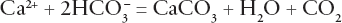
PANGEA’S DEATH AND THE RISE OF RESILIENCE
Around 135 million years ago, giant cracks started to open up among the sand dunes found in a vast desert that lay at the heart of Gondwana. Soon magma welled up through these fissures, and the dunes were drowned in flows of basalt lava hundreds of meters thick. The lavas continued to pour out for hundreds of thousand years, ultimately to form one of the largest of all LIPs—the Paraná-Etendeka Province. Today these lavas are separated by the South Atlantic, with the larger part occurring in South America (Brazil and Argentina), where the Paraná River cuts through them, while a somewhat smaller area is found in the Etendeka Desert of Namibia. Like the earlier CAMP and Karoo-Ferrar provinces, this latest LIP marked the start of continental breakup that was, in this case, to lead to the separation of the South American and African continents. But in the context of this book, the key question is, did the eruptions lead to mass extinction and catastrophic environmental change like the preceding five LIPs of Pangea?
The answer is an emphatic and simple no. The Paraná-Etendeka eruptions occurred during the middle of the Hauterivian, the third stage of the Early Cretaceous, when global extinction rates were extremely low. For lovers of catastrophes, the interval is a distinctly dull one. Perhaps the only interesting event was the temporary loss of shallow-water limestones in the western Tethys. They were replaced with shales as the seas deepened. Despite this regional pause in carbonate formation, overall it was a good time for limestone because planktonic forams began to radiate and their calcite shells had begun to rain down on the seabed in huge numbers. Hitherto, forams had only lived on the seafloor, where they were (and still are) very successful despite the various extinction crises they suffered during Pangean time. Around the middle of the Jurassic, some of them began to spend their lives drifting in the surface waters of the oceans, but it was not until the Early Cretaceous that this mode of life became common.
Planktonic forams were not the only calcareous plankton in the Cretaceous oceans. The coccoliths, which we have already met, made their appearance in the Triassic. By the Cretaceous their range had expanded out from Tethyan shelf seas into the open ocean. By the Hauterivian the combined rain of coccolith and foram skeletons to the seabed was forming a new type of sediment—deep-water pelagic carbonate, which was composed entirely of these tiny microfossils. We will return to the significance of this lime blanket later.
By the Cretaceous it seems the LIPs had lost their teeth. Pangea had seen 80 million years of LIP-driven disasters, but the next 80 million years saw amazing, uninterrupted evolutionary success stories. Feathered dinosaurs began to fly, and recent spectacular finds from China have shown that by the Early Cretaceous this group, the birds, had already attained an impressive diversity of forms. Plants were even more productive during this interval. The Early Cretaceous saw the rapid rise of the flowering plants (angiosperms), which finally brought some new colors to the green terrestrial landscapes. The rise of the social insects, such as termites, ants, and wasps, at the same time may owe something to the ascent of the angiosperms. Meantime, in the oceans, the burgeoning of life was similarly unimpeded. Especially successful were the decapod crustaceans, which produced two new groups—the crabs and lobsters. Sea urchins also underwent some remarkable changes. Hitherto they had lived on the seafloor encased in an armored sphere with defensive spines. This type still thrives today, but in the later Jurassic some started to burrow deep into the sediment, producing highly modified forms known as heart urchins.
All this innovation and diversification could have been brought to an abrupt halt by the Paraná-Etendeka eruptions. Groups as diverse as the birds and heart urchins could have had a brief flowering and so be as obscure today as earlier mass extinction victims such as the megalodontids or crurotarsans of the Triassic. But this did not happen and there was no crisis.
The benign nature of the Early Cretaceous volcanism is starting to attract attention from geologists and may have something to do with where it occurred. Earlier Pangean LIPs are thought to have intruded crust full of “juicy” rocks, such as coals and black shales, that when baked by magma generated huge volumes of greenhouse gases. This is the Svensen Hypothesis, and as we have seen, it has been deployed as a partial explanation for all the crises of Pangea. The crust beneath the Paraná lavas, on the other hand, is old and dry; it consists of already metamorphosed rocks that will not release many gases if baked by magma.
Direct evidence for the gas-poor content of the crust beneath the Paraná-Etendeka lavas has recently come from studying the sulfur content of pyroxene crystals in basalt. Pyroxene is one of the most abundant crystals found in LIP igneous rocks, and although no sulfur is found within its crystal lattice, it is possible for tiny inclusions or impurities to be trapped within the crystal. Analyzing them provides the chemical composition of the magma from which the pyroxenes crystallized. Thus, the Paraná-Etendeka magmas are thought to have contained around 800 ppm sulfur, whereas the CAMP magmas had over three times as much—1900 ppm. Perhaps this was the reason for the failed environmental response? There was simply not enough gas released to do any harm. This interesting notion is testable (and can be shown to be wrong) because the Paraná-Etendeka eruptions were just the first of a whole series of LIPs that formed since the start of the Cretaceous. Some of them passed through some very gas-rich crust indeed, especially the North Atlantic Igneous Province, which erupted 75 million years later. The effects of these younger eruptions are described further below, but they are not linked with a mass extinction.
LIPS AND LIFE
After the Jurassic Period, it becomes a lot easier to study the past environmental history of Earth because a lot of ocean crust is preserved from this time onward, whereas all the older crust has been lost through subduction back into the mantle. Remember that in previous chapters we had to look at much-deformed ocean-floor sediment in places like Japan to piece together what happened in the Panthalassa Ocean—a very imperfect and fragmentary record analogous to a jigsaw puzzle with most of the pieces missing. In contrast, the Cretaceous ocean floor puzzle is mostly complete because a lot of it is still at the bottom of the ocean, and numerous international ocean drilling programs have provided many cores through it. One noteworthy feature of Cretaceous oceans is that there are several giant plateaus. They are composed of sheets of flood basalt that formed from plume eruptions, just like the continental flood basalt provinces, but this time under the ocean. We have already met one example, the Wrangelia Province of western Canada, which formed within the Panathalassa Ocean at the time of the Carnian crisis, although it is now part of the Canadian Rockies (see chapter 4). Several of the Cretaceous ocean plateau eruptions coincide with interesting environmental changes that typically include such familiar themes as global warming and ocean anoxia.
The Ontong Java Plateau, which is found in the southwestern Pacific, is the largest of all the oceanic flood basalt regions. It is not especially well dated—because it is largely inaccessible beneath the waters of the Pacific—but it probably formed some time between 125 and 118 million years ago. Within this interval there is an interesting, short-lived oceanic phenomenon called the Selli Event, which happened 123 million years ago. It is marked by the spread of black shales in the deeper parts of the Pacific and Atlantic Oceans, a big change in carbon isotope ratios, and a warmer climate. All of this sounds very familiar. The changes bear close comparison with some of those seen during the Pangean crises, especially the Toarcian extinction. However, the Selli Event did not coincide with increased extinction rates, although a few groups show some interesting changes. One such is the rudists, a family of large thick-shelled bivalves similar to the megalodontids of the Triassic and the lithiotids of the Jurassic, both of which succumbed during Pangean extinctions. Many rudists disappeared from the fossil record during the Selli Event, but only temporarily, because they reappear after the event. The Selli Event was thus an interval of interesting environmental changes that never became a crisis for life.
The next great oceanic anoxic event, 29 million years later, was a similarly fascinating but relatively harmless episode. Called the Bonarelli Event, it too coincided with a hot climate, carbon isotope changes, and the eruption of oceanic flood basalts, this time in the Caribbean and Columbian regions. There were a few extinctions, including some planktonic foram species, with the result that a mass extinction epithet is sometimes used for this interval,19 but by the standards of Pangean disasters, it was a very minor crisis.
The possible standout exception to the harmless LIPs of the past 135 million years is the Deccan Traps—the product of a vast outpouring of flood basalts in western India 66 million years ago. They famously erupted at the moment that dinosaurs and numerous other groups, including the ammonites, marine reptiles, and nearly all species of calcareous plankton, went extinct. This is known as the Cretaceous-Tertiary mass extinction, the subject of some of the most vociferous scientific arguments of the past forty years because, as surely everyone knows, the extinction also coincided with a giant meteorite impact at Chicxulub. So we have choices: either the meteorite did it or the lava did it or it was some combination of these two. Many books have been written about the Cretaceous-Tertiary extinction and what caused it, but this is not the place for an exhaustive analysis. Fortunately though, the huge efforts that have gone into the study of the disaster provide a fantastic temporal resolution, especially the climate changes leading up to the mass extinction, and they repay some consideration.
In the million or so years prior to the Cretaceous-Tertiary mass extinction, there was a long-term cooling trend that sharply reversed to warmer conditions around half a million years before the end of the Cretaceous. The warm interval was then halted by a sharp cooling about 50,000 years before the end of the Cretaceous. Analysis of fossil plants in North America suggests that the cooling was substantial, maybe as much as 8°C, and was followed by stable temperatures across the boundary itself. So how does this compare with the eruption history of the Deccan Traps? The eruptions began about half a million years before the Cretaceous-Tertiary boundary, thus suggesting that the LIP may have initiated a warming climate—a link also seen during the Pangean LIP eruptions. But what about the subsequent cooling trend? Was it triggered by an especially explosive phase of volcanism injecting volcanic gases high into the stratosphere? Alternatively, did all the freshly erupted Deccan lavas start weathering intensely, thereby removing carbon dioxide from the atmosphere and cooling the planet? It could be either of these alternatives, and both have been suggested as explanations, but what is clear is that these potentially Deccan-driven climate changes predate the mass extinction at the boundary. The precise temporal link between meteorite impact and extinctions has always been the strongest argument in the debate on the cause of the mass extinction. Nonetheless, the debate rumbles on and there are those who claim the impact at Chicxulub was thousands of years before the mass extinction.
Latest Cretaceous extinctions are known, especially among North American dinosaurs, and they are often used as evidence that the world was in (volcanic) trouble even before the meteorite struck. However, it is also possible that the dinosaur losses are a regional climate signal. Remember, there was a transition from warm to cool conditions 50,000 years before the end of the Cretaceous. Diversity declines with temperature, as is immediately apparent if you consider the diversity of life in the Amazon compared to the Arctic tundra. Thus a cooling trend will see the loss of diverse, warmth-loving communities and replacement with less diverse, cold-loving communities. This local decline in diversity does not have to mean that there was a change of global diversity. If cooling was the cause for North America diversity decline, then it becomes hard to argue that the onset of the Deccan eruptions caused any significant extinction.
Disentangling the role of the Deccan eruptions and the meteorite impact in the Cretaceous-Tertiary mass extinction is clearly tricky, but fortunately the next LIP eruption presents a much “cleaner” story uncomplicated by a coincidental meteorite impact. This eruption occurred only 6 million years after the dinosaurs’ demise, and it triggered the onset of rifting between the North America and European-Asian continents. It was, in a sense, the final fragmentation of Pangea. Today the lavas are split between Greenland and the British Isles, with Iceland recording the remnant volcanism of the plume that fed the lavas. The province is termed the North Atlantic Igneous Province (NAIP), and the volume of lava extruded was truly huge even by LIP standards, probably more than 6 million cubic kilometers. In addition, the magma passed through some very organic-rich sedimentary rocks, and it seems likely that vast volumes of gas were emitted from baked sediments together with the volcanic gases. Indeed, Henrik Svensen devised his original hypothesis—of enormous thermogenic gas release from LIPs—based on a study of NAIP lavas in the North Sea.
Giant amounts of volcanism and giant gas release—if anything could replicate a Pangean LIP catastrophe, then the NAIP is clearly the best candidate to do so. So did it? The answer depends on whom you read, but there were certainly some fascinating environmental changes around the time of eruption, most significantly 70,000 years of intense global warming. That period occurred at the Paleocene-Eocene boundary, which has led the climatic episode to be called the Paleocene-Eocene thermal maximum (PETM). It coincides with a loss of ventilation in the deep-ocean waters of the Atlantic, where oxygen-poor conditions developed, and a sharp shift in carbon isotope ratios that become 3 ‰ lighter. Once again we have the ingredients of a Pangean-style mass extinction, and so we might expect to encounter one at this time. Many have indeed argued that this was the case; for example, one such doom-laden analysis in a popular science magazine proclaimed that during the PETM
the earth warmed faster than at almost any other time in its history. The average temperature soared by 9 degrees Fahrenheit, entire ecosystems shifted to higher latitudes, and massive extinctions occurred on land and, most telling, at sea. James Zachos, a paleoceanographer from the University of California at Santa Cruz [says] bottom-dwelling creatures with shells disappeared from the fossil record … [and] it took 60,000 years before sediments again began to show a thick white streak of fossilized shells.20
This sounds truly terrible, almost as bad as the Permo-Triassic mass extinction; however, the assessment of the severity of the crisis is untrue. Extinction losses were very selective and very minor during the PETM. Some deep-sea forams and ostracods in the Atlantic Ocean went extinct, probably because bottom waters became oxygen poor, while a few larger forams in tropical shelf seas also disappeared, but on the whole, this was a time of evolutionary success for a broad range of animals. Mammals did especially well. Many modern families appeared, including our own group, the primates, together with the first members of the deer and horse families. Interestingly though, most mammals were exceptionally small during the PETM. I suspect this was Bergman’s rule in action (see p. 98), according to which high temperatures favor small organisms.
Part of the problem for PETM scientists who claim that there was a crisis is that they are dealing with a spectacular climate event, in particular, a story of rapid warming that was probably driven by the release of greenhouse gases, and so an extinction is anticipated (just as a pending mass extinction is predicted for modern greenhouse warming). And yet, the PETM crisis was only the faintest echo of the Pangean extinctions. Claims to the contrary can be dismissed as mass-extinction envy on the part of those who study the PETM. What actually happened at this time is much more subtle and nuanced.
The best evidence for oceanic changes during the PETM comes from calcareous plankton. Samantha Gibbs (Southampton University) and Heather Stoll (Oviedo University) have pieced together the story and shown that the onset of the PETM is marked by the loss of pelagic limestones in some parts of the deep ocean. This loss does not reflect the extinction of the coccoliths and planktonic forams that form such sediments, because their diversity and abundance in surface waters remained unchanged, but rather the dissolution of the deep-sea carbonate sediment. The cause appears to have been extra organic matter raining down to the seabed, where it decomposed and acidified the sediment. The reason for the extra organics relates to the feedback mechanisms of the ocean-atmosphere system. Global warming increases rainfall, which flushes more nutrients into the ocean, thus stimulating more organic-matter production by plankton. This is the same old familiar process thought to have happened during the Toarcian and Permo-Triassic crises except that this time, during the PETM, there was no long-term effect.
Gibbs and Stoll also showed that the coccoliths responded in a perhaps surprising way during the PETM. Global warming driven by increased atmospheric CO2 will also see a decrease in pH as more of this gas dissolves in the surface waters. This acidification should be to the detriment of coccolithophorids with their calcitic shell, but in fact they thrive better because, despite the subtle reduction in pH, the extra CO2 stimulates more photosynthesis. This idea is supported by observations on modern coccolithophorids, which are found to grow thicker skeletons as carbon dioxide levels increase in sunlit waters. In essence, coccoliths provide a responsive feedback system for increased atmospheric CO2 levels because they help mop up the extra carbon. Ultimately this ends up either as limestones on the seafloor or, in the deepest parts of the ocean where coccoliths dissolve, their carbon becomes part of the ocean’s dissolved carbon content—another key process in lessening the effects of acidification.
So, is coccolith formation the key, new factor in helping moderate the effects of rapid carbon dioxide increases? Consider the following reaction for limestone formation:
This shows that the formation of the calcium carbonate also produces two by-products—water and, problematically, carbon dioxide. Ostensibly this should exacerbate the amount of CO2 in the ocean. However, two biocarbonate ions are used on the other side of the equation, and these are ultimately derived from rock weathering. Acid rain falling on the land surface attacks rocks and produces the bicarbonate ion, which gets flushed into the oceans. The ions increase ocean alkalinity and so help more carbonate to be precipitated. Overall, the two molecules of carbon in bicarbonate (which was originally found in two molecules of atmospheric carbon dioxide) are used up but only one is re-released as carbon dioxide.
The only problem is that the weathering process responds on a time scale of thousands of years, whereas a sudden increase of carbon dioxide emissions from, say, volcanism may occur in decades. So atmospheric carbon dioxide, along with surface water acidity, can increase quickly, whereas responses such as increased weathering and carbonate formation take thousands of years. The dissolution of deep-sea carbonate is another effective way of decreasing ocean acidity but, again, this is also a relatively slow process, because it takes hundreds to a few thousand years for ocean currents to mix the surface waters into the deeper parts of the ocean. These problems are of course at the nub of modern concerns of human-driven atmospheric CO2 increases. Ultimately, much of the carbon dioxide we put into the atmosphere will end up in the ocean either as organic carbon, carbonate, or dissolved carbon compounds; it is just that effects like surface water acidification occur very quickly whereas the negative feedback mechanisms are a little bit slower. Fortunately, coccolithophorids help matters considerably because rather than release carbon dioxide during shell formation, it gets used during photosynthesis.
If coccoliths are the key factor in the improved response of earth’s system to carbon dioxide increases, then the link between Pangea and LIP-extinctions may be entirely coincidental. The burgeoning of calcareous plankton and deep-sea limestone deposition may be the salient issue, not the absence of a supercontinent. This notion can be tested (sort of), by a quick survey of the extinction crises that occurred before Pangea assembled and before the rise of coccoliths. We need to look at the extinctions of the pre-Permian world.
A QUICK LOOK BACK
There was no shortage of extinction crises before Pangea although none were on the scale of the Permo-Triassic and end-Triassic cataclysms. Following the initial dramatic radiation of life, during the Cambrian explosion, the next 200 million years saw nearly a dozen severe crises, of which two merit the “mass extinction” epithet. Of these dozen, none are clearly linked with LIP eruptions although two may be. This weak link may reflect a problem with studying such deep time: these extinctions occurred a very long time ago, and many LIPs from this interval have since been eroded. Often only the deep “plumbing system”—the giant dikes and sills that sourced the magma—remain, whereas the surface lavas have been stripped off. This makes it hard to work out the original size of the volcanic provinces, and their ages also become unclear because the igneous rocks of the plumbing system need not be exactly the same age as the erupted lavas. Herein lies another problem: dating igneous rocks gets less precise the further back in time you travel. Thus, eruption ages for lavas from more than 500 million years ago may come with errors of plus or minus 10 million years.
One of the strongest connections between pre-Pangean extinction and LIPs comes from the Cambrian Period. The Kalkarindji Province, a much eroded but probably originally, large flood basalt region in northern Australia, was erupted around 506 million years ago. This about the same age as the first mass extinction. The principal victims were the archaeocyathids, a group of heavily calcified sponges that constructed large, highly diverse reefs in the Early Cambrian. They disappeared abruptly just prior to the Middle Cambrian, and several geologists have blamed the Kalkarindji eruptions for their demise. Not long after this extinction, there was another one, and this time it eliminated the redlichiids. These were the first major family of trilobites to appear, and they were also the first to disappear, suffering a mass extinction at the boundary of the Early and Middle Cambrian. A double-punch mass extinction like this is reminiscent of both the Permo-Triassic and Early Jurassic crises, and there are other similarities too—black shales became widespread around this time and the carbon isotopes changed. However, explaining how all these Cambrian phenomenon fit together is still at an early and rather speculative stage.
The other proposed LIP–mass extinction link of the pre-Pangean world occurred around 375 million years ago at the boundary between the Frasnian and Famennian Stages of the Devonian. This F-F event, as it is known, was severe, and it has been linked with a little-known LIP in Siberia (not the Siberian Traps, but a smaller one further east), but once again the age dating is only suggestive, not conclusive.
We are still a long way from making a connection between volcanism and extinctions in the pre-Pangean world. Potential research avenues could involve looking for tell-tale clues such as trace metal concentrations in sedimentary rocks. Giant volcanism releases a lot of metals to the atmosphere and oceans, and this volcanism proxy is potentially preserved in rocks. The search has yet to be made for such evidence.
In summary, life since the Cambrian has gone through four broad stages: an initial 180-million-year-long interval prone to mass extinctions and lasting from the Cambrian to the Devonian. It was followed by a more pleasant 100-million-year interlude lacking mass extinctions (and LIPs), during the Carboniferous-Middle Permian interval. Then came Pangea and 80 million years of volcanic hell. Finally, Pangea broke up, and it has been 180 million years of smooth sailing ever since, despite numerous LIP eruptions and one terrible meteorite impact. The discussion of the first two stages requires another book, but I would briefly note that the first stage lacked terrestrial vegetation; the first forests did not appear until toward the end of the Devonian, as the final extinctions struck. There may be a link between these observations.
RESILIENT PLANET
The key point of this book has to been to ask, why, during the reign of Pangea, were LIPs and mass extinctions occurring in lockstep, whereas the connection has been broken since Pangea disintegrated? This lack of extinctions is despite the observation that post-Pangea, LIPs still seem capable of causing interesting and significant global environmental change, especially global warming and ocean deoxygenation. And yet, none have been capable of causing major life crises. Why have LIPs and coincidental environmental changes lost their lethality?
To answer the question requires a return to the issue of how the earth’s ocean-climate system operates and specifically how greenhouse gases are removed from the atmosphere. This is the bread-and-butter of climate modelers such as Yannick Donnadieu of the Laboratoire des Sciences du Climat et de l’Environnement. Using the GEOCLIM modeling program, he showed that the Pangean world was generally very hot and arid with atmospheric CO2 values typically around 3000 ppm. By the Early Cretaceous, the world was much more efficient at removing CO2, with the result that atmospheric concentrations were maintained at an equilibrium value that was only a tenth of the Pangean level. The reason is related to the scale of Pangea. A huge continent has vast areas in the interior that are too far away from the sea to receive much rain. In contrast, smaller, more fragmented continents receive precipitation over a greater area. The crucial point here is that rainfall causes weathering, which consumes atmospheric CO2 that is then converted to the biocarbonate ion 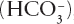 , which runs off into the ocean, increasing ocean alkalinity and making it easier to precipitate limestone. Continental runoff also supplies nutrients to the oceans, which stimulate plankton growth that removes more carbon dioxide, which gets buried as organic carbon in marine sedimentary rocks.
, which runs off into the ocean, increasing ocean alkalinity and making it easier to precipitate limestone. Continental runoff also supplies nutrients to the oceans, which stimulate plankton growth that removes more carbon dioxide, which gets buried as organic carbon in marine sedimentary rocks.
There are two further important factors that make a supercontinent poor at removing atmospheric carbon dioxide. Limestone deposition occurs in shallow shelf seas (because this is where the animals live that secrete the carbonate skeletons that form the limestones) and during Pangean times shelf area was at a minimum. This can be explained by simple observation: the shelf fringe of one supercontinent is a lot less than the shelf length of several smaller continents of overall equal area. Consider the following example. A circle of area 4 square centimeters has a circumference of 7.1 centimeters, whereas four circles with an area of 1 square centimeter have a combined circumference of more than 15 centimeters. Pangea was equivalent to a big circle, so there was not that much shelf space for limestone to accumulate; this way of burying carbon was not very effective. Limited shelf-sea space also ensures that the other main mechanism for carbon removal—organic-matter burial in marine sediment—was not working well either because most organic material is buried in shelf sediments.
Finally, as we have seen, the rise of coccoliths further increased the efficiency of the carbon dioxide drawdown pump. Importantly, since Pangea broke apart, calcareous plankton limestone deposition is no longer just restricted to shallow seas; it also accumulates in deep-water settings. There is thus an enormous increase in the potential area available for limestones to form, and a much more rapid response to changes in ocean pH is ensured. Alkalinity increases in the past caused by, for example, increased global humidity and enhanced runoff of bicarbonate-laden river waters could not be readily balanced by increase limestone formation, because limestones formed only in the limited areas available in shelf seas, and their extent depended upon sea level and the length of continental perimeters. Consequently, an increased rain of planktonic carbonate into the deep seas can counterbalance episodes of elevated alkalinity while elevated acidity can be buffered by dissolving the huge amounts of carbonate sediments in the deep sea, thereby neutralizing the pH decline. Deep-sea carbonate is a superb buffer of changes in ocean pH.
There is another newly evolved group of plankton that has greatly increased the efficiency of carbon burial in sediments. These are the diatoms, which appeared around the same time as the coccoliths, but only in the past few tens of millions of years have they become important. Diatoms are photosynthetic plankton that surround their organic cells with tiny skeletons of silica. This makes them much denser than other types of plankton that do not secrete a mineralized skeleton. Consequently, after their short life cycle in the surface waters, diatoms sink to the seabed quickly. This does not allow much time for their organic cells to decay in the water column, ensuring that more reach the sediment to be preserved and buried.
Terrestrial plants provide another important way of taking carbon dioxide out of the atmosphere (fig.7.1). Having been taken up by photosynthesis, the carbon can be buried in soil, stand around as the biomass of the plants, or return to the atmosphere as carbon dioxide from rotting vegetation. Around half of all the carbon buried in sediments today occurs on land, although the percentage varies enormously across geological time scales. But as a method of counterbalancing sudden increases in atmospheric CO2 levels, carbon burial is not very effective because the rate on land cannot be changed quickly.
During episodes of increased availability of atmospheric carbon dioxide, plant productivity increases, because CO2 is a valuable nutrient for plants, and forests become more luxuriant. So on a very short-term scale, CO2 is removed from the atmosphere and converted to plant matter. This is why modern tropical rainforests, like the Amazon, are so important to the health of the world—they are mopping up a lot of the carbon we are putting into the atmosphere; however, this effect is only modest, and it is only temporary because it does not increase the amount of carbon buried in sediment. The soils on which the Amazon forest grows are, for example, very poor in organic matter. Most plant matter just decomposes back to carbon dioxide on the forest floor.
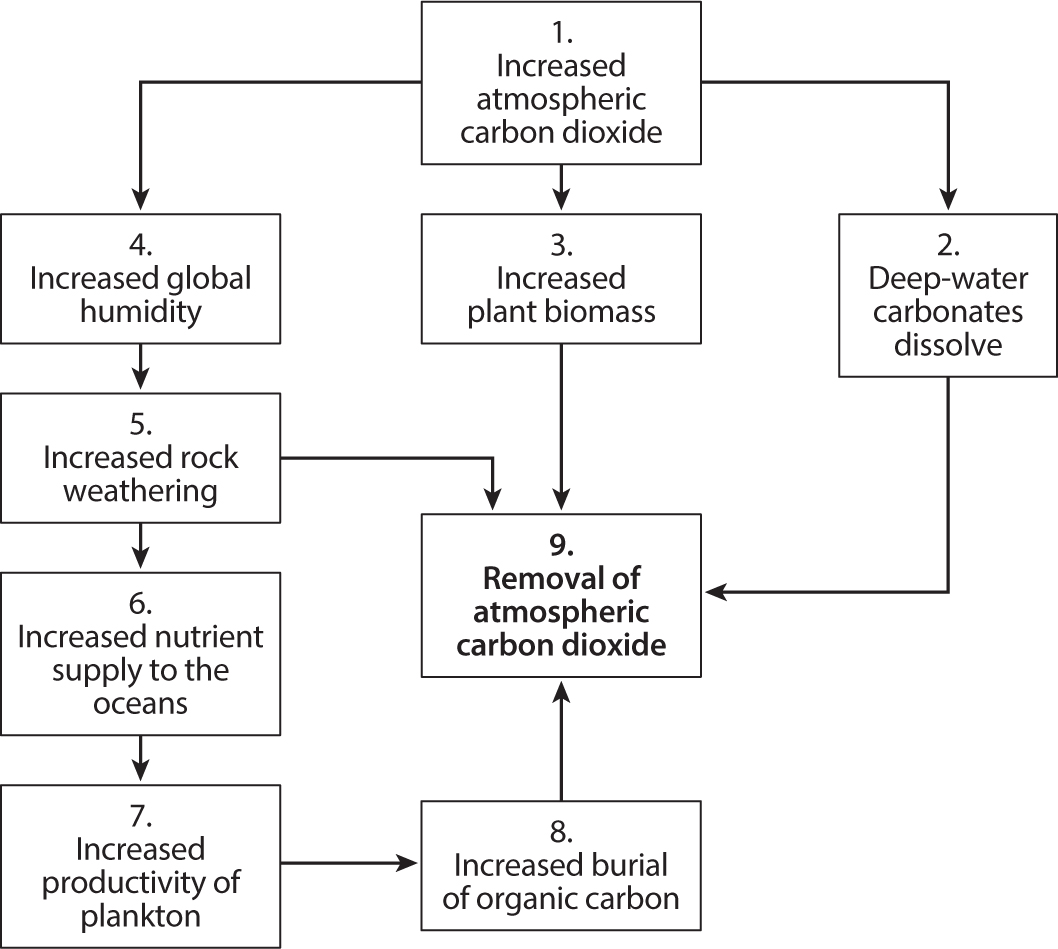
Figure 7.1. The carbon pump: how carbon dioxide is removed from the atmosphere by geological processes operating today.
To effectively bury terrestrial plant matter and so make an increased contribution to removal of atmospheric CO2 requires peat accumulation, which occurs in low-lying, rapidly subsiding swamps (where coals form) in a humid climate. The availability of such settings is controlled by geological processes such as basin formation and so has varied greatly through geological time in a manner unrelated to climate fluctuations. The key issue here is that subsidence is not controlled by climate. This situation is directly analogous to the process of limestone formation before the coccoliths appeared, because geological processes (like supercontinent break-up) control the area of shelf seas available for limestone formation, and this rock formation is not affected by short-term fluctuations in factors like atmosphere CO2 levels. Now that limestones also form in the deep sea, there is much less geological control on their formation but a much closer link to the climate.
Coal formation was extensive in the northern hemisphere during the Carboniferous Period, when low-lying swamps stretched across a vast distance from Pennsylvania through Europe to western Russia. In these regions, extensive equatorial basins proved to be the crucial factor for this coal-forming period, not the climate. In the Permian, such conditions were more prevalent in the high southern latitudes of Gondwana, and most coal formation was focused there at that time. In contrast, for much of the Cretaceous, climates were warm and humid—ideal for plant growth—and yet coal formation was less common at such times because most potential coal-forming basins were under water due to high sea levels. This illustrates the point that geology, not climate, controlled the amount of coal formation.
Burial of plant matter is therefore not intimately linked to the feedback mechanisms that govern the cycling of carbon between the atmosphere and the crust on short geological time scales. By this, I mean it is not very responsive to variations in atmospheric composition, although plants still play a role. Rock weathering is, for example, strongly controlled by organic acids generated by plant roots. Thus, increased humidity in a warmer world will favor plant growth, which in turn will favor accelerated rock weathering, which uses up atmospheric carbon dioxide. Even in the absence of plants, this weathering feedback process will still occur; it is just that plants speed it up. Thus, plants can be important and a world without plants would be much more prone to rapid climatic fluctuations. It was probably a significant factor following the mass extinction of plants at the end of the Permian too.
The efficiency and response time of carbon dioxide drawdown since the disintegration of Pangea has improved, with the result that sudden increases of atmospheric concentrations, such as seen at the onset of LIP eruptions, are no longer capable of causing catastrophes. There are now numerous ways in which the extra carbon in the atmosphere can be rapidly counterbalanced by extra carbon burial, and even in the short term, enhanced dissolution of deep-water carbonates is an effective buffer. Many of the improvements in the Earth’s feedback mechanism relate to the break-up of Pangea, such as the higher overall humidity of smaller continents and the extra area available for carbon burial in the fringing shelf seas of dispersed continents. In addition, the subsequent evolution of planktonic groups with hard parts has greatly speeded up the transfer of carbon to sediments. These changes seem sufficient to stop the runaway climatic effects that cause mass extinctions. In some cases, such as the impressively large NAIP eruptions 60 million years ago, the environmental and climatic effects travel sufficiently far down the road that they begin to resemble a mass extinction but do not go far enough to actually cause one. The warming, acidification, and deep-ocean anoxia never get to the point where they become widespread and/or lethal.
During Pangea’s existence, the effects of sudden climate warming became so severe that nasty feedback effects kicked in and magnified the consequences of warming to a degree that was catastrophic. At the greatest extreme, the failure to stem temperature rise during the Permo-Triassic crisis reached a point at which it became difficult to bury organic matter (because it decays quicker at higher temperatures). The result was temperatures too hot for terrestrial photosynthesis, which in turn retarded organic-matter formation and rock weathering, thereby weakening key parts of carbon cycle. Fortunately, the development of ocean anoxia meant that a new site of organic-matter burial appeared—the deep and cool abyssal depths—allowing normal conditions to be eventually reestablished. It was too late for most marine life, though, because by this time it had had to endure the double whammy of anoxia and high temperature.
The modern world’s feedback mechanisms may be likened to a six-cylinder car engine that runs smoothly, occasionally speeding up when more CO2 is injected but never misfiring (fig. 7.1). In contrast, Pangea’s engine was four-cylinder and sometimes misfired and ran on three. And on at least one occasion, one process—terrestrial plant growth and burial—effectively shut down.
We still do not have all the answers to the questions posed by Pangean mass extinctions. The devastation of land communities is especially hard to explain. The extinction of Late Permian terrestrial communities is a truly awesome phenomenon, which might be related to atmospheric changes such as ozone destruction. Massive volcanic halogen emissions provide one cause, but then why did this only happen during Pangea’s lifetime? More recent eruptions, such as those of the NAIP, would be expected to also emit huge amounts of halogens. Alternatively, some geologists have attempted to link changes in the oceans to events on land. Thus anoxic oceans may have leaked hydrogen sulfide into the atmosphere, where it would interfere with the formation of ozone. Unfortunately, this probably is not the answer. Hydrogen sulfide is immensely reactive with oxygen and is unlikely to ever reach the stratosphere and damage the ozone shield; instead, it will oxidize rapidly close to the sea surface. Perhaps terrestrial warming was the stress factor on land, but it would have been most harsh on forests adapted to living in cold conditions. The extinctions show no such temperature dependence—equatorial forests suffered as badly as those at high latitudes. And so the puzzle remains.
Clearly we do not yet have all the answers, but we have a come along way on the road to understanding the great mass extinctions of the fossil record.
This book is not intended as a warning from prehistory. If anything, its message has been the opposite—it was dangerous to live at a time of a supercontinent, whereas worlds like our own, with widely dispersed continents, are much better. They seem capable of absorbing the impact of sudden, dramatic increases of atmospheric carbon dioxide without any catastrophic change; surely an optimistic conclusion. Much of the “rapidity” of mass extinctions is in any case only rapid to a geologist—collapses in diversity typically happened in thousands of years—intervals that are almost beyond our concern. Would we worry if our actions today were storing up trouble for the world in 5000 or 10,000 years’ time? Nonetheless, LIP eruptions are the only natural process to provide an analogue for our current impact on the atmosphere, and some of the more recent examples appear capable of causing pretty spectacular climate changes, such as the intense warming episode 60 million years ago. Even if the extinction losses at that time were very modest, the effects were significant; most life on land had to shift its distribution poleward as climatic belts expanded away from the equator. This was at a time before man-made landscape change had broken up natural habitats into fragmented, undisturbed oases in a sea of farmland and urban sprawl. Modern life on land is going to have a hard time moving (hopping between fragments) to accommodate warming trends.
In the oceans, the clear story from past mass extinctions is that global warming goes hand in hand with oxygen starvation and that both are very bad for life. The modern oceans are showing the first signs of these changes, and modeling predictions suggest that we may only be a few hundred years away from seeing large expanses of anoxic waters develop. The effect on sea life will not be good, although by then massive overfishing will probably already have destroyed most ecosystems as we currently know them.
The ultimate take-home message of this book is that for 80 million years a series of gigantic volcanic eruptions caused a series of crises that fundamentally changed the course of life and removed groups that would otherwise have thrived. We should pity the animals of Pangea. For the pareiasaurs, dinocephalians, and crurotarsans, all looked to be going so well, but they had their reigns cut unexpectedly short. In recompense it allowed our favorite animals, the dinosaurs, to take center stage for a very long time indeed, but they were simply lucky that their reign coincided with the break-up of a lethally large supercontinent. In the end their luck ran out too.