Darwin behandlar människans utveckling i sitt mest berömda verk, Om arternas uppkomst, i tio innehållsdigra ord: ”Ljus kommer att kastas över människans ursprung och hennes historia.” Så formulerade han sig i den första upplagan, som är den jag alltid citerar såvida inget annat anges. I den sjätte (och sista) upplagan tillät Darwin sig att gå en liten smula längre, och meningen ändrades till: ”Mycket ljus kommer att kastas över människans ursprung och hennes historia.” Jag tycker mig se den store mannen som sitter med pennan redo över den femte upplagan och noga begrundar om han ska unna sig lyxen av ”Mycket”. Även med det tillfogat är satsens utformning avsiktligt tillbakahållen.
Darwin dröjde avsiktligt med att behandla människans utveckling till en annan bok, Om människans härkomst och könsurvalet. Det är inte förvånande att detta senare verks båda volymer ägnar mer utrymme åt ämnet det sexuella urvalet (som han undersökte huvudsakligen hos fåglar), än åt människans utveckling. Då Darwin skrev sina böcker fanns inga fossil alls som kopplade oss till våra närmaste släktingar bland aporna. Han hade bara levande apor att studera, och han använde dem väl och hävdade alldeles riktigt (och så gott som ensam) att våra närmaste levande släktingar alla var afrikaner (gorillor och schimpanser – på den tiden var bonoboapan erkänd som separat från schimpansen, men den är också afrikan). Därför förutsade han att Afrika var stället att leta på, om man någonsin skulle hitta fossil av människans förfäder. Darwin beklagade bristen på fossil men upprätthöll en orubbligt optimistisk inställning till hela frågan. Han hänvisade till sin mentor Lyell, tidens store geolog, och påpekade att ”upptäckten av fossila lämningar av samtliga vertebratklasser har varit en ytterligt långsam och slumpartad process” och tillade: ”Man får inte heller glömma att de regioner där sannolikheten är störst för lämningar som knyter människan till någon utdöd apliknande varelse ännu inte har undersökts av geologerna”. Han menade Afrika, och sökandet underlättades inte av att hans närmaste efterföljare i stort sett nonchalerade hans råd och letade i Asien i stället.
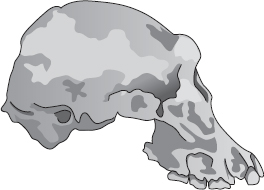
Det var verkligen i Asien som de ”felande länkarna” började bli mindre felande. Men de första fossilen som påträffades var tämligen unga, mindre än en miljon år, och härrörde från en tid då hominiderna hade kommit den moderna människan ganska nära, lämnat Afrika och nått Fjärran östern. De kallades ”javamänniskan” och ”pekingmänniskan” efter de platser där de hittades. Javamänniskan upptäcktes 1891 av den nederländske antropologen Eugene Dubois. Han gav den namnet Pithecanthropus erectus och markerade på så sätt sin övertygelse att han hade förverkligat sitt livs stora ambition och funnit ”den felande länken”. Protester kom från två motsatta håll, vilket bekräftade hans resonemang: somliga sade att hans fossil var rent mänskligt, andra att det var en stor gibbonapa. Längre fram i sitt förgrämda och kverulantiska liv avvisade Dubois hypotesen att de senare upptäckta pekingfossilen liknade hans javamänniska. Han slog järnhårt vakt om sitt fossil och var förvissad om att endast javamänniskan var den autentiska felande länken. För att understryka skillnaden mot de olika pekingfossilen beskrev han dem som långt närmare den moderna människan och sin egen javamänniska från Trinil som en mellanform mellan människa och apa.
Pithecanthropus [javamänniskan] var inte människa utan företrädare för ett gigantiskt släkte knutet till gibbonaporna, men överlägsen dessa tack vare sin utomordentligt stora hjärnvolym och samtidigt utmärkt av sin förmåga att stå och gå upprätt. Den hade antropoidapornas dubbla cefalisation [förhållandet mellan hjärnstorlek och kroppsstorlek] och halva människans. …
Det var hjärnans överraskande volym – som är mycket för stor för en antropoidapa och liten jämfört med den genomsnittliga mänskliga hjärnan, dock inte mindre än den minsta – som ledde till den nu nästan allmänna uppfattningen att ”Apmänniskan” från Trinil på Java egentligen var en primitiv människa. Morfologiskt är emellertid calvaria [hjärnskålen] mycket lik antropoidapornas, särskilt gibbonapans. …
Dubois kan inte ha blivit på bättre humör av att andra tolkade hans ord som att Pithecanthropus bara var en stor gibbonapa, inte alls någon mellanform mellan dem och människan, och han gjorde sig stor möda med att inskärpa sin tidigare ståndpunkt: ”Jag hävdar alltjämt, nu mer bestämt än någonsin, att Pithecanthropus från Trinil är den verkliga ’felande länken’.”
Då och då har kreationisterna använt uppgiften om att Dubois tog avstånd från sitt påstående att Pithecanthropus var en mellanform mellan apa och människa som ett politiskt vapen. Kreationistorganisationen Answers in Genesis (”Svar i Första Moseboken”) har emellertid lagt till det till sin lista på tvivelaktiga argument som de nu säger inte ska anföras längre. Det hedrar dem att de håller en sådan lista överhuvudtaget. Som jag nämnde har både java- och pekingfynden av Pithecanthropus befunnits vara riktigt unga, mindre än en miljon år. De klassificeras nu vid sidan av oss själva i släktet Homo som behåller Dubois specifika namn erectus: Homo erectus.
Dubois valde fel del av världen för sin envisa jakt på den ”felande länken”. Det var naturligt för en nederländare att först bege sig till Nederländska Ostindien, men en så hängiven forskare som han borde ha följt Darwins råd och fortsatt till Afrika, för det var där våra förfäder utvecklades, som vi ska se. Vad hade då dessa exemplar av Homo erectus för sig utanför Afrika? Två utvandringar ägde rum och de får inte förväxlas. Ganska nyligen, för kanske mindre än 100 000 år sedan, lämnade kringresande skaror av Homo sapiens som såg ut ungefär som vi Afrika och delade upp sig i alla de folkslag vi ser runtom i världen i dag: inuiter, ”indianer”, aboriginer, kineser och så vidare. Det är denna utvandring som är mest känd. Erectus-pionjärerna i den tidigare utvandringen lämnade efter sig fossil i Asien och Europa, bland andra på Java och i Peking. Det äldsta kända fossilet utanför Afrika påträffades i Georgien och fick namnet ”georgienmänniskan”: en diminutiv varelse vars (tämligen välbevarade) skalle med moderna metoder har daterats till omkring 1,8 miljoner år. Vissa taxonomer kallar den Homo georgicus (andra erkänner den inte som en separat art) för att ange att den verkar mer primitiv än resten av de första utvandrarna från Afrika, alla klassificerade som Homo erectus. Vissa stenverktyg som är lite äldre än georgienmänniskan har just upptäckts i Malaysia, och det har gett upphov till nytt sökande efter fossila ben på halvön. Hur som helst står alla dessa tidiga asiatiska fossil ganska nära den moderna människan och alla är i dag klassificerade i släktet Homo. För att möta våra tidigare förfäder måste vi bege oss till Afrika, men först ska vi göra en paus och ta reda på vad vi bör vänta oss av en ”felande länk”.
Homo georgicus
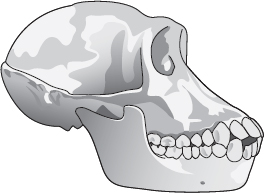
Schimpans
För resonemangets skull ska vi ta den ursprungliga, förvillande betydelsen av uttrycket ”felande länk” på allvar och söka efter en mellanform mellan schimpansen och oss själva. Vi är inte avkomlingar till schimpansen, men det är tämligen säkert att den förfader som vi delar med den mer liknade en schimpans än oss. I synnerhet hade den inte vår enorma hjärna, den gick antagligen inte på två ben som vi gör, den var troligen mycket mer luden än vi är och den saknade med säkerhet sådana avancerade mänskliga företeelser som språk.
Hår och språk lämnar ju inga fossil efter sig, men vi kan få goda ledtrådar till hjärnstorleken av kraniet och goda ledtrådar till gångarten från hela skelettet (inklusive kraniet, för foramen magnum, hålet för ryggmärgen, pekar nedåt hos tvåfotingar, mer bakåt hos fyrfotingar). Tänkbara mellanformer kan ha haft något av följande attribut:
1. Hjärna av mellanstorlek och inte fullt upprätt gång; kanske ett slags lufsande med kutande rygg, inte den stolt rakryggade hållning som underbefäl visar upp på exercisplatsen och danslärarinnor försöker lära ut.
2. Hjärna av schimpansstorlek och upprätt gång.
3. Stor, mer människolik hjärna, gång på alla fyra som en schimpans.
Med dessa möjligheter i minnet ska vi studera några av de många afrikanska fossil som vi nu har tillgång till men som tyvärr inte var påträffade på Darwins tid.
Det finns molekylära belägg (som jag återkommer till i kapitel 10) för att den förfader vi har gemensam med schimpanserna levde för ungefär sex miljoner år sedan eller lite tidigare, så låt oss landa någonstans mittemellan och ta en titt på några fossil som är tre miljoner år gamla. Det mest berömda av den åldern är ”Lucy”, som klassificerades som Australopithecus afarensis av sin upptäckare i Etiopien, Donald Johanson. Tyvärr har vi bara fragment av Lucys kranium, men underkäken är ovanligt väl bevarad. Hon var liten efter moderna mått, dock inte så liten som Homo floresiensis, den diminutiva varelse som tidningarna irriterande nog har gett namnet ”Hobbiten” och som för retsamt kort tid sedan dog ut på ön Flores i Indonesien. Lucys skelett är däremot så komplett att man kan sluta sig till att hon gick upprätt på marken men antagligen också tog sin tillflykt till träd, där hon klättrade vigt. Mycket tyder på att de ben som tillskrivs Lucy verkligen har tillhört en enda individ. Så var det inte med den så kallade ”första familjen”, en samling ben från minst tretton individer som liknade Lucy och levde vid ungefär samma tid. Av någon anledning kom de att begravas tillsammans, också i Etiopien. Fragmenten från Lucy och från första familjen ger ett gott begrepp om hur Australopithecus afarensis såg ut, men det är svårt att göra en autentiskt fullständig rekonstruktion av delar från många olika individer. Lyckligtvis upptäcktes en tämligen komplett skalle, AL 444-2, år 1992 i samma område i Etiopien, och den bekräftade de preliminära rekonstruktioner man hade gjort tidigare.
AL 444-2
Slutsatsen av studier av Lucy och hennes släktingar är att de hade en hjärna av ungefär samma storlek som schimpansernas, men till skillnad från dem gick de upprätt på bakbenen, så som vi gör – det andra av våra tre hypotetiska scenarier. De såg ut ungefär som schimpanser på två ben. Det bekräftas dramatiskt av de rörande fotavtryck som Mary Leakey träffade på i fossiliserad vulkanisk aska längre söderut, i Laetoli i Tanzania. De är äldre än Lucy och AL 444-2, ungefär 3,6 miljoner år. De brukar tillskrivas ett par Australopithecus afarensis som gick sida vid sida (hand i hand?), men det viktiga är att en apa gick upprätt på jorden för 3,6 miljoner år sedan, på två fötter som var ungefär som våra trots att dess hjärna hade samma storlek som schimpansens.
Det förefaller sannolikt att den art vi kallar Australopithecus afarensis – Lucys art – omfattade våra förfäder tre miljoner år bakåt i tiden. Andra fossil har placerats i olika arter av samma släkte, och det är praktiskt taget säkert att våra förfäder var medlemmar av det släktet. Den första australopithecin som påträffades, och typexemplaret av släktet, var det så kallade Taungbarnet. Vid tre och ett halvt års ålder anfölls Taungbarnet av en örn. Beviset utgörs av skadorna på fossilets ögonhålor, som är identiska med skador som moderna örnar åstadkommer på moderna apor när de hackar ut deras ögon. Stackars lilla Taungbarn, som skrek förtvivlat i vinden när du lyftes upp i örnens klor, du skulle inte ha hämtat någon tröst i din förutbestämda berömmelse två och en halv miljon år senare som typexemplaret av Australopithecus africanus. Stackars Taungmamma som grät under pliocenepoken.
Typexemplaret är den första individ av en ny art som får ett namn och officiellt får den första etiketten i ett museum. I teorin kan man jämföra senare fynd med typexemplaret för att se efter om de stämmer sinsemellan. Det var den sydafrikanske antropologen Raymond Dart som 1924 upptäckte Taungbarnet och gav det splitter nya namn på släkte och art.
Vad är det för skillnad mellan ”art” och ”släkte”? Vi ska snabbt reda ut saken så att vi kan gå vidare. Släkte är beteckningen på den större gruppen. En art hör hemma inom ett släkte och delar ofta släkte med andra arter. Homo sapiens och Homo erectus är två arter i släktet Homo. Australopithecus africanus och Australopithecus afarensis är två arter i släktet Australopithecus. Det latinska (eller grekiska) namnet på ett djur eller en växt består alltid av namnet på släktet (med första bokstaven versal) följt av ett artnamn (utan versal). Båda namnen anges med kursiv stil. Ibland förekommer ett extra underartnamn, till exempel Homo sapiens neanderthalensis. Taxonomerna tvistar ofta om namn. Många föredrar Homo neanderthalensis framför Homo sapiens neanderthalensis och upphöjer därmed neandertalaren från underartens till artens status. Namnen på släkten och arter ifrågasätts också ofta och ändras inte sällan i takt med successiva revideringar i den vetenskapliga litteraturen. Paranthropus boisei har hetat Zinjanthropus boisei och Australopithecus boisei* och i dagligt tal hör man beteckningen ”kraftig australopithecin”, i motsats till de båda ”gracila” arterna av Australopithecus som nämns ovan. Ett av huvudämnena för detta kapitel är den zoologiska klassificeringens lätt godtyckliga karaktär.
Raymond Dart gav Taungbarnet, släktets typexemplar, namnet Australopithecus, och ända sedan dess dras vi med detta trist fantasilösa namn på vår förfader. Det betyder ”apa som lever i södern” och ingenting annat. Det har ingenting att göra med Australien, som bara betyder ”sydligt beläget land”. Man kan tycka att Dart borde ha hittat på något fantasifullare namn på ett så viktigt släkte. Han kunde till och med ha gissat att andra medlemmar av släktet skulle upptäckas längre fram norr om ekvatorn.
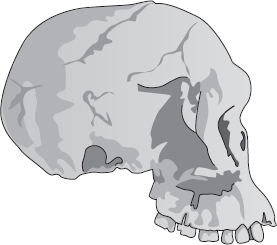
Ett av de allra bäst bevarade kranierna vi har, trots att det saknar underkäke, kallas Mrs Ples och är lite äldre än Taungbarnet. Mrs Ples, som i själva verket kan ha varit en småväxt individ av manligt kön och inte en storväxt individ av kvinnligt kön, fick sitt namn därför att ”hon” från början fördes till släktet Plesianthropus. Ordet betyder ”nästan mänsklig”, vilket är ett bättre namn än ”apa som lever i södern”. När senare tiders taxonomer avgjorde att Mrs Ples och hennes kamrater tillhörde samma släkte som Taungbarnet, borde Plesianthropus ha blivit beteckningen på dem alla. Tyvärr är den zoologiska nomenklaturens regler stränga intill pedanteri. Förnuft och lämplighet får stå åt sidan för ett tvivelaktigt, tidigare knäsatt namn. ”Apa som lever i södern” är ett dumt namn men det spelar ingen roll: det är äldre än det mycket lämpligare Plesianthropus och vi tycks få leva med det, såvida inte … Jag hoppas fortfarande, fast det ser mörkt ut, att någon en dag drar ut en dammig låda i ett sydafrikanskt museum och hittar ett länge bortglömt fossil, helt klart av samma slag som Mrs Ples och Taungbarnet men med en etikett där det står ”Typexemplar av Hemianthropus, 1920”. Alla museer i världen skulle genast få märka om sina exemplar och avgjutningar av Australopithecus och alla böcker och uppsatser om hominidernas förhistoria skulle få följa efter. Ordbehandlingsprogram världen runt skulle arbeta övertid med att nosa reda på Australopithecus och ersätta det med Hemianthropus. Jag kan inte komma på något annat fall där de internationella reglerna är så kraftfulla att de dikterar en världsomfattande och antedaterad språklig förändring över en natt.
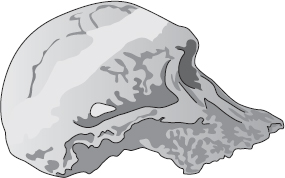
Mrs Ples
Nu kommer vi till mitt nästa viktiga resonemang om godtyckliga namn och påstått felande länkar. När Mrs Ples namn byttes från Plesianthropus till Australopithecus förändrades naturligtvis ingenting i den verkliga världen. Man får förmoda att ingen är frestad att tro något annat. Men vi ska ta upp ett liknande fall där ett fossil underkastas en ny granskning och av anatomiska skäl förs från ett befintligt släkte till ett annat. Eller där rivaliserande antropologer ifrågasätter dess släkttillhörighet, vilket händer mycket ofta. Det ligger ju i evolutionens själva logik att det måste ha funnits individer som satt exakt på gränsen mellan två släkten, till exempel Australopithecus och Homo. Om man tar en titt på Mrs Ples kranium och på ett som tillhör en modern Homo sapiens är det ingen konst att avgöra att de hör hemma i olika släkten. Om vi, som nästan alla antropologer i dag gör, antar att samtliga medlemmar av släktet Homo härstammar från det släkte vi kallar Australopithecus, följer av detta att åtminstone en individ måste ha suttit exakt på gränslinjen någonstans i kedjan från en art till en annan. Detta är en viktig punkt, så jag ska dröja vid den en stund till.
Vi har sagt att formen på Mrs Ples kranium är representativ för Australopithecus africanus för 2,6 miljoner år sedan. Det ska vi hålla i minnet när vi nu studerar skallen KNM ER 1813. Sedan går vi till nästa, KNM ER 1470. Båda är åldersbestämda till omkring 1,9 miljoner år och de flesta auktoriteter placerar båda i släktet Homo. I dag klassificeras 1813 som Homo habilis, men så har det inte alltid varit. Fram till nyligen var 1470 också förd till samma klass, men nu är en rörelse på gång att omklassificera den som Homo rudolfensis. Här ser vi alltså än en gång hur obeständiga våra namn är. Men det spelar ingen roll; båda förs av samstämmiga forskare till släktet Homo. Den uppenbara olikheten ligger i att Mrs Ples och hennes sort hade ett mer framskjutande ansikte och mindre skalle. I båda avseendena förefaller 1813 och 1470 mer mänskliga, Mrs Ples mer ”apliknande”.
Studera nu skallen som kallas Twiggy. Nu för tiden klassificeras också Twiggy i regel som Homo habilis. Men hennes framåtpekande munparti påminner mer om Mrs Ples än om 1470 eller 1813. Några antropologer har placerat Twiggy i släktet Australopithecus, andra i Homo. Vart och ett av dessa tre fossil har vid olika tidpunkter klassificerats som Homo habilis och som Australopithecus habilis. Som jag redan har nämnt har det hänt att vissa auktoriteter gett 1470 ett annat artnamn och bytt ut habilis mot rudolfensis. Och till råga på allt har artnamnet rudolfensis hängts på båda släktnamnen, Australopithecus och Homo. För att sammanfatta har dessa tre fossil av olika auktoriteter vid olika tidpunkter getts följande namn:
| KNM ER 1813: | Australopithecus habilis, Homo habilis |
| KNM ER 1470: | Australopithecus habilis, Homo habilis, Australopithecus rudolfensis, Homo rudolfensis |
| OH 24 (Twiggy): | Australopithecus habilis, Homo habilis |
KNM ER 1813
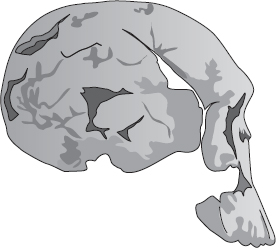
KNM ER 1470
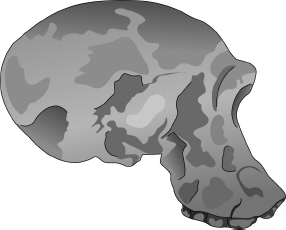
Twiggy
Ska vi låta en sådan röra av namn rubba vårt förtroende för den evolutionära forskningen? Absolut inte. Det är precis vad vi bör vänta oss med tanke på att dessa varelser alla är evolutionära mellanformer, länkar som tidigare felades men inte felas längre. Vi borde vara allvarligt oroliga om det inte fanns några mellanformer så nära gränslinjerna att de skulle vara svåra att klassificera. Enligt den evolutionära uppfattningen skulle det bli omöjligt att sätta åtskilda namn om den fossila dokumentationen vore fylligare. På sätt och vis är det lyckosamt att fossil är så sällsynta. Om vi hade en fortlöpande och obruten serie fossil skulle det bli omöjligt att ge distinkta namn på arter och släkten, eller åtminstone högst problematiskt. Det är en rimlig slutsats att den främsta källan till oenighet bland paleoantropologerna – om det eller det fossilet tillhör ett visst släkte/en viss art eller inte – är djupt och fascinerande meningslös.
Vi tänker oss hypotetiskt att vi av någon lyckträff har välsignats med en löpande fossil dokumentation över alla evolutionära förändringar, utan några felande länkar någonstans. Gå nu tillbaka till de fyra latinska namn som 1470 har fått. Vid första ögonkastet verkar bytet från habilis till rudolfensis vara mindre betydande än det från Australopithecus till Homo. Två arter inom ett släkte är mer lika varandra än två släkten, eller hur? Är inte det hela grunden för skiljelinjen mellan släktnivån (till exempel Homo eller Pan som alternativa släkten av afrikanska apor) och artnivån (till exempel troglodytes eller paniscus inom schimpanserna) i klassificeringens hierarki? Jodå, det är riktigt när vi klassificerar moderna djur, som kan betraktas som ändarna på det evolutionära trädets kvistar, med deras föregångare inuti trädets krona alla behändigt döda och ur vägen. De kvistar som förenas med varandra längre bak (längre in i trädkronans inre) tenderar naturligtvis att bli mindre lika varandra än de som förenas närmare ändarna (som har en gemensam förfader närmare i tiden). Systemet fungerar så länge vi inte försöker klassificera de döda förfäderna. Men så snart vi tar med vårt hypotetiskt kompletta fossilregister faller alla de prydliga separationerna sönder. Som en generell regel blir åtskilda namn omöjliga att tilllämpa. Detta framgår tydligt om vi går bakåt genom tiden i jämn takt, ungefär som vi gjorde med kaninerna i kapitel 2.
Medan vi spårar den moderna Homo sapiens härkomst kommer vi oundvikligen till en tidpunkt då olikheten jämfört med vår tids människor är tillräckligt stor för att motivera ett annat artnamn, till exempel Homo ergaster. Men för varje steg på vägen var individerna förmodligen tillräckligt lika för att deras föräldrar och barn skulle placeras i samma art. Nu går vi längre tillbaka och spårar Homo ergasters härkomst, och vi kommer oundvikligen fram till individer som är tillräckligt olika ”huvudfårans” ergaster för att ett annat artnamn, till exempel Homo habilis, ska vara motiverat. Och nu är vi framme vid poängen med detta resonemang. När vi går ännu längre bakåt måste vi någonstans träffa på individer som är tillräckligt olika den moderna Homo sapiens för att vara förtjänta av ett annat släktnamn, till exempel Australopithecus. Problemet är att ”tillräckligt olika den moderna Homo sapiens” är något helt annat än ”tillräckligt olika den äldsta Homo”, som här betecknas som Homo habilis. Det första exemplaret av Homo habilis hade föräldrar som tillhörde Australopithecus. Hörde hon verkligen till ett annat släkte än sina föräldrar? Det är ju orimligt! Ja, det är det. Men det är inte verkligheten det är fel på, det är vår outrotliga benägenhet för att placera in allting i namngivna kategorier. I verkligheten har det aldrig funnits något första exemplar av Homo habilis. Det fanns aldrig något första exemplar av någon art eller något släkte eller någon ordning eller någon klass eller något phylum. Alla varelser som någonsin har kommit till världen skulle ha klassificerats som tillhöriga exakt samma art som sina föräldrar och barn, om det hade funnits någon zoolog i närheten som genomförde klassificeringen. Men med den moderna forskningens facit i hand och med fördelen – ja, i denna enda paradoxala bemärkelse fördelen – med att de flesta länkarna felas kan vi klassificera i klart åtskilda arter, släkten, familjer, ordningar, klasser och phyla.
Jag önskar att vi verkligen hade en komplett och obruten serie fossil, en bilddokumentation av hela den evolutionära förändringen sådan den gick till. Det önskar jag inte minst för att jag skulle vilja se alla zoologer och antropologer som ligger i livslånga fejder med varandra om huruvida det och det fossilet tillhör den eller den arten, eller det eller det släktet, stå där som fån. Ni käbblar om ord, inte om verkligheten. Som Darwin själv sade i Om arternas uppkomst: ”I en serie former som omärkligt går från någon apliknande varelse till människan sådan hon nu existerar, vore det omöjligt att fastslå någon bestämd punkt där termen ’människa’ bör användas.”
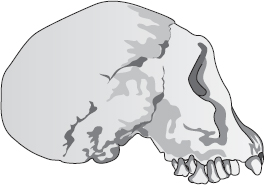
Nu går vi vidare genom fossilen och söker efter några senare länkar bland dem som inte längre felas, även om de felades på Darwins tid. Vilka mellanformer kan vi träffa på mellan oss själva och till exempel 1470 och Twiggy, som ibland kallas Homo, ibland Australopithecus? Vi har redan mött några, bland andra javamänniskan och pekingmänniskan, som brukar klassificeras som Homo erectus. Men de båda levde i Asien, och det finns tillförlitliga belägg för att större delen av vår mänskliga evolution ägde rum i Afrika. Javamänniskan och pekingmänniskan och deras sort hade utvandrat från moderkontinenten Afrika. Inom själva Afrika klassificeras deras motsvarigheter numera i regel som Homo ergaster, trots att de i många år alla kallades Homo erectus – ännu en illustration av hur obeständiga våra namngivningsprocedurer är. Det mest berömda exemplaret av Homo ergaster, och ett av de mest kompletta prehumana fossil som någonsin påträffats, är Turkanapojken eller Nariokotomepojken. Det upptäcktes av Kamoya Kimey, den skickligaste av Richard Leakeys alla paleontologer på att hitta fossil.
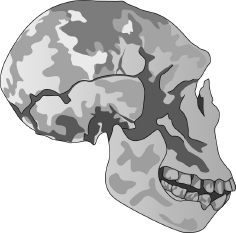
Homo erectus
Turkanapojken levde för cirka 1,6 miljoner år sedan och dog då han var ungefär elva år. Det finns tecken på att han skulle ha blivit 1,80 lång om han hade levt till vuxen ålder. Hans vuxna hjärnvolym har projicerats till cirka 900 kubikcentimeter. Detta var typiskt för hjärnan hos Homo ergaster/erectus som varierade kring 1 000 kubikcentimeter. Det är betydligt mindre än den moderna mänskliga hjärnan, som varierar mellan 1 300 och 1 400 kubikcentimeter, men större än den hos Homo habilis (omkring 600 cm3), som i sin tur var större än den hos Australopithecus (omkring 400 cm3) och schimpanser (ungefär samma volym). Vi kom ju fram till att vår förfader från 3 miljoner år bakåt i tiden hade en hjärna stor som schimpansens men gick på bakbenen. Med ledning av detta kan vi förmoda att historiens andra hälft, från 3 miljoner år till våra dagar, skulle handla om ökande hjärnvolym. Och det gör den mycket riktigt.
Homo ergaster/erectus, som vi har många fossil av, är en mycket övertygande, inte längre felande länk halvvägs mellan dagens Homo sapiens och Homo habilis för två miljoner år sedan som kan sägas ha varit en schimpans som gick på två ben. Hur många länkar behövs innan man går med på att de inte längre ”felas”? Och kan vi också fylla luckan mellan Homo ergaster och den moderna Homo sapiens? Ja. Vi har en rik åder av fossil som täcker några hundra tusen år bakåt och är mellanformer i den långa serien. Några har fått artnamn, som Homo heidelbergensis, Homo rhodesiensis och Homo neanderthalensis. Andra (och ibland samma) kallas ”arkaiska” Homo sapiens. Men som jag framhåller gång på gång spelar namnen ingen roll. Det som betyder något är att länkarna inte längre felas. Det vimlar av mellanformer.
Vi har alltså utmärkt fossildokumentation av en successiv förändring, hela vägen från Lucy, den ”upprättgående schimpansen” från 3 miljoner år tillbaka i tiden. Hur klarar historieförnekarna av detta bevismaterial? Några förnekar det bokstavligt. Det stötte jag på 2008 i en intervju jag gjorde för Channel Fours TV-dokumentär The Genius of Charles Darwin. Jag intervjuade Wendy Wright, ordförande för ”Concerned Women for America”. Hennes åsikt att ”dagen efter-pillret är pedofilens bäste vän” ger en ganska klar bild av hennes förmåga att föra ett resonemang, och i vårt samtal besannade hon alla mina föraningar på den punkten. Bara en liten del av intervjun kom med i dokumentären. Här följer en mycket fullständigare utskrift, men för detta kapitels syfte har jag begränsat mig till de avsnitt där vi talade om människans härkomst dokumenterad av fossil.
Wendy: Vad jag hänvisar till är att evolutionisterna fortfarande inte har någon forskning att stödja sin hypotes på. Det som händer i stället är att forskningsresultat som inte stöder evolutionen blir bortcensurerade. Till exempel att man inte kan bevisa evolutionen genom att gå från en art till en annan. Om evolutionen hade ägt rum skulle det väl finnas åtminstone ett enda bevis för den, om den nu går från fåglar till däggdjur eller till och med längre än så.
Richard: Det finns massor av bevis, men ni på er sida upprepar ständigt det där som ett mantra därför att ni bara lyssnar till varandra. Om ni bara ville öppna ögonen och studera bevisen.
Wendy: Visa dem för mig, visa mig benen, visa mig den döda kroppen, visa mig bevisen för de mellanliggande stadierna från en art till en annan.
Richard: Varenda gång man hittar ett fossil som ligger mellan två arter säger ni: ”Aha, nu har vi två luckor där det bara fanns en tidigare.” Näsan vartenda fossil man hittar representerar en mellanform mellan två arter.
Wendy [skrattar]: Om det vore så skulle Smithsonian Natural History Museum vara proppfullt av sådana exempel men det är det inte.
Richard: Det är det, det är det … i fråga om människor finns det nu sedan Darwins tid en enorm mängd bevis i mänskliga fossil för mellanformer och det finns olika arter till exempel av Australopithecus, och så har vi Homo habilis – de är mellanformer mellan Australopithecus som var en äldre art och Homo sapiens som är en yngre art. Varför kan du inte betrakta dem som mellanformer?
Wendy: … om ni verkligen hade bevis för evolutionen skulle de visas upp på museer, inte bara som illustrationer.
Richard: Jag berättade ju just för dig om Australopithecus, Homo habilis, Homo erectus, Homo sapiens – den arkaiska Homo sapiens och sedan den moderna Homo sapiens – det är en perfekt serie mellanformer.
Wendy: Ni saknar fortfarande konkreta bevis så …
Richard: De konkreta bevisen finns där. Gå till museet och titta på dem … Jag har dem förstås inte här, men du kan gå till vilket museum och se Australopithecus, du kan se Homo habilis, du kan se Homo erectus, du kan se den arkaiska Homo sapiens. En utsökt serie mellanformer. Varför säger du hela tiden ”Lägg fram bevisen” när jag redan har gjort det? Gå till museet och se själv.
Wendy: Det har jag gjort. Jag har gått till museerna och vi är många som ändå inte är övertygade …
Richard: Har du sett Homo erectus?
Wendy: Och jag tycker att ni försöker övertyga oss och censurera oss på ett ganska aggressivt sätt. Det verkar bero på att ni är frustrerade över att så många fortfarande inte tror på evolutionen. Om evolutionisterna vore så säkra på sin sak skulle de inte behöva censurera bort information. Det visar att evolutionen fortfarande inte kan bevisas och är tvivelaktig.
Richard: Jag är … jag medger att jag är frustrerad. Det är inte fråga om att tysta ner er, det handlar om att jag har berättat för dig om fyra fem fossil … [Wendy skrattar] och du bryr dig helt enkelt inte om vad jag säger … Varför går du inte och tittar på de fossilen?
Wendy: … Om de fanns på museerna där jag har varit många gånger skulle jag betrakta dem objektivt, men vad jag vill komma tillbaka till är …
Richard: De finns på museet.
Wendy: Det jag vill komma tillbaka till är att evolutionens filosofi kan leda till ideologier som har varit djupt destruktiva för människosläktet …
Richard: Ja, men vore det inte en god idé om du försökte förstå darwinismen, i stället för att peka på missuppfattningar av darwinismen, som har missbrukats på det hemskaste sätt politiskt. Då skulle du kunna motarbeta dessa förfärliga missförstånd.
Wendy: Vi pressas faktiskt ofta av aggressiviteten av dem som tror på evolutionen. Vi slipper sannerligen inte undan denna information som ni hela tiden lägger fram. Den är inte okänd för oss, för vi kan inte komma undan den. Den prackas på oss hela tiden. Men jag tror att du är frustrerad därför att så många av oss som har sett din information fortfarande inte köper din ideologi.
Richard: Har du sett Homo erectus? Har du sett Homo habilis? Har du sett Australopithecus? Den frågan har jag ställt till dig.
Wendy: Det jag har sett är att när de än påstår sig visa de evolutionära skillnaderna mellan olika arter på museerna och i läroböckerna bygger det hela tiden på illustrationer och teckningar … inte på konkreta bevis.
Richard: Du måste kanske resa till museet i Nairobi för att se originalfossilen, men du kan se avgjutningar av dem – exakta kopior av dessa fossil i alla större museer du gör dig besvär att gå till.
Wendy: Jag förstår inte varför du är så aggressiv. Varför är det så viktigt för dig att alla tror på samma sak som du?
Richard: Det är inte fråga om att tro, jag talar om fakta. Jag har berättat för dig om vissa fossil, och varje gång jag frågar dig om dem smiter du undan och börjar prata om något annat.
Wendy: … Det borde finnas tonvis med konkreta bevis inte bara för en isolerad företeelse, men återigen finns det inga bevis.
Richard: Jag råkade ta hominidfossil därför att jag trodde att de skulle intressera dig mest, men du kan hitta liknande fossil från vilken grupp ryggradsdjur du vill.
Wendy: Jag går tillbaka till varför det är så viktigt för dig att alla andra ska tro på evolutionen …
Richard: Jag gillar inte ordet tro. Jag föredrar att bara be människor granska bevisen, och jag ber dig granska bevisen. … Jag vill att du ska gå på museer och se på fakta och inte tro det du har fått höra om att det inte finns några bevis. Gå bara och se bevisen med egna ögon.
Wendy [skrattar]: Ja, och vad jag skulle vilja säga …
Richard: Det är faktiskt inte roligt. Jag menar, gå på museet, gå dit bara. Jag har berättat för dig om hominidfossil, och du kan gå och se hästens evolution, du kan gå och se de första däggdjurens evolution, du kan gå och se fiskarnas evolution, du kan gå och se övergången från fisk till landlevande amfibier och reptiler. Och du hittar allt detta på vartenda museum av kvalitet. Öppna ögonen bara och se på fakta.
Wendy: Och jag skulle vilja säga öppna ögonen själv och se på de samhällen som har byggts av dem som tror på en kärleksfull Gud som har skapat oss alla …
Jag verkar kanske onödigt obstinat i denna replikväxling med att hamra in uppmaningen till henne att gå på ett museum och se med egna ögon, men jag menade det verkligen. Dessa människor har blivit tränade att säga: ”Det finns inga fossil, låt mig se beviset, visa mig ett enda fossil …” och de säger det så ofta att de har lärt sig att tro på det. Därför gjorde jag experimentet att nämna tre fyra fossil för denna kvinna och inte låta henne slippa undan med att inte låtsas om dem. Resultaten är nedslående och ett utmärkt exempel på den vanligaste taktik som historieförnekarna tar till när de ställs inför historiska bevis – bara nonchalera dem och upprepa mantrat: ”Visa mig fossilen. Var finns fossilen? Det finns inga fossil. Visa mig bara ett enda fossil av en mellanform, det är det enda jag begär …”
Andra villar bort sig i namn och den ofrånkomliga tendens namn har att bilda falska uppdelningar när det inte finns några. Varje fossil som eventuellt kan vara en mellanform klassificeras alltid som antingen Homo eller Australopithecus. Den mest perfekta mellanform man någonsin kan tänka sig skulle fortfarande vara inklämd i antingen Homo eller Australopithecus. I själva verket skulle den ena hälften av paleontologkåren troligen kalla den för Homo och den andra hälften för Australopithecus. Och i stället för att gå samman, som paleontologerna borde göra, och enas om att svårtolkade mellanformsfossil är precis vad vi bör vänta oss av evolutionsteorin, kan vi nog tyvärr lita på att de skulle ge oss ett alldeles falskt intryck genom att nästan ta till knytnävarna, så osams är de om terminologin.
Det är ungefär som den juridiska distinktionen mellan vuxen och minderårig. Av juridiska skäl och för att avgöra om en ung människa är gammal nog att rösta eller gifta sig måste man dra en absolut skiljelinje. År 1969 sänktes rösträttsåldern i Storbritannien från tjugoett till arton år (1971 genomfördes samma förändring i USA). Nu talas det om att den ska sänkas till sexton. Men vilken rösträttsåldern än är finns det ingen som på allvar tror att man blir en annan människa efter tolvslaget på natten till sin artonårsdag (eller tjugoettårsdag eller sextonårsdag). Vi inser naturligtvis alla att hela uppväxtperioden är en enda lång övning i utveckling. En del av oss växer egentligen aldrig upp, kan man nog säga. På samma sätt bestod människans evolution, från något som Australopithecus afarensis till Homo sapiens, av en obruten kedja av föräldrar som födde barn som en samtida taxonom med visshet skulle ha fört till samma art som deras föräldrar. I backspegeln, och av skäl som inte ligger fjärran från det juridiska, envisas moderna taxonomer med att sätta en etikett på varje fossil, där det måste stå ungefär Australopithecus eller Homo. Museietiketterna får under inga omständigheter ha texter som ”halvvägs mellan Australopithecus africanus och Homo habilis”. Historieförnekarna kastar sig över den benämningskonventionen som om den vore bevis på att mellanformer saknas i den verkliga världen. Man kan precis lika gärna säga att det inte finns någon åldersgrupp mellan barn och vuxna därför att alla man ser visar sig vara antingen röstande vuxna (arton år och däröver) eller barn utan rösträtt (under arton år). Det är liktydigt med att säga att den juridiska nödvändigheten av en tröskel för rösträttsålder bevisar att det inte finns något uppväxande släkte.
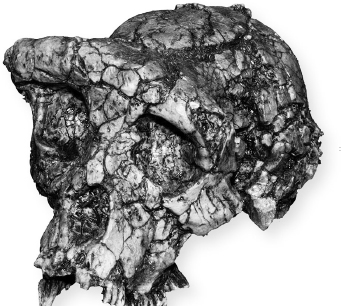
Åter till fossilen. Om kreationisterna har rätt är Australopithecus ”bara en apa”, så dess egna föregångare är ovidkommande för sökandet efter ”felande länkar”. Vi ska ändå kasta ett öga på dem. Det finns några spår, om än ganska fragmentariska. Ardipithecus, som levde för 4–5 miljoner år sedan, är huvudsakligen känd med ledning av tänder, men man har hittat så många skall- och fotben att i varje fall de flesta anatomer som har sett dem säger att den gick upprätt. De som upptäckte två ännu äldre fossil, Orrorin (”millenniemänniskan”) och Sahelanthropus (”Toumai”), har dragit samma slutsats. Sahelanthropus hamnar i en särställning genom att den är mycket gammal (6 miljoner år, nära den förfader vi delar med schimpansen i ålder) och påträffades långt väster om Rift Valley (i Tchad, där ”Toumai” betyder ”hopp om livet”). Andra paleontologer är skeptiska till att Orrorin och Sahelanthropus gick på två ben, så som deras upptäckare hävdar. För varje sådant problematiskt fossil återfinner man några tvivlare bland dem som har upptäckt andra, tillägger kanske cynikern.
Mer än andra vetenskapliga discipliner är paleoantropologin notoriskt hemsökt – eller upplivad? – av rivalitet. Vi måste medge att ett fossilregister som binder samman den upprättgående apan Australopithecus och den (förmodligen) fyrfota förfader som vi delar med schimpanserna alltjämt är svagt. Vi vet inte hur våra förfäder reste sig på bakbenen. Vi behöver fler fossil. Men låt oss åtminstone glädjas åt den tillförlitliga fossildokumentation som vi, i motsats till Darwin, har tillgång till och som visar oss den evolutionära övergången från Australopithecus, med dess hjärna av schimpansstorlek, till den moderna Homo sapiens med vår ballongliknande skalle och stora hjärna.
Sahelanthropus
I hela detta avsnitt har jag återgett bilder av kranier och uppmanat alla att jämföra dem. Det framskjutande käkpartiet eller ögonbrynsbågen faller i ögonen. Ibland är olikheterna hos en del fossil subtila, vilket gör det lättare att uppfatta de successiva övergångarna från ett äldre fossil till ett yngre. Men nu ska jag föra in en komplikation, som blir till ett eget intressant ämne att studera. De förändringar som äger rum inom en individs livstid, medan den blir vuxen, är hur som helst mycket mer dramatiska än de förändringar vi ser när vi jämför vuxna i på varandra följande generationer.
Kraniet på s. 194 tillhör en schimpans kort före födseln. Den är givetvis helt olik den vuxna schimpansskallen på s. 176 och mycket mer lik en mänsklig (vuxen såväl som spädbarn). Det finns en ofta återgiven bild av en nyfödd och en vuxen schimpans, som får illustrera den intressanta tanken att människan bevarar drag karakteristiska för barndomen in i vuxen ålder (det vill säga vi blir könsmogna innan kroppen är vuxen, vilket inte behöver vara samma sak). Bilden var så bra att jag inte riktigt vågade tro på den, så jag skickade den till min kollega Desmond Morris och bad honom om ett expertutlåtande. Kunde den vara en förfalskning? Hade han någonsin sett en schimpansunge som såg riktigt så mänsklig ut? Morris är skeptisk till ryggen och axlarna, men själva huvudet har han inga invändningar mot. ”Schimpanser har kutande hållning och den här har en märkligt rak mänsklig nacke. Men om man begränsar sig till huvudet är bilden trovärdig.” Sheila Lee, som är förlagets bildredaktör och har skaffat bilderna till den här boken, spårade den ursprungliga källan till detta berömda foto, en expedition till Kongo 1909–15 som var utsänd av American Museum of Natural History. Djuren var döda när de fotograferades, och hon påpekar att fotografen, Herbert Lang, också var djurkonservator. Det vore frestande att anta att schimpansungens egendomligt mänskliga hållning beror på felaktig uppstoppning – om inte Lang enligt museet hade fotograferat sina exemplar innan han stoppade upp dem. Man kan ändå justera en död schimpanskropp på ett sätt som man inte kan göra med en levande. Desmond Morris slutsats ser ut att stå sig. Hållningen på schimpansungens axlar är misstänkt, men huvudet är tillförlitligt.
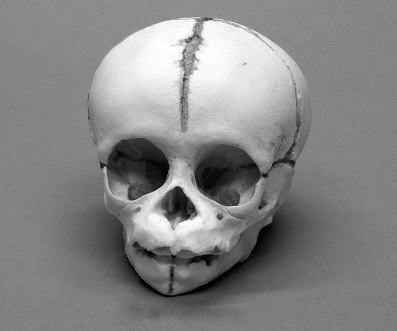
Schimpans kort före födseln
Langs foto av en nyfödd och en vuxen schimpans
Om vi accepterar att huvudet stämmer med verkligheten ser vi genast hur en jämförelse med vuxna fossila skallar kan missleda oss. Eller också kan vi uttrycka det mer konstruktivt och säga att den dramatiska skillnaden mellan vuxna huvuden och barnhuvuden visar hur lätt ett kännetecken som ett framskjutande käkparti kan förändras i exakt rätt riktning för att bli mer – eller mindre – mänskligt. Schimpansens embryologiska utveckling ”vet” hur den ska göra ett människolikt huvud därför att den gör det med alla schimpanser när de passerar barndomsåren. Det förefaller högst sannolikt att Australopithecus på sin väg genom diverse mellanformer till Homo sapiens kortade av käkpartiet hela vägen och gjorde det genom att bevara drag karakteristiska för barndomen in i vuxen ålder (den process som kallas neoteni, omnämnd i kapitel 2). Hur som helst beror den evolutionära förändringen till stor del på förändringar i den takt som vissa delar växer i jämfört med andra delar. Detta kallas heterokronisk (”olika snabb”) tillväxt. Det jag vill ha fram är att den evolutionära förändringen faller sig helt naturlig när man väl har accepterat den embryologiska förändringens konstaterade fakta. Embryon formas av differentierad tillväxt – olika delar av dem växer i olika takt. En nyfödd schimpansunges skalle förändras till en vuxens genom att käkpartiet växer ganska snabbt jämfört med andra ben i skallen. Jag upprepar: Alla djur av alla arter förändras under sin egen embryologiska utveckling långt mer dramatiskt än den typiska vuxna gestalten förändras från generation till generation medan de geologiska tidsåldrarna passerar. Därmed kommer vi in på kapitlet om den embryologiska utvecklingen och dess relevans för evolutionen.
* Till skillnad från sjukdomar, som ofta uppkallas efter den som identifierat dem, får nya arter namn av sina upptäckare men inte efter dem. Det är utmärkt tillfälle för en biolog att hedra en annans namn eller, som i det här fallet, en finansiär. Min framstående kollega W. D. Hamilton hedrades flera gånger på detta sätt, alldeles som sig borde. Han var en av Darwins främsta efterföljare på 1900-talet och hade ett dystert sinnelag som påminner om A. A. Milnes I-or (naturligtvis inte den eländiga Disneyversionen). Hamilton befann sig en gång på en liten båt på en expedition uppför Amazonfloden när han blev stucken av en geting. Hans följeslagare visste att Hamilton var en framstående entomolog och frågade: ”Vet du vad den getingen heter?” ”Ja”, mumlade Bill med sin mest I-orliknande röst. ”Den är uppkallad efter mig.”