Figure 5.1. World map at the time of the Triassic-Jurassic boundary, 200 million years ago, showing the Pangean supercontinent at its peak size.
TRIASSIC DOWNFALL
The final few million years of the Triassic saw the landmasses of South and North China gradually collide into one another. This was followed by their joint collision, together with other bits and pieces of Southeast Asia, into the northern coast of Pangea. The result was a Pangean supercontinent at its zenith; all the pieces of the jigsaw were together (fig. 5.1). It would now have been possible to walk on every piece of continental crust without once having to cross an ocean. No need for boats.
While continental drift was making world geography very simple, life was thriving, having long forgotten the earlier extinctions of Pangea’s history. Following the enigmatic Carnian climatic episode, terrestrial communities were diverse and consisted of large crurotarsans and, to a lesser extent, dinosaurs, plus rarer groups such as the pterosaurs and mammals. Forests were back on the scene and well established, and plants were abundant enough to form coal deposits. Insects were starting to look very modern, with flies and beetles evolving fast alongside more exotic groups like the titanopterids—giant, overgrown grasshoppers with wingspans in excess of 20 centimeters. In the seas coral reefs had appeared in the warm waters of western Tethys, while plankton was undergoing a fundamental change in composition as red replaced green in the great algal changeover. However, this is a book about great disasters, not successes, and once again the restless nature of plate tectonics was to be the undoing of evolution’s hard work. Another gigantic phase of world-ending volcanism was about to strike, this time centered in western Pangea.
Figure 5.1. World map at the time of the Triassic-Jurassic boundary, 200 million years ago, showing the Pangean supercontinent at its peak size.
Andrea Marzoli of the University of Padua, in Italy, is one of the foremost volcanologists to investigate a LIP that he has termed the Central Atlantic Magmatic Province, or CAMP for short. This name is somewhat misleading (or puzzling) because the Atlantic did not exist when CAMP started erupting. However, Andrea chose this epithet because the site of eruption eventually became the central part of the Atlantic as the fragments of Pangea drifted apart over tens of millions of years. A consequence of the separation and drift is the fragmentation of formerly united lava fields. For a long time this meant that the unity of the original province was overlooked, but recent study of the dispersed fragments has allowed geologists to piece together the extent of a formerly huge LIP—its lavas may have exceeded 5 million cubic kilometers in volume.
Two hundred million years later (i.e., today), the effects of erosion and burial have left only remnants of CAMP’s igneous rocks, although some, such as the Palisades Sill of New Jersey and New York, are still impressive. Where the basalts have been eroded, the remaining igneous plumbing system bears testimony to the magnitude of the event: dikes—vertical cracks filled with cooled magma—in Amazonia, for example, are 200 meters wide and 300 kilometers long. A single dike in the Iberian Peninsula has been traced for more than 500 kilometers.
With the identification of the huge scale of CAMP volcanism in the late 1990s came the realization that its being 200 million years old placed it in the frame for the end-Triassic mass extinctions. Although relatively little studied at the time, the crisis has long been recognized as one of the big five, and the identification of such an enormous smoking gun was to spark an intense research effort that continues to this day. Nowadays the end-Triassic catastrophe is one of the most actively investigated of all mass extinctions. The link with LIP volcanism inevitably causes many parallels to be drawn with the Permo-Triassic mass extinction/Siberian Traps nexus, and there are indeed many close similarities. But before racing on to consider the question of how it happened, we must first look at what died and when.
THE VICTIMS
Dinosaurs, the iconic group of paleontology, ruled the world for the 135 million years of the Jurassic and Cretaceous Periods, but as we have seen, they had been around for tens of millions of years prior to this, which raises the question, why did it take them so long to become dominant? The answer seems to be that it took a mass extinction to remove their chief competitors, the crurotarsan groups—such as the aetosaurs, the rauisuchians, and the phytosaurs. These diverse animals were present right up to the end of the Triassic, but none are known to have survived into the Jurassic. The survivors included true crocodiles (the only surviving legacy of the crurotarsan dynasty), pterosaurs, and mammals, but it was the dinosaurs that were able to respond with alacrity to the emptied environments and went on to rapidly fill the earliest Jurassic landscape.
The idea that the crurotarsan/dinosaur transition was caused by a mass extinction was a highly controversial notion when initially suggested more than thirty years ago. Many had thought the Late Triassic had witnessed a prolonged competitive struggle with the dinosaurs finally coming out on top. Subsequent work has now shown that the crurotarsans were in fact doing well until near the end of the Triassic—it needed a catastrophe to remove them from the stage. The dinosaurs therefore owed their success to a combination of luck and an ability to survive the harsh conditions of the extinction interval. However, even if the nature of the transition is now well accepted, the precise timing is still debated. Here we need a digression on the divisions of Late Triassic time. This interval is subdivided, from oldest to youngest, into the Carnian, Norian, and Rhaetian Stages (see fig. 1.1). Some have argued (notably Spencer Lucas of the New Mexico Museum of Natural History) that the main terrestrial animal extinctions occurred at the end of the Norian Stage, whereas others suggest it happened later, toward the end of the Rhaetian Stage. It would be very useful to resolve this debate because the marine extinctions (described below) happened in the late Rhaetian and so may (or may not) coincide with the terrestrial crisis. Our understanding of Late Triassic time is not helped by the great uncertainty regarding the duration of the Rhaetian Stage. Estimates range from a mere million years to as much as 10 million years. The larger estimates make it possible that the terrestrial extinctions may have substantially predated the marine extinctions if the latter occurred at the end of the Norian. This would mean that the two events had nothing to do with each other but were in fact totally separate extinctions—an untidy conclusion. On the other hand, a very short duration for the Rhaetian Stage would mean that the marine and terrestrial crises occurred about the same time, so we could be talking about a single mass extinction.
Some remarkable and pertinent evidence to help resolve the debate comes from southwestern England and Wales, and it is found in Carboniferous limestones. This may sound strange, because Carboniferous rocks formed 150 million years before the Late Triassic. However, by Rhaetian times the limestones in this region had been uplifted to form small islands up to 16 kilometers in extent, which were surrounded by a warm shallow sea. Importantly, the limestone landscape was riddled with caves and fissures. As a result, animals walking around the islands often fell down holes, got trapped, and thereby became fossils. This fissure fauna, as paleontologists have called it, contains a remarkable diversity of animals dominated by small lizards called sphenodontians and other reptile groups. Unfortunately though, it does not provide a complete census of latest Triassic life but rather a subset: a specialized island-dwelling community. It notably lacks the larger crurotarsan animals, probably for the simple reason that it is hard for a big animal to fall down a small hole. However, there are rare finds of somewhat bigger beasts, including aetosaurs, phytosaurs, and dinosaurs, some of which approached two meters in length, so some of the crevices must have been quite large. Mammals, on the other hand, are absent despite their small size, probably because they are not very good at dispersing to islands even today. The high metabolic rate of mammals requires that they eat and drink every day, whereas the process of island colonization often requires that animals be clinging to driftwood for many days or even weeks. This is something that reptiles but not mammals can survive.
Despite the odd composition of the Rhaetian fissure faunas, it significantly contains many animals that failed to survive into the Jurassic. This suggests they are the record of a pre-mass-extinction island community and so supports the idea of a latest Rhaetian age for the terrestrial extinctions. We also have evidence from plant remains that the Late Triassic mass extinction happened in the latest Rhaetian and not earlier.
Sporomorphs (fossil spore and pollen grains) provide much better detail on the age of the end-Triassic extinction because they fossilize easily and are found in many types of sedimentary rocks. The best sporomorph data has come from studies of Triassic-Jurassic boundary sections in northwestern Europe and Greenland. These reveal a late Rhaetian crisis in which there were many interesting changes. Conifers, cycads, and bennettitaleans dominated the Late Triassic forests, but this situation abruptly changed in the later part of the Rhaetian, when fern spores become prolific. This “fern spike” was short-lived though, and conifer-dominated forests reappeared before the end of the Triassic. However, things had changed; the post-fern-spike forests were noticeably less diverse and their composition was different. The main conifers were now the cheirolepidiaceans, an extinct group that resembled modern cypress trees and were thought to prefer warmer and more arid conditions. So, like the plant crisis during the Permo-Triassic transition, the European and Greenland forests disappeared in the Late Triassic, but unlike the Permo-Triassic event, it was a brief phenomenon with plenty of changes and less extinction. The Triassic-Jurassic crisis may also have been of more regional extent.
The sporomorph story from North America is similar to that in northwestern Europe and Greenland, which is perhaps not that surprising given that they were all in the same region of tropical western Pangea in the Late Triassic. Significantly though, plants from more distant locations show much less dramatic changes. Thus, the Junggar Basin of northwestern China lay in northeastern Pangea, and it contains a thick and monotonous pile of fine-grained lake sediments that formed during the Late Triassic and Early Jurassic. Neither the sediments nor the fossil spores and pollen change much at boundary (although a short-lived fern spike may be present), suggesting that there were no major, long-term climatic changes in this area. Similar muted changes in Australia point to the same story in southeastern Pangea. The dramatic change in plant communities in central Pangea may relate to their proximity to the CAMP volcanism and its regionally damaging effects. More distant locations appear to have weathered the crisis somewhat better.
Let’s stay for a moment with the ever-so-useful sporomorph studies. The extraction of such tiny organic fossils from sedimentary rocks also releases other bits of organic detritus, scattered among which are charcoal fragments—the product of past fires. As we look at charcoal amounts, it appears that there was a marked increase in the latest Triassic that persisted for a million or so years into the Jurassic. The evidence for increased burning is also supported by the presence of organic molecules called polycyclic aromatic hydrocarbons (PAHs), which are produced by combustion of vegetation. So, what can we infer from the charcoal/PAHs evidence? Once again the data come from central Pangea, where we know there were substantial vegetation changes. The more frequent wildfires likely reflect increasing aridity and more flammable plants in the changed forests. Some researchers have suggested that it was also a cause of the terrestrial mass extinction. Death by fire certainly sounds like a suitably hellish way of causing extinction, but it is a highly unlikely proposition. It certainly gets no support from the study of modern terrestrial ecosystems. Many habitats actually benefit from fires because they cause openings in forest canopies and increase plant diversity.
Plant fossils, especially of leaves, can also provide clues to atmospheric changes. Leaves have tiny holes in their surface called stomata, which are used for gas exchange—during photosynthesis they let carbon dioxide in and oxygen out. This arrangement is very useful, but the downside of stomata is that they also let water vapor escape and cause desiccation of the plant. The density of stomata on the leaves of plants is therefore a careful balance between having enough for gas exchange but not so many that the leaves dry out too easily. Now when atmospheric carbon dioxide levels are very high, plants can get by with few stomata, but when the levels of the gas fall, then they need more. The stomatal abundance on leaves is thus a monitor of atmospheric carbon dioxide concentrations—a very useful relationship for those who study past climates. Care has to be taken though; for example, if the fossil-leaf record shows a doubling of stomatal density, it is safe to infer that carbon dioxide levels increased, but it does not mean they have doubled; some plants are just more responsive than others. So when dealing with changes in stomatal density, it is important that the same type of plant is being studied, something that is not always possible. Despite such caveats, any method for evaluating past atmospheric gas concentrations is highly desirable, and many studies have looked at leaf stomata across the Triassic-Jurassic transition. Jenny McElwain of University College Dublin has led much of this work, and she has shown that carbon dioxide levels rose steeply in the latest Triassic to reach a peak of about 2500 ppm.15 This high point persisted for around 300,000 years before declining in the early Jurassic. For comparison, preindustrial atmospheric CO2 levels were 280 ppm, and they are now around 400 ppm. It is safe to assume that it got very warm at the end of the Triassic, much warmer than today.
The end-Triassic seas and oceans also saw substantial losses among a whole range of environments and organisms, from Tethyan reefs to the tiny, planktonic radiolarians that drifted in the surface waters of Panthalassa. The most detailed extinction stories have again come from Europe, particularly the coastal cliffs around Britain. These accessible outcrops record a rather unusual depositional setting. A few million years before the end of the Triassic, the sea level rose, causing a marine embayment to develop over a large part of northwestern Europe (leaving islands of limestone with potholes, which were a danger for the resident animals). This Rhaetian Sea invaded a vast and very flat desert plain, and consequently the sedimentary layers that accumulated were of remarkable persistence. I have studied cliffs of Rhaetian rocks on the Devon coast, in southern England, and around Larne, in Northern Ireland, and been able to trace exactly the same series of beds even though the two locations are 600 kilometers apart. In addition to this remarkable uniformity, this vast inland sea was also unusually saline, no doubt because it was surrounded by the deserts of Pangea and subject to intense evaporation. The hypersalinity is manifest by an impoverished fossil content. Animals that were accustomed to normal marine salinity, such as ammonoids, brachiopods, corals, bryozoans, and echinoderms, are mostly absent, whereas groups able to tolerate high (and fluctuating) salinities, such as bivalves and ostracods, are present in abundance. It is the bivalves that tell us most about the extinction in this end-Triassic sea.
The initial sediments to accumulate in the Rhaetian Sea consisted of black shales belonging to the Westbury Formation. These are overlain by the white limestones of the Lilstock Formation, which in turn are overlain by the Blue Lias, a formation of limestone and shale beds that are famous for their Jurassic ammonites. The black, white, and blue stratigraphy provides information on a fascinating series of events (fig. 5.2). The Westbury Formation yields abundant and fairly diverse bivalves, but the majority of species (15 of 25) disappear at the base of the Lilstock Formation, thus marking the level of the mass extinction. About a meter above this level, the beds are intensely contorted in a way that is diagnostic of earthquake shaking. Geologists call such beds seismites, and just above them there is evidence for uplift and emergence in the form of deep desiccation cracks. The next limestone beds look innocuous enough in the field, but a study of the organic carbon isotopes at this level reveals that they are very light; that is, they are enriched in carbon-12 and depleted in carbon-13 compared to the beds on either side. This negative carbon isotope spike is a feature that we have met several times already during Pangea’s earlier mass extinctions, especially during the Permo-Triassic crisis. This is a key level in the succession because it also corresponds to a change in the composition of spore and pollen fossils and is where the fern spike seen elsewhere in Europe and North America developed.
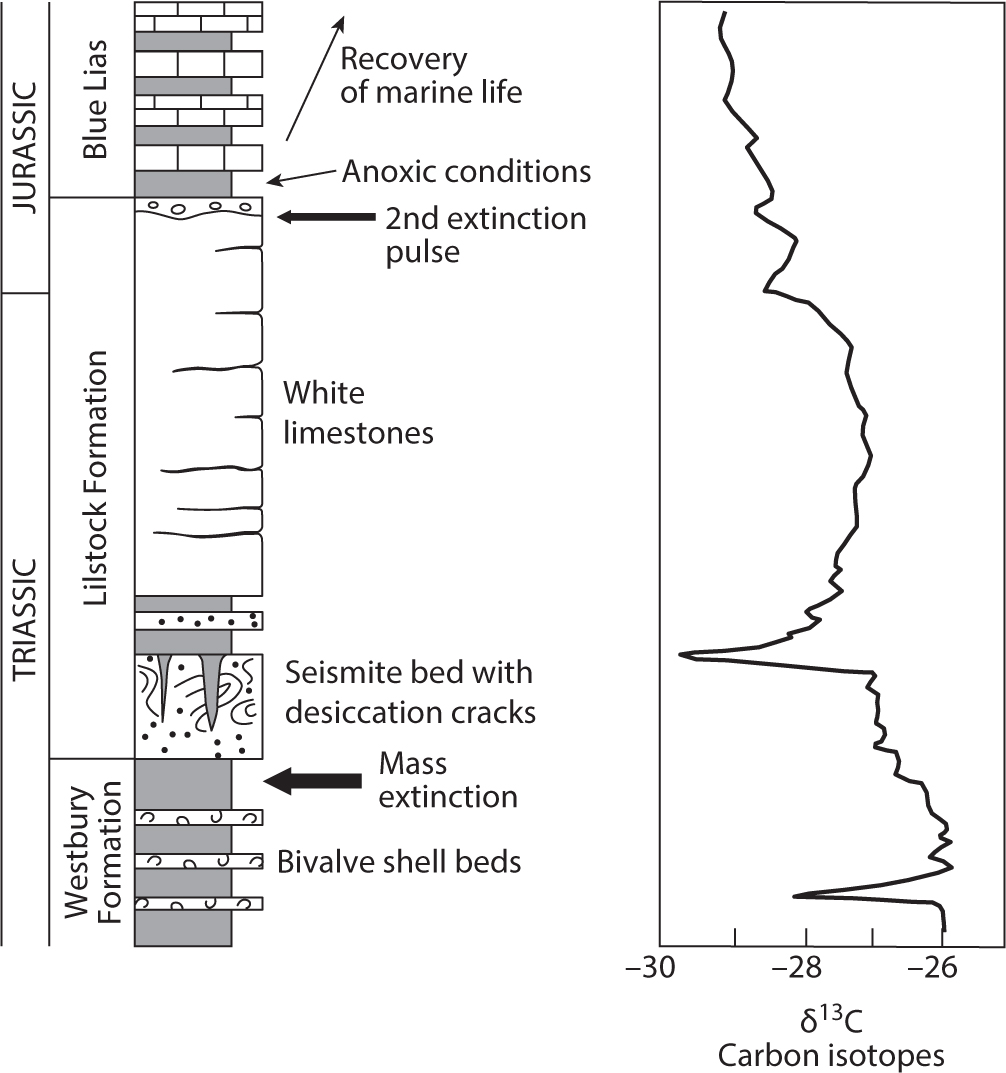
Figure 5.2. Triassic-Jurassic boundary rocks in southwestern England showing the levels of extinction (left) and the carbon isotope values (right) recorded in organic matter.
Higher in the Lilstock Formation, a few new bivalve species appear, indicating the start of recovery, but the real diversity bounce back was not until the beginning of the Jurassic. Prior to this, at the base of the Blue Lias a series of black shales and limestones developed, revealing that seafloor oxygenation was exceptionally poor at this time. There was clearly a lot going on in the late Rhaetian, but perhaps the most surprising aspect is that the first thing to happen—the bivalve mass extinction—predates everything else. By extending our view of the end-Triassic marine extinction to other regions we find that, like the Permo-Triassic mass extinction, the timing of losses varied around the world, and it may be more appropriate to talk of a two-phased crisis once again.
In southern Europe the Triassic-Jurassic boundary is found in shallow marine limestones that form lovely Alpine mountains in western Austria and southernmost Germany. The most profound change here is the extermination of diverse reefs constructed by scleractinian corals. The bivalves also had a tough time here too. Several groups of giant bivalves had evolved in the Late Triassic—the megalodontids, dicerocarditids, and wallowaconchids—and none of them survived into the Jurassic.
The Alpine limestones formed in the warm, shallow, equatorial seas at the western end of Tethys. If we follow the southern coastline of this ocean, we move gradually southward into temperate latitudes, where interbeds of limestone and terrigenous sediments (sandstones and shales) accumulated in shallow seas. Today some of these sedimentary rocks are found at great altitude in the Tibetan Himalayas, where they can be reached with difficulty. Tony Hallam and I visited one of the most accessible locations, close to the Tibetan border around fifteen years ago. I say “accessible” because the rocks were adjacent to the main Lhasa-Kathmandu highway, but this is a subjective term when used to describe the road infrastructure of 1990s Tibet. Tony has recounted some of our escapades on this trip in his book Catastrophes and Lesser Calamities. Other than the discomfort of spending two weeks sharing the back seat of a Mitsubishi Pajaro with three other geologists, I well remember taking a supposed shortcut over a mountain on a narrow, gravelly track that barely clung to the mountainside. When not spending time on death-defying journeys, our accommodation was in a Chinese army barracks, from which we had a splendid view of Mount Everest, but sadly we had no toilet facilities other than a narrow trench next to the parade ground. Overall the Tibetan visit was an ordeal, but it was worth it because it revealed a Triassic-Jurassic mass extinction story that is different from that of the Rhaetian Sea.
The Tibetan sediments formed in open marine conditions in which there was a high diversity of marine invertebrates. Usefully, they included the ammonite Psiloceras, whose first appearance defines the start of the Jurassic. Remarkably, most of the diverse Triassic bivalves survive for a short distance above this level, indicating that the end-Triassic extinction in this region actually happened in the earliest Jurassic. Detailed comparison with the European record is problematic, but it is possible that this later extinction in Tibet corresponds to a weak extinction pulse in Britain. Thus, a few bivalve species are lost at the base of the Blue Lias at the point of onset of black shale deposition (fig. 5.2). If so, this makes the chronology of the end-Triassic mass extinction incredibly similar to that of the Permo-Triassic—a double-phase extinction separated by a few hundred thousand years with the strength of each pulse variable from region to region.
Moving our attention to the open ocean, we find that the end-Triassic crisis was also manifest here. The radiolarians, which we last encountered struggling across the Permo-Triassic boundary, had rediversified and produced many new forms, including amusing hat-shaped species (hat-rads, if you like), and strange, fancy forms with twisted spines. All these types disappeared at the end of the Triassic to be replaced by radiolarians with much simpler, spherical morphologies in the Early Jurassic. Hat-rads lived in the surface waters and their demise indicates the severe nature of the stresses in the uppermost water column.
The radiolarian extinction had a brief but noticeable effect on ocean-floor sedimentation, which is now preserved in Japan. Red siliceous cherts made of radiolarian skeletons accumulated on ocean floors throughout the Triassic and Jurassic, but at the Triassic-Jurassic boundary this deposition was interrupted by a red mudstone that marks the temporary cessation of the rain of radiolarians to the ocean floor. If one looks down a microscope, the mudstone can be seen to contain fragments of volcanic glass and also pyrite—both components unique to this level. The glass suggests some far-distant volcanism (it is, of course, tempting to say this is a manifestation of the CAMP eruptions even though these were happening thousands of kilometers away from the middle of the Panthalassa Ocean). The pyrite suggests a brief pulse of ocean anoxia. Remarkably, it is also possible to infer climatic changes at this time even though the red mudstones were forming several kilometers below the ocean surface and thousands of kilometers from land. The evidence comes from subtle changes in the isotopic ratio of the trace metal osmium, which indicate a major but brief increase in runoff into the oceans at the end of the Triassic.
Other extinctions in the water column include the loss of the conodonts (a sad loss for paleontologists and geochemists, who love to analyze these fossils) and most of the ammonoids too. However, both of these groups appear to have been in decline throughout the Rhaetian Stage, suggesting that their final disappearance in the late Rhaetian was a coup de grâce for already impoverished groups. The real puzzle of the conodont and ammonoid decline is therefore, what was suppressing the usual evolutionary ebullience of these groups during the Rhaetian? Perhaps the Triassic-Jurassic extinction should be considered a protracted crisis? We have once again returned to the problem of not knowing the duration of the Rhaetian. If this stage only lasted a million years, then the entire extinction episode is brief, geologically speaking, but a 10-million-year Rhaetian implies an exceptionally long period of environmental stress. There is much work still to do to resolve these problems.
Ichthyosaur diversity also crashed in the later Triassic, although the precise timing is poorly resolved due to the rarity of fossils of these marine reptiles. Taking the fossil record at face value, it seems that many disappeared in the Norian Stage followed by the appearance of new types, such as Ichthyosaurus, in the Rhaetian. This hints that the Late Triassic crisis of water-column dwellers, such as the ichthyosaurs and ammonoids, was out of kilter with the other marine life. Overall though, for most groups the extinction was a late Rhaetian affair, but the larger water-column dwellers appear to have already suffered a (separate and earlier?) crisis by this time. In some regard, the end-Triassic mass extinction has parallels with the Permo-Triassic crisis, when nektonic predators were among the relatively few groups to be unaffected.
When attempting to understand the end-Triassic mass extinction, we are faced with explaining many of the same attributes of earlier mass extinctions. This was an environmental disaster bad enough to kill vast amounts of life everywhere: the dominant animals on land were wiped out, forests disappeared, and the tiniest plankton in the oceans suffered greatly, and yet intriguingly it afflicted other groups less harshly. We also have the same potential culprit—a series of giant flood basalt flows. Can we therefore cut and paste the same extinction cascade (fig. 3.4) from the older to this younger event? The answer is yes, sort of.
First, how does the timing of the eruptions compare with the timing of the extinctions? Much effort has been expended on this question, and the answer depends on how the volcanism is dated. One approach is to directly date the lavas using tiny zircon crystals because they contain significant amounts of uranium that allow the use of uranium-lead dating methods. This approach reveals that the initial eruptions began in Morocco and rapidly expanded (probably within less than 20,000 years) to cover the Newark Basin of the eastern United States. This huge burst of eruptions happened at the same time as the fern spike; we could not ask for better evidence of synchrony between volcanism and extinction. However, it is a bit more complicated than that. There is other indirect evidence to suggest that eruptions may have begun earlier. Large-scale volcanism affects the trace-metal content of the oceans. Oceanic osmium is particularly susceptible to volcanic influence because the isotopic ratio of osmium atoms in the mantle is different from that in the crust. By interpreting a change in the osmium content of seawater (recorded in marine Tethyan limestones), Andrea Marzoli has suggested that CAMP volcanism began at the start of the Rhaetian Stage. A similar change is seen in the contemporary deep-sea cherts of Japan, giving further credence to Marzoli’s notion. This would make the Moroccan lavas not the oldest of the CAMP but just the oldest that have been sampled so far. Some fragments of the LIP, such as little-known remnants of lava fields in the Amazon region of Brazil, could be older.
We therefore have two possibilities regarding the role of volcanism in the latest Triassic extinctions. Initial eruptions may have begun at the start of the Rhaetian (when pelagic groups such as ichthyosaurs and ammonoids began to suffer and maybe the crurotarsans were wiped out on land) followed in the late Rhaetian by a climax of eruptions and a major extinction on land and sea. Alternatively, eruptions may have begun in the late Rhaetian, and the earlier extinctions are either just misdated (e.g., the crurotarsans) or unrelated to volcanism (e.g., the ichthyosaurs and ammonoids). At present we cannot decide between these scenarios, although volcanism is accorded the main killers’ role in both.
As for the case of the Permo-Triassic mass extinction, the principal candidates for end-Triassic kill mechanisms are the gas effusions from basalt flows. The volumes of carbon dioxide release were probably impressive, as they were with the Siberian Traps, but it is unclear if these alone were sufficient to cause devastating warming or whether additional greenhouse gases were needed. The negative carbon isotope spike, noted above, could be a sign of methane release from gas hydrates (as has been argued for the same trend seen earlier). Thus, a warming pulse triggered by volcanic CO2 may have caused the release of methane, an effective greenhouse gas, and thereby exacerbated the temperature increase—a perfect positive feedback. Further stimulus may have come from the release of carbon dioxide from sediment baking beneath the CAMP lava pile (Svensen’s hypothesis again; see p. 34). As discussed above, there is certainly ample evidence from leaf stomata for this warming. Changes in vegetation recorded by sporomorphs also support a transition to warmer conditions.
Estimates of past temperatures can also come from fossil soil horizons,16 and studies from the Newark and Hartford Basins have provided some intriguing resolutions to the Triassic-Jurassic boundary warm phase. Rather than being a single warm interval, it has been resolved into a series of rapid warming-gradual cooling oscillations, each lasting a few hundred thousand years. What is going on? The answer probably lies in the nature of lava flows and their tendency to weather. The latter is immediately apparent to anyone who visits modern lava fields, such as those in Hawaii, where even flows that erupted only a few hundred years ago have soil developing on their upper surface. The weathering of fresh volcanic rock consumes a lot of atmospheric carbon dioxide (which ends up as bicarbonate washed into the ocean), and so eruptions are in a sense self-limiting: they put carbon dioxide into the atmosphere, but they also take it out again as the lava weathers. There is a lag, though, because the chemical breakdown of lava takes tens to hundreds of thousands of years. Hence, the warming-cooling oscillations at the Triassic-Jurassic boundary probably reflect episodes of eruption followed by quiescence and weathering. Such cycles only occur if the lava flows are erupted on land, as happened with the CAMP basalts, and in distinct pulses. There is no such feedback if lava flows follow each other quickly, because they become protected from weathering by the next lava blanket, or they erupt beneath the sea, where they get buried in sediment. This last scenario appears to have been the case with the Siberian eruptions, which occurred at sea level, and may partly explain the prolonged, continuous, and catastrophic warming trend at this earlier time.
Was warming in any way directly responsible for the extinctions? Jenny McElwain has certainly proposed this, but among the majority of Triassic-Jurassic cognoscenti, the favored cause of marine death is ocean acidification. Anoxia is also accorded a role in some extinction models although it is usually accorded secondary status in kill lists after acidification. This double whammy is also popular for the Permo-Triassic mass extinctions, but as we saw earlier, the preferred order is reversed—anoxia is clearly implicated whereas acidification is less certain. Michael Hautmann (University of Zurich) was the first to propose acidification as the end-Triassic killer. He noted that in many Triassic-Jurassic boundary records from around the world, limestone beds temporarily disappeared and were replaced by shales, and he interpreted this change to have been caused by intense ocean acidity. This transition is not seen everywhere though. In Britain the extinction occurs at the transition from black shales to the white limestones of the Langport Member. Perhaps the unusual chemistry of the isolated Rhaetian Sea in this region ensured that it was somehow protected from changes in the ocean pH?
The extinctions among bivalves have also been used to support an acidification scenario. Bivalves construct their shells using either aragonite or calcite, and Hautmann observed that the species with the more easily dissolved aragonite shells suffered a little bit more during the end-Triassic crisis. But, as noted in chapter 3, the rate of dissolution of both shell types is unlikely to be a serious issue in the lifetime of the bivalves.
Acidification and anoxia cannot be the whole story of the end-Triassic crisis. The hat-shaped radiolarians are unlikely to have been affected by either of these environmental factors—their silica skeletons would not been damaged by increased acidity, and the ocean’s surface waters where they lived are unlikely to have suffered oxygen deprivation. Also, for life on land, something else must have done the damage. The nature of the abrupt extinction, with a short-lived fern spike, suggests a short-term crisis. Intense warming caused by peak volcanism would fit the bill, as would ozone depletion. As with the Siberian Traps, CAMP volcanism at its peak may have been substantial enough to release devastating concentrations of ozone-destroying halogen gases. Large-scale volcanism is also capable of generating very short cooling episodes caused by sulfate aerosol formation. Perhaps a brief but severe pulse of cooling, lasting only a decade or so (the sort of duration of volcanic cooling episodes), contributed to the extinctions? There is, however, no geological evidence to support this idea; all the data suggest rapid warming during the extinction. For now though, we cannot say for certain which of the volcanic gases—halogens or sulfur dioxide—were the deadliest for terrestrial life, only that they acted quickly.
Before leaving the end-Triassic crisis, it is worth mentioning the possible role of meteorite impact because there is some intriguing evidence. It may be recalled that the Rhaetian sediments of Britain have a seismite bed found immediately above the extinction horizon (see fig. 5.2). This bed can be traced across hundreds of kilometers, implying a huge earthquake that may have been, just possibly, caused by an impact. As it happens, a crater of approximately the right age has now been found near the town of Rochechouart in the Limousin region of western France. The crater has been much changed by subsequent tectonic movements, but its original diameter is estimated at around 40 kilometers. Sadly for proponents who like the death-by-meteorite-impact scenario, this crater size is not especially big. In fact, quite a lot of craters of this size are known. The Rochechouart impact is therefore unlikely to have caused global environmental catastrophe, as shown by comparison with other impact craters. For comparison, the Chicxulub crater associated with the dinosaurs’ extinction is nearly 200 kilometers in diameter, indicating the size of impact required to be implicated in a mass extinction. Aside from Chicxulub, the largest meteorite impact crater is to be found in the Manicouagan region, in Quebec, and measures nearly 100 kilometers in diameter. At one time tentatively linked with the end-Triassic mass extinction, the Manicouagan crater is now known to have formed around 15 million years before the end-Triassic extinction, within the Norian Stage, a time noted for its lack of extinctions. The message here is that the threshold for lethal crater size needs to be in excess of 100 kilometers.
The Rochechouart impact is an unlikely cause of mass extinction, and it may not even have generated the British seismites. The onset of CAMP volcanism could have generated numerous violent earthquakes and shaken the seafloor sediments of the Rhaetian Sea. It thus seems more plausible that the seismic events record CAMP volcanism—the true killer of the Late Triassic. The consequence of these eruptions, like the Emeishan, Siberian, and Wrangelian eruptions before them, was a disaster for life and the environment. Recovery began immediately, as the dinosaurs and ammonites dusted themselves off and got on with business of taking over the world. But the bad times were not over yet. There was still another flood basalt province and another extinction to come.