att det inte började brinna överallt med så mycket syre i luften. Det kanske hände. Skogsbränder måste ha varit vanligare än nu för tiden, och fossilen tyder på en hög frekvens eldbeständiga växtarter. Det är inte klart varför atmosfärens syrehalt nådde en topp under karbon- och permperioderna. Det kan ha att göra med att så mycket kol avsattes under jord.
* Han var en Oxfordlärare av den gamla skolan som ansåg att han var där för att undervisa studenter, och han skulle inte ha överlevt i dagens kultur där all forskning ska granskas och utvärderas. Han hade knappt publicerat en enda artikel, och arvet efter honom vilar hos de generationer av tacksamma lärjungar som han delgav sin visdom och åtminstone lite av sin enorma lärdom.
* Man har sagt mig att namnet inte ger någon mening på grekiska. Om det stavades Progonochelys skulle det ge fullgod mening, ungefär ”ursköldpadda”, och jag tror faktiskt att det var vad de ursprungliga författarna avsåg när de namngav släktet. Tyvärr är den zoologiska nomenklaturens regler strikta, och inte ens uppenbara felaktigheter kan rättas till när de väl har befästs i en publicerad namnlista. Taxonomin vimlar av sådana fossiliserade felaktigheter.