ANIMAL THE CANNIBAL: NATURE’S WAY?
Cannibals prefer those who have no spines.
Stanislaw Lem, Holiday
I WAS KNEE-DEEP in a pond that seemed to be composed of equal parts rainwater and cowpats when I felt a nibbling on my leg hair.
‘If you stand still for long enough, they’ll definitely nip you,’ I was informed.
The ‘they’ were spadefoot toad larvae (commonly known as tadpoles) and the warning had come from Dr David Pfennig, a biology professor at the University of North Carolina who had been studying these toads in Arizona’s Chiricahua Mountains for over twenty years.
At Pfennig’s invitation, I had arrived at the American Museum of Natural History’s Southwestern Research Station in mid-July – just after the early summer monsoons had turned cattle wallows into nursery ponds, and newly hatched tadpoles into cannibals. But the real reason I had come wasn’t because the tadpoles were eating each other. It was because some of them weren’t eating each other. In fact, when this particular brood had hatched about a week earlier they were all omnivores, feeding on plankton and general pond detritus. Then, two or three days later, something peculiar had taken place. Some of the tiny amphibians experienced dramatic growth spurts, their bodies ballooning overnight. Now, as I waded, scoop-net in hand, through ‘Sky Ranch Pond’ (a slimy-bottomed mud hole with delusions of grandeur), these pumped-up proto-toads were four or five times larger than their peers. They looked like two different species. I also noted that the larger individuals were light tan in colour while the little guys had bodies flecked with dark green.

Pfennig explained that initially people had indeed thought they were different species, and as I used a magnifying glass to get a better look at my captives I saw that the differences went far beyond body size and colour. The larger tadpoles were also sporting powerful tails and sharp beaks.
‘They’re made of keratin,’ Pfennig explained, referring to the beaks – the same tough, structural protein found in our nails and hair.
Later, while comparing the two tadpole morphs under a dissecting microscope, I saw that behind a set of frilly lips, the flat keratinous plates had been transformed into a row of sharp-edged teeth in the cannibalistic forms. It was also evident that the jaw muscles were significantly enlarged in the cannibals, especially the jaw-closing levator mandibulae. Studies had shown that myofibres, the cells making up these muscles, were larger and greater in number in the large specimens – producing a more powerful bite. Of course, the extra bite force was necessary to subdue and consume their omnivorous pond-mates.
Not quite so obvious was a significant shortening of the gastrointestinal (GI) tract in the cannibals, as a result of the dietary differences that accompanied the tadpole transformations. In the omnivores, a long GI tract is required for the breakdown of tough-to-digest plant matter, while a shorter GI tract suffices for the meat-eaters.1
Over a three-day period, I watched and captured tadpoles. I learned a great deal about the three species of Spea that laid their eggs in such dangerously unpredictable conditions. Much of this information centred on their ecology, behaviour and evolution, and, of course, their cannibalism – something the local research teams seemed to view as perfectly normal.
Until relatively recently, cannibalism in nature would have been regarded as anything but normal. As a result, until the last two decades of the twentieth century, relatively few scientists spent any time studying a topic they regarded as biologically insignificant. The party line was that cannibalism, when it did occur, was the result of either starvation or the stresses related to captive conditions.
It was as simple as that.
Or so we thought.
IN THE 1970S Laurel Fox, a University of California Santa Cruz ecologist, took some of the first steps towards a scientific approach to cannibalism. She had been studying the feeding behaviour of predatory freshwater insects called backswimmers. Fox determined that, while the voracious hunters relied primarily on aquatic prey, cannibalism was also a consistent part of their diets.
I contacted Fox and asked her about the resulting transition that had taken place in the scientific community. She told me that her observations in the field had sparked her interest and that, soon after, she began compiling a list of research papers in which cannibalism had been reported. Although there turned out to be hundreds of references documenting the behaviour in various species, no one had linked these instances together or come up with any general rules. By the time Fox’s review paper came out in 1975, she had concluded that cannibalism was not abnormal behaviour at all, but a completely normal response to a variety of environmental factors.
Significantly, Fox also determined that cannibalism was a far more widespread occurrence than anyone had previously imagined, and that it took place in every major animal group, including some that were long considered to be herbivores, such as butterflies. She emphasised that cannibalism in nature, which some researchers referred to as ‘intraspecific predation’, also demonstrated a complexity that seemed to match its frequency. Fox suggested that the occurrence of cannibalism in a particular species often depended on variables like population density and changes in local environmental conditions. Fox even followed cannibalism on the human branch of the evolutionary tree. After pondering reports that ancient cultures practising non-ritual cannibalism lived in ‘nutritionally marginal areas’, she proposed that consuming other humans might have provided low-density populations with 5–10 per cent of their protein requirements. Conversely, she suggested that cannibalism was rare in settlements where populations were dense enough to allow for the production of an adequate and predictable food supply.
In 1980 ecologist and scorpion expert Gary Polis picked up the baton and began looking at invertebrates that consumed their own kind. Like Fox, he noted that, while starvation could lead to increases in the behaviour, it was certainly not a requirement. Perhaps Polis’s most important contribution to the subject was assembling a list of general rules under which most examples of invertebrate cannibalism could be placed, namely:
1. immature animals get eaten more often than adults;
2. many animals, particularly invertebrates, do not recognise individuals of their own kind, especially eggs and immature stages, which are simply regarded as a food source;
3. females are more often cannibalistic than males;
4. cannibalism increases with hunger and a concurrent decrease in alternative forms of nutrition; and
5. cannibalism is often directly related to the degree of overcrowding in a given population.
Polis emphasised that these general rules were sometimes found in combination, such as overcrowding and a lack of alternative forms of nutrition (a common cause and effect), both of which now fall under the broader banner of ‘stressful environmental conditions’.2
In 1992 zoologists Mark Elgar and Bernard Crespi edited a scholarly book on the ecology and evolution of cannibalism across diverse animal taxa. In it, they refined the scientific definition of cannibalism in nature as ‘the killing and consumption of either all or part of an individual that is of the same species’. Initially the researchers excluded instances where the individuals being consumed were already dead or survived the encounter – the former they considered to be a type of scavenging. Eventually, though, they decided these were variants of cannibalistic behaviour observed across the entire animal kingdom. Although there are certainly grey areas (encompassing things like breast feeding or eating one’s own fingernails), my fallback definition of cannibalism is this: The act of one individual of a species consuming all or part of another individual of the same species. In the animal kingdom this would include scavenging (as long as the scavenged body was from the same species as the scavenger) and maternal care in which tissue (e.g. skin or uterine lining) was consumed. In humans, cannibalism would extend beyond the concept of nutrition into the realms of ritual acts, medicine and mental disorder.
As the study of cannibalism gained scientific validity in the 1980s, more and more researchers began looking at the phenomenon, bringing with them expertise in a variety of fields. From ecologists we learned that cannibalism was often an important part of predation and foraging, while social scientists studied its relationship to courtship, mating and even parental care. Anatomists found strange structures to examine (like the keratinous beak of the spadefoot toad) and field biologists studied cannibalism under natural conditions, thus countering the previous belief that the behaviour was captivity-dependent.
By the 1990s, Polis’s observations had been confirmed among widely divergent animal groups, both with and without backbones, supporting the conclusion that the benefits of consuming your own kind could outweigh the often substantial costs. Once these general rules were established, and as a new generation of researchers built upon foundations constructed by pioneers like Fox and Polis, cannibalism in nature, with all of its intricacies and variation, began to make perfect evolutionary sense.
Arizona’s lowland scrub stood in stark contrast to the lush peaks and boulder-strewn valleys of the state’s Chiricahua Mountains. These ‘sky islands’ provided a spectacular backdrop to examine yet another transient pond.
The air temperature had risen to ninety-five degrees Fahrenheit, which kept most of the area’s land-dwellers hiding in shade or below ground, but the inhabitants of Horseshoe Pond reminded me of overexcited kids tearing around a playground. By this time, I had already begun to see distinct patterns of behaviour in the spadefoot tadpoles that motored hyperactively just below the water’s surface.
I noticed that the smaller omnivores generally stuck to the shallows bordering the shoreline. They buzzed through the brown water in a non-stop, seemingly random quest for food, changing direction abruptly and often. One explanation for the patternless swimming became apparent as I waded further away from the shore, for here in the deeper water was the realm of the cannibals. I stood quietly and watched as hundreds of conspicuously larger tadpoles crisscrossed the pond, making frequent excursions from the deeper water toward the shore in a relentless search for prey.
‘They remind me of killer whales hunting for seals,’ said Ryan Martin, a former student of Pfennig’s, now a professor at Case Western Reserve, who was also studying spadefoot toads here in Arizona.
The comparison was apt. But the question remained: why did the local spadefoot larvae exhibit cannibalistic behaviour? There certainly seemed to be enough organic matter suspended in these algae-tinted ponds to feed them all and more.
As I spoke to Pfennig and his team of researchers, I learned that the answer was directly linked to the aquatic environments where the parents deposited their eggs.3 Formed by spring and early summer monsoons, the transient ponds frequented by the spadefoots (spadefeet?) are often little more than puddles, and as such they can evaporate quite suddenly in the hot, dry environment of south-eastern Arizona. Natural selection, therefore, would favour any adaptation enabling the water-dependent tadpoles to ‘get out of the pool’ as quickly as possible. In this instance, the phenomenon that evolved can be filed under the rather broad ecological heading of phenotypic plasticity: when changing environmental conditions allow multiple characteristics or traits to arise from a single genotype (the genetic makeup of an organism).
One (non-cannibalistic) example of this is the water flea (Daphnia spp.). Water fleas are tiny aquatic crustaceans, named for a swimming style in which they appear to jump. In response to the appearance of backswimmers, they develop tail spikes and protective crests. Although the genetic potential for body armour is always there, it doesn’t exhibit itself until a specific environmental change occurs, in this case the arrival of Daphnia-munching backswimmers.

Another example of this is the bluehead wrasse, a reef-dweller famous for its habit of removing parasites from much larger fish, even entering into their mouths. Here, however, it’s the removal of a male wrasse from its harem of thirty to fifty females that alters their local environment. Rather than waiting for a new male to arrive, something extraordinary takes place in the harem. Within minutes, one of the females begins exhibiting male-typical behaviours. Relatively quickly, the former female transforms into a male. The opposite can also occur, as in the case of the clownfish, whose real-life behaviour could have offered an intriguing alternative resolution to Finding Nemo.
In spadefoot toads, though, it’s not the appearance of a predator or the loss of a harem’s personal sperm bank that initiates the alternate phenotype (the cannibalistic larvae). The selection pressure lies in the temporary nature of the ponds where the eggs are deposited and hatch, and where the tadpoles develop. The period from egg to juvenile toad usually takes around thirty days unless the pond dries out first, killing the entire brood. In response to this particular environmental selection pressure, spadefoot toads evolved a means by which some of the tadpoles can mature in about two-thirds of the time (twenty days). The increased growth rate occurs because the cannibal larvae are getting a diet high in animal protein as well as a side order of veggies, the latter in the form of nutrient-rich plant matter consumed by their omnivorous prey.
It’s interesting to note that a related species of spadefoot, Spea couchii, does not have this ability to transform into cannibalistic morphs but has evolved an alternative solution to the transient-pond problem: they can go from egg to toad in only eight days – an amphibian record.
Though the spadefoot toad story has been well researched, it is not fully resolved. No one has been able to identify the precise stimulus within these brood ponds that triggers the appearance of the cannibal morphs. Until recently, the prime candidates were a pair of microscopic fairy shrimp species. David Pfennig and his colleagues proposed that the consumption of the shrimp by some of the spadefoot tadpoles served to trigger the cascade of genetically controlled developmental changes that transformed the shrimp-eaters into outsized cannibals.
But what was it that set this transformation into motion? Pfennig hypothesised that iodine-containing compounds found in the shrimp might cause the activation of specific genes in the tadpoles, genes that weren’t turned on in the individuals that didn’t consume shrimp. The most likely trigger substance turns out to be thyroxin: a thyroid hormone whose functions include stimulating metabolism and promoting tissue growth. However, this theory was undermined by experiments showing that even tadpoles not fed fairy shrimp could still undergo the transformation to cannibals, indicating that (at the very least) something besides thyroxin intake must initiate the changes.
‘What if it’s not what they’re eating but the mechanism of chewing itself that serves as a tactile trigger?’ I made the suggestion while brainstorming the problem with biologist Ryan Martin. ‘What if chewing on something alive like a fairy shrimp, something larger or something that struggles when you clamp onto it, sets this developmental cascade into motion?’
Martin shot me a ‘not bad for a bat biologist’ look. ‘Sounds like a good grad student project.’
‘Hey, it’s all yours,’ I said with a laugh. We then set to work, drawing up an outline for a potential experiment to test the hypothesis.
Although the jury is still out, Pfennig and his co-workers have previously worked on a completely different cannibalism trigger in another amphibian. And this one happened to be one of North America’s most spectacular species.
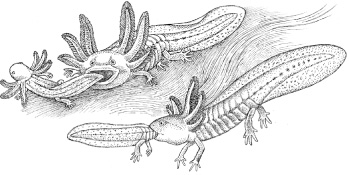
Tiger salamanders are the largest salamanders in the United States, reaching lengths of up to thirteen inches. These thick-bodied, sturdy-limbed urodelans are widespread across much of the country.4 Their markings, yellow blotches against a black body, make them easy to identify but they are rarely seen in the open except during annual marches to a nuptial pond. Tiger salamander eggs are laid in the late winter or early spring and, like other salamanders (and their cousins the frogs and toads), their larvae are fully aquatic with external gills and fish-like tails. They typically feed on zooplankton and other micro-invertebrates, but under certain environmental conditions a small percentage of them develop traits that include huge heads, wide mouths and elongated teeth. Consequently, these toothy individuals exploit larger prey, which includes other tiger salamander larvae.
Pfennig and his colleagues set up lab experiments on their fertilised eggs to investigate the stimuli that set these changes into motion. First, the researchers determined that the cannibal morphs only developed when larvae were placed into crowded conditions. Next, they used a variety of experiments to determine whether the larval transformation might be triggered by visual cues, smell, or touch. They concluded that the tactile cues were significant: the act of bumping into each other was key in the production of the cannibals.
Movement- and touch-related, I thought, remembering my suggestion about a possible tactile trigger for the spadefoot cannibals. But instead of speculating about my own half-formed ideas, the conversation turned toward the pros and cons of cannibalism, especially as it pertained to consuming kin.
As we have seen, one of Gary Polis’s general rules about cannibalism is that immature animals get eaten far more often than adults. Ultimately, this makes larvicide (or infanticide) the most common form of cannibalism in the animal kingdom. Intuitively, it doesn’t seem logical to eat the next generation, but the behaviour can make evolutionary sense for several reasons. Young animals not only provide a valuable source of nutrition but in most species are also relatively defenceless. As such, they present instant nutritional benefit but little or no threat to larger members of the same species, most of which are invulnerable to attacks from immature forms.
But beyond acquiring a meal, cannibalism enables individuals of species such as spadefoot toads to accelerate their developmental process, thus allowing them to quickly outgrow a stage where they might be preyed upon or perish due to unpredictable environmental conditions. In species like the flour beetle, this may also impart a reproductive advantage, since studies have shown that cannibalistic individuals produce more eggs than non-cannibals. Finally, many animals maintain specific territories, within which they are intolerant to the presence of conspecifics (i.e. members of the same species). According to Polis, crowding increases the frequency with which individuals violate the space of others. By reducing overcrowded conditions, cannibalism can serve to decrease the frequency of territory violations.
But, of course, there are also obvious and serious drawbacks to being a cannibal.
Perhaps the most significant of these is a heightened chance of acquiring harmful parasites or diseases from a conspecific. Both parasites and pathogens are often species-specific and many of them have evolved defences to defeat their host’s immune system. As a result, predators that consume their own kind run a greater risk of picking up a disease or a parasite than predators that feed solely on other species. In the most famous example of cannibalism-related disease transmission, the Fore people of New Guinea were nearly driven to extinction by their ritualised consumption of brains and other tissues cut from the bodies of their deceased kin, kin who had themselves been infected by kuru, an incurable and highly transmissible neurological disease. More on that topic later, but the potential for disease transmission stands as a prime example that non-humans and humans alike share some of the negative consequences of cannibalism.
Cannibals can also experience decreases in a measure of evolutionary success known as ‘inclusive fitness’, in which the survival of an individual’s genes, whether they’re from an offspring or a collateral relative such as a brother, sister, or cousin, is the true measure of evolutionary success. A cannibal that consumes its own issue, siblings or even more distant relatives, removes those genes from the population, so it reduces its own inclusive fitness. Since this is disadvantageous, natural selection should favour cannibals that can discriminate between kin and non-kin.
As a result, it made perfect sense that David Pfennig and his colleagues had also worked on questions related to kin recognition. The researchers found that their study subjects could often identify their own relatives.
They do so by means of what’s known as ‘the armpit effect’, Pfennig told me, whereby an individual forms a template for the scent of its kin based on its own smell. He cited a species of paper wasps that regularly raid the nests of other paper wasps to provide food for their own. In these species, individuals learn that if an individual smells like your nest or burrow … you don’t eat them.
Similarly, tiger salamander larvae are more likely to eat the larvae of unrelated individuals than they are to consume relatives. Pfennig explained that he and his colleagues determined this experimentally by preventing them from being able to smell by applying superglue to their nares.
In the wild, cannibal tadpoles have been found to release siblings unharmed yet consume non-relatives. However, in the lab, apparently all bets are off if the cannibals are deprived of food then placed in a tank with other tadpoles. In these cases, starvation becomes the great equaliser and both kin and non-kin are eaten. As I would learn from researchers, unearthing new evidence about the behaviour in humans, this particular aspect of cannibalism spans the entire animal kingdom.
On the plane ride back to New York, I thought a great deal about the cannibalism I’d seen in the temporary ponds.
Cannibal morphs.
I wondered whether H. G. Wells knew about their existence when he wrote The Time Machine in 1895. In Wells’s classic novel, the Time Traveller encounters two human species: the child-sized and docile Eloi and the brutish Morlocks, who raise the Eloi in order to feed upon them. Wells explained the Morlocks’ cannibalistic behaviour by suggesting that they were once members of a worker class, toiling underground for lazy upper-class surface dwellers. The Time Traveller speculates that a food shortage (i.e. an environmental change) forced the subterraneans to alter their diets – at first rats but ultimately something a bit larger. Eventually, this behaviour resulted in a race of hulking cannibals, feeding on the surface-dwellers, whose own evolutionary path would produce the sheep-like Eloi, pampered, well fed and eventually slaughtered for food.
In his cautionary tale of class distinction, Wells imagined a biological phenomenon remarkably similar to what scientists like David Pfennig and his colleagues are working on today.
Many scientists now believe that phenotypic plasticity offers the perfect building blocks for the type of evolutionary change described by Wells over a century ago. These building blocks could be novel traits like the tiger salamander’s jaws or the spadefoot tadpole’s serrated beak – each having originated as an environment-dependent alternative to an already established ancestral trait. What these scientists hypothesise goes far beyond the realm of cannibalism and into the very mechanisms of evolution itself. Their claim is that the appearance of new traits in a population, generally regarded as a first step toward the evolution of new species, can occur by means other than the accumulation of micromutations, the classic mechanism by which new traits, and eventually new species, are thought to appear. Some researchers now believe that in given generations, novel traits originating as examples of phenotypic plasticity even have the potential to produce separate species.

This idea originated with the German/American geneticist Richard Goldschmidt (1878–1958), infamous for his stance that micromutations accumulating over long periods of time were inadequate to explain the evolution of different species. He proposed two additional mechanisms; the first, speciation by macromutations (i.e. those causing a profound effect on the organism), eventually led to the derision associated with his name and the legacy-destroying label of ‘non-Darwinian’. Less well known is Goldschmidt’s suggestion (quite correct, it appears) that mutations can result in major changes during early development, and that these can lead to large effects in the adult phenotype. This hypothesis and the related concept of developmental adaptability are two of the key principles of the modern field of evolutionary developmental biology, informally known as ‘evo-devo’.
Okay, so now that I’d captured and examined cannibalistic tadpole morphs and heard all about their outsized salamander cousins, it was time to look into other examples of cannibalism in nature and to determine why these creatures were eating each other. I decided that the best way to cover and divvy up the material was to look at what I considered to be the most dramatic examples of Gary Polis’s cannibalism-related generalisations. Admittedly, some of what I uncovered was hard to categorise, thus leading me to the realisation that cannibalism can extend far beyond the realm of generalisation. I also learned that, normal behaviour or not, sometimes cannibalism in the animal kingdom can get downright weird.
Footnotes
1 A lengthy intestine is a hallmark of many herbivores since longer guts entail longer passage times for the food moving through them – allowing for additional chemical digestion and more thorough absorption of nutrients. In many animals, though (including all vertebrates and even invertebrates like termites), the digestive tract cannot digest cellulose, the polysaccharide that makes up plant-cell walls. The problem is solved by the presence of endosymbiotic bacteria or protozoans (‘gut flora’) that produce cellulases – enzymes capable of digesting polysaccharides. In ‘foregut fermenters’ like cows, a multi-chambered stomach serves as a homestead for the enzyme-generating microbial horde, while in ‘hindgut fermenters’ like horses, a pouch-like section of the intestine called the cecum houses the endosymbionts.
2 Tragically, Dr Polis drowned when his research vessel sank during a storm in the Sea of Cortez in 2000, an accident that also claimed the lives of a graduate student and three Japanese ecologists.
3 Toads and frogs belong to the amphibian order Anura (from the Greek for ‘no tail’). Most anurans lay their eggs in fresh water, with hatchlings undergoing complete metamorphosis from gill-bearing tadpoles to lung-breathing juveniles.
4 Urodela (Greek for ‘conspicuous tail’) is the order containing approximately 655 salamanders, lizard-shaped amphibians generally found in moist terrestrial environments.