Some of the adaptations that the Skhul-Qafzeh humans acquired to deal with Neanderthal predation were purely physical, involving modifications to the human body and its reproductive system. Others were behavioural. Some behavioural responses were obviously learned and passed on culturally from one generation to the next. But others appear to be innate or instinctive, and include fears, sexual preferences, attitudes and other emotions that, I argue, are now part of human nature. It is imperative that we understand how these genetically programmed emotions, innate behaviours and instincts were biologically encrypted into the Skhul-Qafzeh genome, so that thousands of years later
they are inherited by modern humans. And are still pulling our strings.
Ethology, a branch of zoology, is the study of genetically programmed behaviours known as instincts and innate behaviours in animals. In 1938, two of the founding fathers of ethology (and winners of the 1973 Nobel Prize) Konrad Lorenz and Nikolaas Tinbergen gave a name to the kind of innate responses that animals regularly display. They called them fixed action patterns of behaviour503—because when they are triggered by subliminally perceived stimuli, these behavioural responses tend to be rigid (or fixed).
In 1951, Tinbergen gave a name to the stimuli (or cues) that trigger fixed action patterns. He called them sign stimuli, and they included any sensory stimuli or cue which evoked an innate behavioural response.504
A sign stimulus is also sometimes called a releaser because it releases the innate behavioural response.
To date, sign stimuli and fixed action patterns of behaviour have been used almost exclusively in the study of nonhuman animal behaviour, but it is my intention here to apply ethological principles to recent human evolution. I take the view that there is essentially no difference in the way human and nonhuman animals acquire new innate behaviours and therefore ethological research on ducks, geese, and other animals can be applied to reveal the origins of human nature.
Borrowing the ethological terminology of Lorenz and Tinbergen and ap- plying it to NP theory suggests that modern humans have inherited a number of Neanderthal-specific fixed action patterns from their Levantine ancestors. These fixed behavioural responses lie dormant until subliminally triggered (or released) by a sign stimulus, which can be a physical feature, a behaviour or something in the environment that acts as a cue.
Sign stimuli can trigger either a positive or negative fixed action. For example, among Palaeolithic males, flat female chests would tend to produce negative responses, while large rounded breasts would produce positive responses. In actuality, small variations in each trait acquired considerable significance as humans became adept at differentiating subtle differences between Neanderthal and nonNeanderthal forms. My speculative list of physical traits that may act as Neanderthal sign stimuli includes:
face
prominent brow ridges and connected eyebrows; midfacial projection; lack of symmetry; thin lips—no lips; flat nose with forward facing nostrils; large bulbous eyes with slit pupils and brown scleras; absence of defined chin; wrinkled, leather-like skin with little elasticity.
body
hunched shoulders; thick neck; bow-legged; flat buttocks; thick body hair; ape-like gait; humpback
babies
minimal antipodal body fat; wrinkled skin.
behaviour
brutish demeanour; lack of aesthetic appreciation; grunting—restricted vocal range; limited emotional repertoire; unanimated facial expressions; sexual promiscuity; incest; incomprehensible language, alien speech; stupidity; no sense of humour; no personal hygiene; smelly, dirty; public defecation; lack of sexual modesty; flagrant display of genitalia; sniffing and playing with excrement; public sex and masturbation; incapacity to form lasting pair bonds or fall in love; rape; no interest in clothes as fashion; no decorative art; no interest or talent for music; dance or singing; no body ornamentation, tattooing, scarification, jewellery, etc.; sniffing female genitalia; sexual predation; ‘shadowing’ (stalking) females for sexual opportunities
environmental
darkness; moonless nights; sudden noises (especially at night); caves and other subterranean abodes; dogs barking at night, forests at night.
It is a major tenet of NP theory that these clandestine Neanderthal sign stimuli became the most important single mechanisms of postNeanderthal human evolution. But this brings us to the imperative question— how did the Skhul-Qafzeh humans genetically acquire their Neanderthal sign stimuli in the first place? That is to say, Skhul-Qafzeh sign stimuli are innate and inheritable, which means they must have been biologically encrypted in their DNA. So how did this extraordinary molecular encryption of the Skhul-Qafzeh genome take place? This is such a seminal question—it extends out like a great veil to encompass the most basic questions of life on earth. How do animals (including humans) acquire new instincts and innate behaviours? How does information from the external environment get coded into DNA?
The most obvious answer is that sign stimuli are derived from random mutations in the nucleotides of protein-coding genes, which become fixed because they confer a functional advantage. There are several major problems with this idea. Point mutations are random, essentially errors of DNA replication, and contain no coherent information in themselves. But sign stimuli are anything but random. They are external sensory cues, comprised of specific environmental information. Such non-random packages of coherent environmental information can have been acquired only from an individual’s environment. They are therefore external to the genome. The information contained in sign stimuli are often so complex, detailed and specific to the organism’s current environment that they could not have accrued by random means.
Secondly, point mutations only code for protein synthesis, which in turn builds physical traits—noses, cartilage, teeth, blood, capillaries and so on. No protein-coding gene for a complex behaviour or instinct has yet been discovered. Sign stimuli, on the other hand, are not physical. They are simply packages of ‘non-physical’ environmental information.
In biology, when information is acquired from the environment, the environment is said to instruct the genome, and this poses a problem for natural selection. This is because natural selection is not an instructionist process—it is a selectionist process—it just selects the most favourable random mutations that periodically occur. The distinction is important because the only way natural selection can function adaptively is if it prevents the environment instructing the genome with new physical traits. This is known as the central dogma of biology.
As formalised by Nobel Prize winner Francis Crick,505 the central dogma means that genetic information does not flow from the environment to the genome. In practical terms it means that physical traits acquired during the life of the individual (such as bigger muscles, suntans, and broken legs) cannot be passed on to the next generation via inheritance because the consequences would be catastrophic. It may seem adaptive for a blacksmith to inherit his father’s big muscles, (which he acquired through exercise and hard work) but that would mean he could also inherit his father’s angina and lumbago, acquired during the blacksmith’s life. In other words, while it might be advantageous to inherit certain acquired physical characteristics, ultimately, the risk of fatally contaminating the genome with acquired detritus means that instructionist inheritance (Lamarckian inheritance) is untenable.
The Lamarckian concept of acquired characteristics incorrectly supposes that by constantly stretching to reach higher branches, giraffes develop longer necks and these can be passed on to progeny.
And yet to survive, animals need instincts that are specifically tailored to their current environments. Animals need to identify the latest predators and prey. They need to know how to find their favourite food, how to mate and generally survive the specific (and often variable) conditions of their current environment. Birds need to inherit complex maps to locate breeding grounds thousands of miles away. These instincts are particularly important in species that cannot learn such skills from their parents, like fish, reptiles, insects and many animals. So, while the central dogma pre- vents the environment instructing the genome with physical traits, when it comes to behaviours—and, in particular, instinctive behaviours—then it is advantageous for the environment to instruct the genome with new environment-specific information and behaviours.
No-one was more aware of this paradox than Charles Darwin. On his voyage in HMS Beagle, Darwin observed dozens of examples where the environment appeared to be instructing animals with new innate behaviours. He was particularly struck by how new environmental threats and dangers were quickly incorporated into new instincts. For instance, Darwin observed that large (edible) birds that had no contact with people did not fear humans, but species of large birds that had been recently hunted by humans quickly developed an innate fear of people which their offspring inherited.
As a result of his observations, Darwin came to believe that a second evolutionary process (besides natural selection) was responsible for animals acquiring new environment-specific instincts which were then inherited by their offspring.506,507 Eventually, he developed an instructionist theory which he called pangenesis,508 based on French naturalist Jean-Baptiste Lamarck’s theory of acquired characteristics. Unfortunately pangenesis, like Lamarck’s own theory, was soon shown to be incorrect.509,510 In the absence of a new theory to explain how animals acquire environment-specific instincts, most evolutionary psychologists simply resort to Darwinian mutational theory.511 But despite enormous advances throughout the last century in every field of biology, 21st century scientists still did not understand the innate ability of new-born turkey chicks to recognise predatory hawks, nor how this amazing feat was first encoded into its genome.
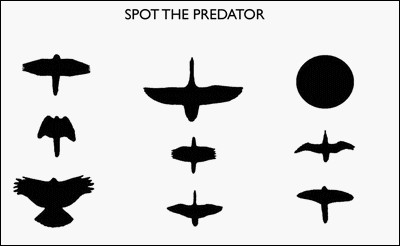
Using bird models flying along wires, ethologist Nikolass Tinbergen demonstrated that new born turkey chicks can recognise predatory birds from their shape. When they recognised a hawk, they instinctively ran for cover, but did not do so for other birds.
In 2005, I proposed a new nonLamarckian explanation for how the environment instructs the genome of multicellular animals with new instincts and innate behaviours.
My paper, ‘Noncoding DNA and the Teem Theory of Inheritance, Emotions and Innate Behaviour’512 and subsequent papers513,514 demonstrate that high intensity emotions (typically caused by predatory attacks, accidents and other traumatic misadventures) can be imprinted into an area of an animal’s DNA called nonprotein-coding DNA (ncDNA). This is the area of DNA that was often dismissed as ‘junk DNA’.
For example, if a turkey survives a hawk attack, its emotional response (terror, anguish etc.) can be encoded into its ncDNA and, under certain circumstances, these traumatic emotions can be inherited by its offspring.
When this molecular encoding occurs, it is inherited by progeny as a nebulous emotional memory. It is this emotional memory that future generations of turkey chicks use to identify dive-bombing hawks. Importantly though, the episodic memory of the experience is not inherited, nor are any physical consequences. If a boy is mauled by a dog, his lacerated wounds are not inherited, nor is the intellectual memory of the attack. Only the traumatic emotions are inherited. Because no physical effects are inherited, this evolutionary process (which I call teemosis) does not violate Crick’s central dogma.515
Fracturing genetic homeostasis and encoding an emotional memory into ncDNA requires intense emotions—those usually associated with traumatic life and death experiences—the kind of powerful, out-of-the- ordinary affective experiences that would never be forgotten. For example, a young Skhul-Qafzeh girl abducted by a group of Neanderthals and subjected to a prolonged sexual assault may have been so psychologically traumatised by the attack that she encoded a ‘rape teem’ through which she could pass on the experience to her descendants—as an emotional memory of the assault (but not an actual memory).
Because only the emotional memory of a traumatic experience is encoded into the genome, I call these packages of inherited emotion teems (Trauma Encoded Emotional Memory). Teems provide a nonLamarckian means by which the genomes of multicellular animals can be encoded with adaptive information from their environment.
The development of Teem theory involved the identification and systematic analysis of many human teems to determine the traumatic environmental circumstances that may have been responsible for encoding them. For example, if most humans instinctively react to something small, dark and furry crawling on their skin, one can conclude that this is a ‘spider teem’, encoded by a forebear’s harrowing encounter with an arachnid. Similarly, if you experience an anxiety attack while walking through a forest at night, it could suggest that a distant ancestor had been traumatised in a dark forest.
Species acquire many teems, but usually it is only the adaptive ones (those that contribute to survival) that are selected and passed on. For instance in 1960, Cornell University researcher Eleanor Gibson conducted a series of experiments that demonstrated that rats, kittens, lambs, puppies and human toddlers instinctively recognise a precipitous cliff and shy away from the edge.516,517
According to Teem theory, Gibson’s experiments demonstrated a simple ‘cliff teem’, encoded perhaps by a prehistoric infant who survived a traumatic fall from a great height and encoded a teem as a result. Because the teem provided the toddler’s ancestors with an innate aversive reaction to steep cliffs, it was highly adaptive and eventually became fixed.

An infant intuitively backs away from what it believes is a cliff. From M. W. Matlin and H. J. Foley (1997) Sensation and Perception (4th Edition) Allyn and Bacon. P448
Similarly, if humans around the world are innately concerned with body odour, it suggests that when this teem was first encoded, it provided an adaptive advantage and so was retained and spread to fixation. This kind of analysis led directly to the formation of NP theory.
To understand why modern humans still carry so many vestigial teemic responses to Neanderthal predation, we need only note that teems are encoded only by extremely intense emotions. And predation—for those that survive it—is all about trauma and near-death experiences. We know from field studies, for example, that predation can be so traumatic that survivors often die later of shock from non-lethal injuries. Eugène Marais, who studied free-ranging baboons in northern Transvaal in the
1930s, observed that constant predation by lions and leopards meant that “the life of the baboon is in fact one continual nightmare of anxiety.”518
While the physiological effects of predation stress on victims is well- known—including elevated levels of glucocorticoids,519 cardiovascular disease and damaged immune responses520—similar responses have also been detected in individuals who witness a close relative or companion being killed.521 Among modern humans, even bereavement caused by natural causes is known to increase morbidity and mortality,522 so we may expect the family and friends of Levantine humans taken by Neanderthals to be especially traumatised. This suggests it was not only the direct survivors of Neanderthal attacks that encoded Neanderthal teems but, by virtue of the generalised trauma of predation, their relatives and friends too.
Given this, and the fact that Neanderthal predation was consistently traumatic over so many millennia, it is hardly surprising that human nature is littered with Neanderthal teems. If, as my interpretation of the data suggests, every Neanderthal sign stimuli in the modern human genome was encrypted there (by the teemosis evolutionary process) as a consequence of an individual’s traumatic personal experience with Neanderthal predation, then it strengthens the view that our ancestors led much more traumatic and fearful lives than the fossil record indicates. In addition, Teem theory pinpoints human sexuality as having been subject to a traumatic upheaval at some stage in human evolution.
For a more detailed explanation of the Teem theory of behavioural evolution, see my forthcoming book The Second Evolution, or my website (www.thesecondevolution.com). But for the purposes of NP theory, by outlining the biological mechanism by which traumatic emotional encounters with Neanderthals could be encrypted into the human genome and inherited, Teem theory fills a gap in the Neanderthal predation paradigm. Importantly, it reveals that Neanderthal sign stimuli are derived directly from Neanderthal teems, and those teems in turn are derived from the traumatic personal interaction between Neanderthals and early humans.

Ancestral humans could identify Neanderthals using their Neanderthal teems. These emotional pictures were sufficiently nuanced to allow someone to identify a Neanderthal without ever having seen one before. The same teemic emotions guided our reconstruction of the La Ferrassie Neanderthal (above).