SELECT A SPECIES
I taught a class on the topic of de-extinction recently for graduate students studying ecology at UC Santa Cruz. For their first assignment, I asked each of the students to choose an extinct species that they’d like to see brought back to life and to become that species’ de-extinction advocate. I was curious to learn not only which species they would choose but also what their motivations for choosing a species would be. Because the students were ecologists, I expected them to focus on the impact of each potential “de-extinctee” on the environment into which it would be released, should de-extinction be successful. They did not.
The students selected, among other species, the Yangtze River dolphin, the dodo, the moa, the Tasmanian tiger, the Cascade Mountains wolf, Steller’s sea cow, and Thismia americana—a tiny, translucent plant that is so poorly described that it lacks a common name. Some of their arguments in favor of de-extinction were purely research-oriented—imagine what could be learned by studying this species—while others were more practical—imagine how this species might create new opportunities for ecotourism. Most students discussed the technical challenges of de-extinction—it would be hard to find well-preserved dodo remains, for example, or a surrogate mother for a Tasmanian tiger. Some students acknowledged that suitable habitat might be hard to find, and that existing laws might make it hard to protect the species once it was released into the wild. Few, however, discussed what effect introducing an unextinct species into an existing community might have, which surprised me.
As the class progressed, it became clear that the students had different motivations for selecting candidate species for de-extinction. Some students wanted to bring a species back simply because it would be exciting to do so. Others chose species that they believed could provide significant ecological and environmental benefits, or that might improve our understanding of the evolutionary processes that lead to diverse forms of life. One student selected the species that he believed had the fewest technical barriers in the way of success.
None of these are “wrong” reasons to choose a species for de-extinction. However, the diversity of motives within this small group highlights the first significant challenge faced by scientists doing on-the-ground de-extinction work: to agree on what to bring back. How do we decide which species should be the first targets of de-extinction? Should we choose the species that will be the easiest to bring back? The most awe-inspiring? The most likely to draw attention, perhaps motivating further investment into the technology? Or should we focus on those species whose de-extinction is clearly scientifically justifiable? And, if the latter, what does that mean exactly? Finally, and just as importantly, who is the “we” that gets to decide?
THE “RIGHT” REASONS FOR DE-EXTINCTION
As I suggest above, there are likely to be many reasons to select (or not to select) a particular species for de-extinction. Whether de-extinction is technically feasible, and whether suitable habitat exists into which a species might be reintroduced are important considerations. These questions address whether a species can be brought back, however, rather than whether it should be brought back. The latter is, unsurprisingly, a much more difficult question to answer.
Let’s consider the Yangtze River dolphin, for example. Bringing back the Yangtze River dolphin would certainly be exciting, which some may feel is sufficient motivation to try. Those who would benefit most from its de-extinction would probably be more likely to advocate for it over other candidate species. But who would those people be? The students offered three substantive arguments in favor of bringing back the Yangtze River dolphin, each highlighting different potential benefits, and therefore beneficiaries, of Yangtze River dolphin de-extinction.
The demise of the Yangtze River dolphin—also known as the baiji—is a terribly sad case. My friend Sam Turvey, who works for the Zoological Society of London, has devoted a big part of his life to looking out for species that are on the brink of extinction. In 2006, he led an expedition to survey the Yangtze River for any signs of river dolphins. Sam and his team searched the Yangtze River system for two months and saw no dolphins or signs of dolphin life. With heavy hearts, they declared the Yangtze River dolphin functionally extinct.
The first of the students’ arguments in support of Yangtze River dolphin de-extinction emphasized the evolutionary distinctiveness of the Yangtze River dolphin. Only two other freshwater dolphin species—the Ganges River dolphin of Southeast Asia and the Amazon River dolphin of South America—are known. When scientists first described the river dolphins, they noticed that the three species looked very much alike. All three river dolphins have long, narrow mouths, for example, with lots of teeth. They also have small eyes, compared with their marine relatives. Scientists decided that these morphological similarities probably meant that the three river dolphin species were descended from a single common ancestor species that was also a river dolphin. When genetic data became available, however, it was clear that this was not true. Instead of confirming a single evolutionary lineage, the genetic data indicated that each species made a separate transition from the ocean to freshwater. The morphological similarities among them arose from a combination of shared, deep ancestry and convergent evolution—life in similar environments led to the emergence of similar traits. This makes each freshwater dolphin species particularly valuable from a scientific perspective. We can compare their genomes, physiologies, and behaviors to better understand how species adapt to freshwater environments. Scientists would be one group to benefit from Yangtze River dolphin de-extinction.
The second of the students’ arguments pointed out that rare things intrigue everyone, and not only scientists. If the Yangtze River dolphin is brought back to life, the spectacle of its existence would likely be sufficient to attract certain types of tourists who would be keen to see the animal firsthand. Ecotourism is one of the most rapidly growing sectors of the tourism industry. It both provides jobs and inspires communities to take advantage of local natural resources. Tourists would come to take photos, sleep in a local hotel, eat at a few local restaurants, and maybe even buy a stuffed toy dolphin replica to take home. Yangtze River dolphin de-extinction would have a positive economic impact. The people who live in the region of reintroduction would benefit, as would tourists—some may even be inspired to care just a little bit more about the plight of the native species back home.
The students’ final argument postulated that Yangtze River dolphins should be brought back because their de-extinction would necessarily have a positive impact on the environment. The Yangtze River is presently too polluted to support dolphins, so this situation would have to change. Bringing back the river dolphin would require making the river ecosystem a cleaner, healthier one, with far-reaching ecological benefits.
This same multi-faceted rationale holds true for other species. For example, another group of animals that my students felt were good candidates for de-extinction were the moa of New Zealand. As with the Yangtze River dolphin, the reasons for bringing back the moa are both scientific—moa have no close living relatives, so understanding their biology and behavior is nearly impossible without bringing them back—and economic—living moa would provide yet another reason for people to visit New Zealand, which is already a popular ecotourism destination. Resurrected moa may also re-establish missing interactions with other species, benefiting the native ecosytems of New Zealand.
Moa were enormous birds that did not fly (figure 1). Some species of moa reached more than three meters tall with their necks outstretched and weighed more than two hundred kilograms. Because they didn’t fly, moa were easy targets for the first inhabitants of New Zealand—the Māori—who hunted them for food, used their bones to make jewelry and fishing gear, and fashioned their skins and feathers into clothing. Māori and moa coexisted on the islands of New Zealand for more than three hundred years before hunting and habitat loss eventually led to the moa’s extinction.
Figure 1. Sir Richard Owen and his reconstruction of a giant moa, Dinornis novazealandiae. In his right hand, Owen holds the first moa bone that he examined. This photograph was first published in Owen’s book, Memoirs on the Extinct Wingless Birds of New Zealand, vol. 2 (London: John van Voorst, 1879). Courtesy of the University of Texas Libraries, The University of Texas at Austin.
In New Zealand, moa are a symbol of national pride. Very briefly in the 1890s, New Zealand was officially dubbed the “Land of the Moa,” thanks in part to a play of the same name written by George Leitch. New Zealanders have created moa artwork, moa poetry, and even moa beer, and many New Zealanders are strongly in favor of bringing moa back to life. New Zealand has a strong record of environmentalism and protection of native species and habitat, which means that unextinct moa would probably be provided with a safe place to live should they be resurrected. However, some of the challenges of de-extinction mean that unextinct moa would probably not be 100 percent identical to the species that once inhabited New Zealand but rather genetic hybrids with nonnative birds. It is not clear how these hybrid animals might be accommodated within the environmental philosophies of many New Zealanders.
A third popular choice among my students was the dodo (figure 2). Dodos were large, flightless pigeons that were endemic to Mauritius, a volcanic island situated in the Indian Ocean about 1,200 miles from the southeastern coast of Africa. In 1507, Portuguese sailors landed on Mauritius, which was at that time uninhabited by people, after being blown off course by a cyclone. The Portuguese were not particularly interested in the island and did not establish a permanent colony. Dutch sailors arrived about ninety years later but didn’t stick around either. They did, however record for the first time a large, fat, flightless bird with little to no fear of humans. In 1638, twenty-five Dutch sailors returned to Mauritius and established the first permanent human settlement. Twenty-four years later, the dodo was extinct. Based on written accounts of the interactions between humans and dodos, humans are clearly to blame for the dodo’s extinction.
As with the Yangtze River dolphin and the moa, scientific, ecological, and economic interests can be cited as reasons to resurrect the dodo. The dodo is a large, flightless pigeon whose closest relative is a small, strong flyer. Studying its genome could help scientists to better understand how traits such as flightlessness and gigantism evolve. Reintroducing dodo populations to Mauritius would require the creation of suitable habitat, which would mean removing invasive species and establishing new protected zones, which would benefit both the local people and the native ecosystem. The dodo is a special case for de-extinction, however, because it, more than any other species, is the international symbol of human-caused extinctions. If candidate species were to be ranked according to the potential psychological impact of their de-extinction, the dodo would be very high on the list.
A SHORT GUIDE TO DE-EXTINCTION DECISION MAKING
The three examples above highlight what I have come to understand as a general principle of selecting de-extinctees. Most people are at least somewhat uncomfortable with the idea of de-extinction. However, when forced to come up with one suitable candidate species, nearly everyone chooses something that is extinct because humans made it so. The Yangtze River dolphin is extinct because we destroyed its habitat. Moa are extinct because we hunted them to death. Dodos are extinct because we introduced cats and rats and pigs to Mauritius, and these cats and rats and pigs made easy meals of all the dodo eggs they could find. If it had not been for humans, each of these species would probably still be alive.
In addition to how a species became extinct, other characteristics of species make them more or less popular choices for de-extinction. Perhaps unsurprisingly, most people would prefer to resurrect herbivores, rather than carnivores. Less obviously, most people choose large species for de-extinction rather than small species, presumably because large species are, well, bigger. And most people choose to bring back animals, as opposed to plants, fungi, or any other living thing.
It is enormously important to make an informed rather than emotional decision about whether an extinct species should be brought back to life. Different species require different technical innovations, different amounts of hands-on manipulation, and different habitat. Some species would be decidedly easier to bring back than others. Some species would proceed relatively straightforwardly through the early steps of de-extinction, such as sequencing their genome, but present potentially insurmountable challenges during later steps, such as identifying suitable habitat into which they could be released. When considering whether a species is a good candidate for de-extinction, it is tempting to focus only on those steps leading to the birth of a newly unextinct animal and to ignore the later steps of rearing and reintroduction into the wild. It is unwise, however, and I would go so far as to say unfair, to proceed through these first steps without careful evaluation of the entire process from fertilized egg to free-living population. What is the point, after all, of bringing a species back from the dead if it is not to reestablish a wild population?
To simplify navigation through the process of selecting a species for de-extinction, I propose seven questions that should be asked and answered. The questions fall into two broad categories. The first set of questions attempts to place that species in the context of its ecosystem. How did it interact with and affect other species when it was alive, and how might that be different today? The second set of questions turns to the nitty-gritty of the science. Is de-extinction of this species—or at least of some specific traits that defined this species—practical, given current and future technologies? My focus for now is on the technical aspects of de-extinction and not the ethical questions that will undoubtedly arise throughout the process. Obviously, these questions are not exhaustive, and not every question applies to every species. These questions do, however, provide a useful means to think through the implications of de-extinction and, perhaps, avoid some potential disasters.
Is There a Compelling Reason to Bring This Species Back?
Probably—and hopefully—the first question that comes to the minds of most people when contemplating de-extinction is “Why?” Why this species? Why now? Why here? As I noted before, most people advocate for species that they know, beyond reasonable doubt, are extinct because of something that humans have done. Bringing these species back goes some way to mitigate the guilty conscience of an ecological savvy human. But mitigation of guilt is not a compelling reason to bring something back to life. I may feel some guilt by association with my Native American ancestors, who probably participated in hunting short-faced bears, and with my European ancestors, who probably were in some way involved with the extinction of the Neandertals. This does not mean I want to bring back short-faced bears and Neandertals. Indeed, in both of these cases, de-extinction for the purposes of alleviating guilt seems remarkably selfish; what kind of existence would either of these have in the world today?1
Compelling reasons to bring something back to life are more likely to relate to the species themselves and the roles these species are likely to play in the environment of the present day. For example, if the species filled a particularly important niche within its ecosystem, then its loss is likely to have resulted in chaotic destabilization of that ecosystem. Bringing it back might restore lost interactions between species and restabilize the ecosystem, in turn saving other species from extinction. The kangaroo rats that I mentioned earlier are a good example of keystone species that play important, stabilizing roles in their ecosystems. The Cascade Mountains wolf—an additional species suggested for de-extinction by a student in my class—is another. Importantly, both of these are very recent extinctions, and their ecosystems may not yet have adapted to accommodate their loss.
The potential role of the Cascade Mountains wolf in maintaining ecological balance can be extrapolated from work in Yellowstone National Park over the last two decades. When wolves were reintroduced to Yellowstone National Park in 1995, many people were convinced this was going to lead to disaster. The cause for concern was that wolves are predators and, as such, would likely depredate livestock from local farms, much to the dismay of the ranchers who depend on the livestock. This was an appropriate concern. As wolf populations have grown, there have been many instances of wolves taking livestock. However, the main source of the wolves’ diet is local wildlife, and elk in particular. By 2006, the elk population in Yellowstone had shrunk to 50 percent of the size it had been when wolves were first reintroduced to the park. Today, elk are no longer overgrazing the plants and young trees that grow along the meadows and valley bottoms, and consequently, woody plants are making a comeback throughout the park. The increase in woody plants provides a greater diversity of habitat for small mammals, whose populations are also on the rebound. Wolves are outcompeting coyotes, whose populations had become much larger after the disappearance of wolves. Fewer coyotes is good news for the animals that coyotes like to eat, including red foxes, pronghorn, and sheep.
Certainly, wolves are predators. Wolves will take livestock when they have the opportunity to do so. However, it seems clear that restoring wolves to Yellowstone National Park has played a critical role in stabilizing the Yellowstone ecosystem.
The Cascade Mountains wolf is a subspecies of gray wolf that lived in the mountains of Washington, Oregon, and British Columbia until around 1940. Based on the positive results of the Yellowstone wolf reintroduction, there is compelling ecological reason to bring back the Cascade Mountains wolf and reintroduce it to its former range.
The case of the Cascade Mountains wolf touches on another intriguing issue. This wolf was a subspecies of gray wolf, and not its own, distinct species. This raises a different question: is it appropriate to select a subspecies for de-extinction?
Before attempting to answer that question, I should first clarify what it means to be a subspecies as opposed to a species or population. From an ecological perspective, a population is a group of individuals of the same species that live together in the same place. The individuals interbreed, compete with each other for resources, and share the same geographic space. A species tends to be defined as an evolutionary lineage that is reproductively isolated from all other evolutionary lineages. Individuals of the same species can move between populations with little consequence to their ability to find a mate and reproduce. Individuals belonging to different species cannot mate. Or if they do, the offspring that are born either do not survive into adulthood or cannot have offspring themselves.
This species-as-reproductively-isolated-lineages concept, known as the biological species concept, was formally described by Ernst Mayr in 1942. The concept turns out to have some flaws. Specifically, some lineages that we strongly believe are separate species are not strictly reproductively isolated. Polar bears and brown bears, for example, are commonly considered to be two different species. But bears born from crosses between brown bears and polar bears survive and can continue to mate and produce offspring. Dogs, wolves, and coyotes can and do interbreed frequently. Cows and bison and yaks can all interbreed and produce fertile offspring. And ancient DNA from Neandertal bones revealed that our species can (and did) mate with Neandertals and that, as a result of this hybridization, Neandertal genes survive in all living humans with Asian or European ancestry.
Why do biologists hold on to this confusing system? As humans, we are compelled to categorize. When we see chaos, we desire to transform that chaos into something ordered so that our brains can make sense of it. Clearly, evolution does not work in absolutes. An animal is not born one day as an entirely new species, incapable of reproducing with anyone in its parents’ species. Instead, speciation is a long process involving many underlying genetic and behavioral changes. Populations become geographically isolated and evolve along independent trajectories. Eventually, enough changes will have evolved so that individuals are incapable of breeding between populations. As we see with brown bears and polar bears and with humans and Neandertals, however, what common sense would call species-level differences will sometimes evolve before the two lineages are completely reproductively isolated.
To impose order on the disorder that is biology, Carl Linnaeus, an eighteenth-century Swedish biologist and physician, devised a taxonomic system to describe and categorize all forms of life. His system provides a hierarchical classification of everything according to its relationship with everything else. The biggest bins classify organisms into kingdoms: Animalia, Plantae, Fungi, Protista, Eubacteria, and Archaeobacteria (although the latter two are sometimes grouped into one kingdom, the Monera). Wolves, dogs, bears, snakes, and rabbits are all animals, so they all belong to the kingdom Animalia. Within that, wolves, coyotes, bears, and rabbits are mammals (class Mammalia). Wolves, coyotes, and bears are carnivores (order Carnivora). Wolves and coyotes are canids (family Canidae). Both belong to the genus Canis, but wolves are Canis lupus and coyotes are Canis latrans, where lupus and latrans are the official Latin names for the two different species.
After that, it gets messy. Sometimes species are subdivided into subspecies. But this is tricky. Some taxonomists will refer to a population that seems to be particularly isolated from other populations as a subspecies, while a different taxonomist might look at the same population and decide that it is not sufficiently different to merit subspecific status. Unlike a species, there’s really no rule to go by to decide whether a subspecies is real or not.
What does all of this have to do with de-extinction? A lot. If a subspecies is not real, or if it is just a slightly different version of something that is not extinct, should time and energy be spent to bring that subspecies back to life?
Sometimes subspecies are defined geographically. This means that, while there may be no physical or genetic barrier to interbreeding, they are simply too far apart in geographic terms for interbreeding to take place. For example, it is not particularly likely that the Iberian wolf will breed with the Mexican wolf. In the absence of interbreeding, the two populations will accumulate genetic differences that make them look and act differently from each other. There is no doubt, however, that both of these are wolves. So, if either Mexican wolves or Iberian wolves were to go extinct, would it be reasonable to use de-extinction technology to bring them back?
Consider a hypothetical scenario in which there are two subspecies that are ecologically very important in their ecosystems—keystone species—and one of these goes extinct, destabilizing the ecosystem in which it lived. The two subspecies are very closely related to each other. In fact, the only things that differentiate them are that they lived in different places and had some small morphological difference—let’s say they had slightly differently shaped ears. To restabilize the ecosystem, we plan to reintroduce the extinct keystone species. Is it better to bring it back to life using de-extinction science or to introduce the close relative? In other words, how different does an extinct lineage have to be from a living lineage to justify its de-extinction?
From a technical perspective, de-extinction of a subspecies like the Cascade Mountains wolf would be much simpler than de-extinction of a distinct species. As I will discuss, assembling the genome sequence of an extinct organism can be extremely challenging and requires a guide genome to act as a scaffold onto which the short, damaged fragments of ancient DNA can be mapped. The Cascade Mountains wolf genome could be assembled using another gray wolf genome as a guide, simplifying this process greatly. Cascade Mountains wolf embryos could be implanted into a mother gray wolf, and families of gray wolves could rear the pups in established gray wolf packs. This then raises the question, how would the purportedly unextinct Cascade Mountains wolf differ from the wolf subspecies into which it is born? Would it be preferable simply to introduce another gray wolf subspecies into the Cascade Mountains?
While some species or subspecies seem too similar to living species to justify their de-extinction, other extinct species have no evolutionarily close living relatives. This argues both for their de-extinction, because bringing them back will restore more evolutionary novelty than would bringing back something that has a close, living relative, and against their de-extinction, because bringing them back will be much more costly to achieve.
Moa, for example, were subdivided into three extinct families within the order Dinornithiformes, which has no living representative. The closest living relative to the moa is the tinamou, and the common ancestor of moa and tinamou lived around 50 million years ago. The moa represents a long history of independent evolution, and bringing it back would restore many unique traits to the world. However, with no close relative, it would be extremely hard to assemble the broken bits of DNA recovered from moa bones into a reasonably accurate moa genome. In this case, the guide genome would be more than 100 million years diverged—twice the evolutionary distance to the common ancestor—from the ancient genome. The same is true for any extinct species that lacks an evolutionarily close living relative. Identifying appropriate surrogate mothers or eggs within which the embryos could develop would be extremely challenging for species that lack close living relatives. We would also have little way to know what the native behaviors of such species should be, how much parental care would be required to rear them, or how to mimic this parental care or other important social interactions. In a sense, these individuals might be too different from anything that is alive for de-extinction to succeed.
The ideal candidate for de-extinction has both sufficiently closely related living relatives to make its de-extinction feasible and unique traits or adaptations to a particular habitat. The golden toad, for example, was last seen in the cloud forests of Costa Rica in 1989. It was such a peculiar, bright orange color that Jay Savage, the herpetologist who described it, had trouble believing it was real and not some trick. The golden toad was tiny—adult males measured a bit more than five centimeters in length—and is a good candidate for de-extinction. It belongs to the genus Bufo, which is species-rich and diverse and therefore has many close relatives that are not extinct. However, among its many close relatives, the golden toad was the only Bufo to display such a striking orange color. What if the proteins that made the orange color had some undiscovered medical purpose, or psychoactive properties? We’ll never know until somebody licks one, and for that we’ll have to bring it back.
Finally, the ideal candidate for de-extinction may be one that has only recently gone extinct. Ecosystems are constantly in flux. They are influenced by abiotic changes, such as how much rain falls or how cold the winters are, and biotic changes, including both species extinctions and introductions. When a species goes extinct, the ecosystem in which it lived changes to adapt to its disappearance. If the extinction took place many thousands or possibly even only several hundred years ago, it may be that reintroducing that species would actually destabilize whatever new equilibrium that ecosystem had achieved. This does not mean that the only acceptable de-extinctions will be of recently extinct species. Certainly, some species, such as large herbivores, played roles in ancient ecosystems that have not been filled in their absence. As I discuss below, it may be that careful evaluation of the relative risks and rewards of their de-extinction will lead to the conclusion that they, too, should be brought back on the grounds of their potential beneficial impact on the environment of the present day.
Why Did This Species Go Extinct the First Time?
People tend to be most interested in bringing back species that went extinct because of human activities. Given that proposition, asking the question “Why did they go extinct?” seems a bit silly. In fact, it is not at all silly. Humans are remarkably creative when it comes to killing things.
We killed many species using brute force. In the nineteenth century, we netted and shot billions of passenger pigeons, eventually leading to their extinction (figure 3). When Europeans arrived in North America, passenger pigeons were thought to make up 25–40 percent of the bird population in the eastern United States. In 1866, a report described a single flock of more than 3.5 billion passenger pigeons flying across the Ohio skies, taking more than fourteen hours to pass overhead. At one o’clock in the afternoon on the first of September 1914, the last remaining passenger pigeon, named Martha, died in captivity in the Cincinnati Zoo (plate 1).
Figure 3. A flock of migrating passenger pigeons. Drawing from The Illustrated Shooting and Dramatic News, July 3, 1875. ©The Archives and Manuscripts Department, John B. Cade Library, Southern University and A&M College.
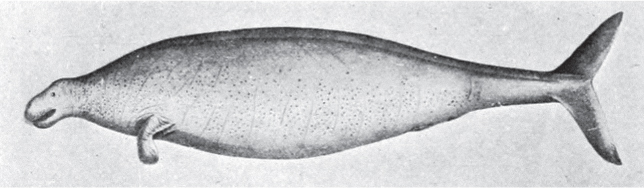
Extinction due to overexploitation is a common theme in human history, and a human tendency that we are still grappling to overcome. Steller’s sea cow was a large—nine meters long and up to ten tons in weight—marine mammal whose closest living relative is the dugong (figure 4). Steller’s sea cows were once abundant throughout the North Pacific but were hunted to death after their discovery in the eighteenth century. Overexploitation is also blamed for the extinction of the great auk, which was taken for its fat, feathers, meat, and oil. We continue to overexploit the species we rely on today. In 2012, the State of the World Fisheries and Aquaculture Report reported that 30 percent of the world’s fisheries were overexploited and would require strict management to be sustainable into the future.
Figure 4. Steller’s sea cow, Hydrodamalis gigas. Illustration by J. F. Brandt, 1846. This image was published in Extinct Animals, by E. R. Lankester (London: A. Constable, 1905).
But we don’t only kill things with brute force. Indirect effects of human population growth—including the conversion of wild habitat for cities, towns, and agricultural land; deforestation; monoculture; and the construction of roads and highways to connect all of these things—result in changes to habitats that disrupt and destabilize ecosystems, leading to extinctions. Bird species are particularly susceptible to habitat destruction; the gradual clearing of forests in order to make way for people and agriculture on the islands of the Pacific alone has been blamed for the extinction of thousands of bird species. Indeed, habitat destruction accounts for more than half of the endangered birds of the world today.
We also move around the world, and as we move we bring stuff with us, both accidentally and intentionally. We introduce parasites, predators, and competitors to ecosystems in which these organisms did not exist previously, resulting in extinctions. Again, birds on islands are particularly susceptible to introduced predators, especially rats, cats, and snakes, which are keenly adept at finding and consuming eggs. Rats and cats are blamed for the extinction of the Tahitian sandpiper and Society parakeet in the Society Islands; the dodo, solitaire, Reunion pigeon, Rodrigues starling, and red rail in the Mascarenes; and the Raiatea parakeet, white-winged sandpiper, and Maupiti monarch in French Polynesia, to name only a few. In addition to introduced predators, diseases brought to these new locations spread from domestic to wild species, also causing extinctions.
And, with the byproducts of agriculture and industry, we pollute the world around us. The Yangtze River dolphin is an example of how pollution, combined with habitat destruction, can lead to extinction. The Madeiran large white butterfly was declared extinct recently, its extinction blamed on a combination of habitat destruction and pollution from agricultural fertilizers.
In some cases, even when it is clear that humans are ultimately at fault for an extinction, it is very difficult to know what the proximate causes of that extinction were. If the cause of the extinction is not completely understood, the species is most likely not a good candidate for de-extinction. If we don’t know what caused a species to go extinct the first time, how can we know that it won’t quickly become extinct again? Equally, some species for which we do know the most proximate cause of extinction are poor candidates for de-extinction. For example, despite the emotional allure of dodo de-extinction, if we were to bring dodos back and reintroduce them to Mauritius, their eggs would be promptly gobbled up by the large rat and cat populations that thrive on the island today.
Is There a Place for This Species to Live if We Successfully Bring It Back?
If we know why the species we are considering for de-extinction went extinct in the first place, is it possible to correct whatever went wrong? In the case of the dodo, we would have to create a cat- and rat-free zone on Mauritius into which the unextinct dodos could be placed. If this is not possible, either because there is no habitat that could be set aside for these purposes or because it’s just too hard to keep out rats and cats, then the dodo is not an appropriate candidate for de-extinction.
The loss of suitable habitat—whether by deforestation, development, pollution, or the introduction of parasites and predators—seems to be the most common cause of extinctions that have been caused indirectly by humans (if we assume that humans only cause extinctions directly by overexploitation). Yet, more and more humans live on this planet every day, which means we take up more and more space and need more and more food, which translates into the increasingly rapid and destructive conversion of natural habitat into land for human use. A major challenge to any de-extinction effort will therefore be to secure suitable habitat for the unextinct species. Suitable habitat will have to (i) include appropriate prey or food to sustain the unextinct population; (ii) exclude predators or competitors (including invasive species) that would drive the species back into extinction, while at the same time leaving sufficient numbers of carnivores in the system so as not to destabilize the food web; (iii) be devoid of disease, parasites, or pollutants; (iv) imitate as much as possible the temperature and precipitation regimes of the native habitat of the species; and (v) be sufficiently large to support a self-sustaining population.
Interestingly, this whole process might be simpler for species that we drove to extinction directly through hunting, as their habitat may still exist. Of course, the survival of these species would rely on our not hunting them to death a second time. Like exotic species that are alive today, unextinct species would have to be protected from increasingly creative and dangerous poachers, using laws and statutes that are in many cases difficult to enforce.
How Will Introducing This Species Affect the Existing Ecosystem?
The extinction of a species changes the ecosystem in which it once lived. Over time, the ecosystem restabilizes, and the niche that was once filled by the extinct species is either filled by a different species or eliminated. The longer a species has been gone, the more likely it is that the ecosystem has adapted to its absence. So when the species is reintroduced, what effect will that reintroduction have on the species that live there today?
When passenger pigeons flocked in large numbers across eastern North America, the landscape was different than it is today. The deciduous forest was more widespread, American chestnut trees were plentiful, and people were not. Passenger pigeons were a dominant and destructive force in the eastern deciduous forest. They fed primarily on large seeds: oak acorns and the nuts of hickories, beeches, and chestnuts. Flocking in the billions, hungry passenger pigeons could destroy the entire seed crop of a forest stand in very little time. When they nested, as many as five hundred birds would nest in a single tree, and when they left the nests, they tended to leave behind dead trees covered in bird droppings. When they went extinct, this avian version of a non-stop EF5 tornado came to a screeching (squawking?) halt. Since that time, humans have converted much of the historic deciduous forest into towns, cities, and agricultural land. What would a flock of a billion passenger pigeons eat today? What effect would their de-extinction have on the remaining deciduous forest? On agriculture? On the bird species and other animal species that live there today, with which unextinct passenger pigeons would compete for access to food and nesting grounds?
It may be that some unextinct species would have minimal destabilizing effects on present-day ecosystems. Careful evaluation of the consequences of reintroduction is required, however, and not simply from the perspective of whether that particular species would survive after reintroduction. If a species’ de-extinction would lead to changes to the existing habitat that in turn would threaten living species, then that species is not a good candidate for de-extinction.
Finally, as disingenuous as it may seem at first, it would be remiss not to think about the effect that the de-extinction would have on human populations. Few East Coasters would be overjoyed at the site of a billion passenger pigeons darkening the sky just above their freshly manicured lawns and newly waxed SUVs, for example. But there are subtler aggravations that would probably make this particular de-extinction unpopular. If unextinct passenger pigeons were protected as endangered species, people who enjoy hunting pigeons for sport may find themselves facing new restrictions about when and where pigeons can be hunted, or even whether pigeons can be hunted at all, given the presumed difficulty of distinguishing the unextinct passenger pigeons from common pigeons that were once unprotected game. And a billion passenger pigeons would probably require quite a lot of protected habitat, which would have to be repurposed from somewhere.
These problems of course extend beyond passenger pigeons. If new and different categories of protected species emerge, regulations enacted to prevent their (re)extinction may make previously accessible wilderness off-limits to recreation, much to the chagrin of hunters, campers, hikers, and so on. Farmers are unlikely to support de-extinction of species like the Carolina parakeet, which was driven to extinction precisely because it was an agricultural pest. And ranchers already unhappy about the idea of wolf reintroduction will hardly embrace the idea of saber-toothed cats roaming freely in proximity to their livestock, looking for an evening meal.
Other species are less objectionable on the grounds of how much they might annoy their human neighbors. The mammoth, for example, may be one of the least potentially annoying species on the list of candidate species for de-extinction. The most appropriate habitat for the mammoth is, after all, the Arctic, where human populations are sufficiently small and isolated that the mammoth is not likely to get in their way.
Indeed, Dr. Sergey Zimov, the director of the Northeast Science Station of the Russian Academy of Science in Cherskii, Russia, is intent on recreating the habitat of the mammoth so that it has somewhere to live once its de-extinction is successful. His Pleistocene Park is a nature reserve on the Kolyma River, south of the science station in Cherskii, in northeastern Siberia. Pleistocene Park is in the most northerly part of the habitat that was once the mammoth steppe—the rich tundra grassland that supported mammoths and other grazing herbivores during the Pleistocene ice ages (plate 2). Zimov has already introduced horses from the Urals, bison from eastern Europe, and four different species of deer into Pleistocene Park, and these populations are healthy and self-sustaining. Recently, Zimov decided to expand his operation and establish a second Pleistocene Park in a more southerly location, where the less harsh climate is more amenable to supporting large numbers of herbivores. This second park, which he calls Southern Pleistocene Park, is located in the Tula region, approximately 250 kilometers south of Moscow. Over time, Zimov plans to introduce bison, auroch, horses, wolves, and large cats into Southern Pleistocene Park, which—unlike the Pleistocene Park in northeastern Siberia—is easily accessible by car from Moscow. Both of these locations would likely provide appropriate mammoth habitat, re-creating communities that existed more than ten thousand years ago that should be able to persist without bothering or being bothered by humans.
Will It Be Possible to Learn the Genome Sequence?
With this question, we transition from the big picture to the finer grains of de-extinction. In other words, we now ask whether de-extinction is technically possible, or whether it will become so in the foreseeable future. Each of these topics will be covered in detail in the next chapters and so are touched on only briefly below.
The first technical step in de-extinction is to learn the genome sequence of the extinct species. Well, not just the genome sequence. Really, we want to know what the key genetic differences are between the extinct species and any closely related living species. I’ll explain what that means in more detail later, but for now let’s simply ask: “Can we sequence all of the nucleotides in the genome of this extinct species and then piece those nucleotides together to learn what the sequence of that genome is?”
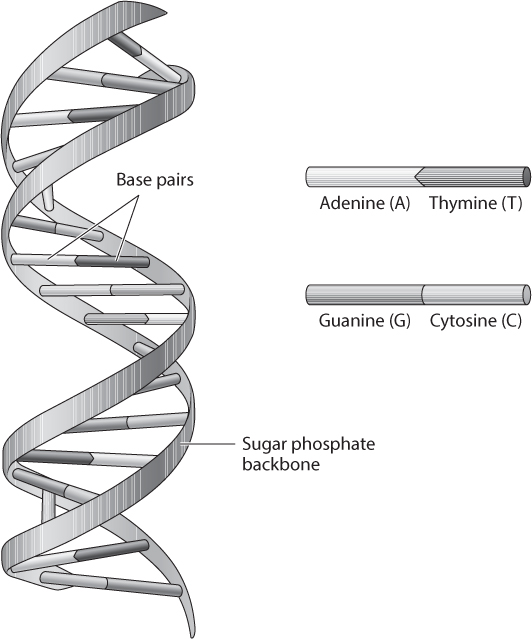
First, some vocabulary. Genomes are big places, but the molecules that make up genomes are tiny (figure 5). Genomes are made up of chromosomes, which in turn are made up of long strands of nucleotides—the building blocks of DNA. Nucleotides each contain a nitrogen base, a five-carbon sugar, and a phosphate group. DNA genomes contain four different nucleotides, each with a different nitrogen base: adenine (A), guanine (G), cytosine (C), and thymine (T). Nucleotides are strung together along a sugar phosphate backbone to make up nucleic acids like deoxyribonucleic acid, or DNA. Genomic DNA is double-stranded, which means that when it is in a stable state, the nucleotide on one strand is bound to a complementary nucleotide on the other strand. A nucleotide that is paired to its complementary nucleotide is called a base-pair. Genome sizes are usually reported in base-pairs, which will be half the number of nucleotides.
Figure 5. The structure of DNA. DNA is made up of four chemical building blocks called nucleotide bases: adenine (A), cytosine (C), guanine (G), and thymine (T). DNA exists in a winding, two-stranded “double helix” structure, which is formed because the nucleotide bases pair up with each other, creating a ladder-like structure and connecting the two strands. The order of the nucleotide bases, which is also known as the DNA sequence, contains the information necessary to build and maintain an organism.
Genomes vary considerably in the number of base-pairs they contain and in the number of chromosomes among which these base-pairs are distributed. The human genome comprises around 3.2 billion base-pairs, which are found on 23 chromosomes. The loblolly pine genome comprises 22.2 billion base-pairs but only 12 chromosomes. The common carp genome contains 1.7 billion base-pairs distributed among 100 chromosomes. The huge variation in plant and animal genomes is linked neither to the complexity of the organism nor to the number of genes that are encoded by those genomes.
Chromosomes are too long to sequence all at once using existing sequencing technologies. When scientists sequence DNA, they therefore begin by shearing the chromosomes into smaller fragments. These fragments are double-stranded, so their length is also reported as a number of base-pairs. Depending on the sequencing technology to be used, these fragments will vary from fewer than 100 base-pairs long to several thousand base-pairs long. After the DNA has been sheared and sequenced, the fragments are reassembled into chromosomes. To summarize the process of sequencing a genome: First, cut it up. Then, put it back together again.
Now that some of the jargon has been demystified, let’s outline the steps of sequencing and assembling the genome of an extinct species. First, we collect remains from the species that we plan to bring back to life—bones, teeth, skin, hair, whatever we can find. Then, we extract and collect as much DNA as we can from those remains. Next, we sequence the DNA. Finally, we take that DNA and carefully assemble the tiny pieces together to make bigger and bigger pieces, and eventually chromosomes.
If you were paying attention, you’ll have noted that we skipped the step in which we shear the DNA into smaller fragments. When working with ancient DNA, we don’t need this step. The DNA comes pre-sheared. In fact, over-sheared is a better way to refer to it. Over-shearing is bad: the shorter a fragment of DNA is, the harder it is to figure out where it goes in the genome.
There is more. These short DNA fragments are also in pretty bad shape. Thanks to chemicals and other biomolecules in the environment, individual nucleotides can become broken or damaged in a way that changes their molecular structure. Molecules with altered structures are read incorrectly during the sequencing process, resulting in mistakes in the genome sequence. The rate of DNA decay is slower in some environments (for example, in the Arctic, where the mammoth lived) than in others (for example, in the tropics, where the dodo lived). This means that species whose native ranges did not include regions of the world where remains are likely to be preserved are probably not ideal for de-extinction.
Finally, we have to deal with what we call contamination. Contamination in the broadest sense refers to any DNA that is co-extracted from the bone or other tissue that does not come from the organism whose genome we are trying to sequence. It might be DNA from microorganisms that colonized the bone after it was buried in the ground, or from plants whose roots grew around the bone while it was in the ground. It might also be DNA that was introduced into the bone as it was being excavated or handled in the lab. A bone might contain an enormous amount of preserved DNA only a very tiny fraction of which is of interest to us.
Professor Svante Pääbo leads a research group at the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany, and he and his research group have recently sequenced and assembled the Neandertal genome. His group is very interested in understanding what it means to be human. One way to approach this question is to compare the human genome with closely related genomes of great apes and ask what genetic changes have happened within our genome sequence since we diverged from our common ancestor with other great apes. Our closest living relative is the chimpanzee. The human and chimpanzee genomes are around 98–99 percent identical, which means that what distinguishes us from chimpanzees probably has to do with the other 2 percent. But 2 percent of a 3.2 billion base-pair genome is still a lot of DNA to sort through. Neandertals are much more closely related to humans than are chimpanzees. By sequencing the Neandertal genome, Pääbo is able to focus more narrowly on those genetic changes that are unique to our species.
The first complete Neandertal genome that Pääbo’s team published was a combination of DNA data that was sequenced from three different Neandertal bones. Each bone contained less than 5 percent Neandertal DNA, with the remaining 95 percent or more comprising mostly environmental DNA—soil microbes and their pathogens, plants, and the like. Of the Neandertal DNA sequences that were recovered from these bones, the average fragment length was forty-seven base-pairs. The human genome contains 3.2 billion base-pairs, so this is a bit like having a puzzle that can only be solved by correctly assembling 68 million puzzle pieces. Of course, thanks to damage and contamination, what they actually had was far more pieces than they needed, some of which were from the same puzzle but cut in a different way and some of which actually belonged to a different puzzle.
To help them to assemble the Neandertal genome, Pääbo’s team used the human genome, which was already sequenced and assembled, as a guide. To extend the puzzle analogy, if the forty-seven base-pair fragments of Neandertal DNA were the puzzle pieces, the human genome was the picture on the box top. Only that picture (because it was of a human and not a Neandertal) was not exactly the same as what the puzzle would look like when it was finished. Not identical, but close—maybe the picture was a different color, or maybe part of it was covered by a text box stating “contains very small parts.”
Assembling the Neandertal genome was not an easy task. However, it was much easier than assembling many other paleogenomes will be. First, the human genome is the best-resolved genome of any species to date, so the picture on the puzzle box top was nearly complete. The number and diversity of sequenced genomes is growing, but most species genomes are still only partially sequenced and assembled. Second, humans and Neandertals have a common ancestor within the last million years, probably closer to half a million years ago. This means that there hasn’t been much time for a huge number of differences to evolve between humans and Neandertals. The picture on the box top pretty closely reflected what the final puzzle would look like.
Not so for many species. In fact, the more evolutionary time that has passed between the divergence between the extinct species and the living species that would be used as a reference, the harder assembling the genome will be. At some point, the picture on the box goes from a slightly discolored version of the end product to something that you rescued from your dog’s mouth and then tried to piece together using your imagination and some sticky tape, to something that a herd of mammoths trampled while escaping a pride of cave lions. In the rain.
If there are no remains that contain recoverable DNA, then the species is not a candidate for de-extinction. If there are remains with recoverable DNA, but the species has no close relatives, assembling the genome from that DNA will be challenging—maybe very challenging. Critically, however, it is not impossible to assemble at least large chunks of the genome, even when the DNA that is preserved is in terrible condition.
Is There a Way to Transform the Genome Sequence into a Living Organism?
If we’ve made it to the step in which we’re considering how to create a living organism, we presumably have been able to generate a genome sequence (or a partial genome sequence), even if it might have been tough to do so. Now we have to transform that string of letters into a living thing. How?
There is no clear path from genome to living thing that can be followed for every organism. Some genomes, such as those sequenced from bacterial or viral organisms, are likely to require very little push to start behaving like they are alive. Other genomes are nowhere near becoming a living thing.
Two paths are generally considered when contemplating de-extinction. The first is to do what most people are referring to when they talk about cloning. To clone Dolly the sheep in 1996, scientists at the Roslin Institute, which is part of the University of Edinburgh in Scotland, removed a small piece of mammary tissue that contained living cells from an adult ewe and used the DNA in these cells to create an identical copy of the adult ewe. This process is called somatic cell nuclear transfer, or, more simply, nuclear transfer. I’ll explain how it works later, but for now it is sufficient to know that this is not likely to be the process used to bring many extinct species back to life. Cloning by nuclear transfer, unfortunately, requires intact cells. Unless tissue was taken from a living individual prior to the species’ extinction, nuclear transfer will not work. If we’re dealing with a species whose genome we have had to sequence and assemble, then we need a different approach.
The other path to creating a living organism is eerily reminiscent of the movie Jurassic Park. As is likely to be true in real-life de-extinction projects, Jurassic Park scientists were only able to recover parts of the dinosaur genome—in the case of the movie, from the mosquito blood that was preserved in amber. When they came across holes in the dinosaur genome, they used frog DNA to complete the sequence. Unfortunately, they weren’t able to know beforehand which bits of DNA were important to making a dinosaur look and act like a dinosaur, and which bits were just junk. We can only assume that these fictional scientists were hoping that the holes that they were filling in were mostly in the junk-containing regions of the dinosaur genome. But, of course, they were wrong, and some of that frog DNA let the unextinct dinos switch sexes miraculously, leading to disaster and $400 million in box office earnings.
In real-life de-extinction science, the plan is to know which parts of the genome are important in making the extinct species look and act the way it did. We would then find the corresponding parts of the genome of a close living relative, cut out these important sequences, and replace them with the extinct species’ version.
Of course, this is all easier said than done.
Let’s say we want to bring a mammoth back to life by editing an elephant genome to look like a mammoth genome. First, we have to learn what all the differences between the elephant genome and the mammoth genome are. Then, because making all the changes might be too much to accomplish (at least in the first de-extinctions), we could narrow down which changes to make by deciding which of the differences are important. We might learn, for example, that mammoths have a different copy of a gene called Ucp1—mitochondrial brown fat uncoupling protein 1—than elephants do. Experiments with mice have shown that Ucp1 is involved with thermoregulation. Since mammoths lived in very cold places and elephants do not, we might hypothesize that the mammoth version of this gene helped mammoths to stay warm. Our goal is to turn an elephant into an animal that can survive in cold places, and converting this gene from the elephant version to the mammoth version would help to achieve that goal. So, we construct a molecular tool that can go into an elephant cell, find the spot in the genome that codes for the Ucp1 gene, and edit that gene so that it looks like the mammoth version.
To make the complete mammoth genome, all we have to do is repeat this for every important difference between mammoths and elephants.
Next, we take the cell with the edited genome and inject it into an egg cell that has had its nucleus removed. That cell begins to divide and develop into an embryo, following the familiar path of cloning by nuclear transfer. We then place that embryo into the uterus of a surrogate mom, where it continues to develop and is eventually born.
That last step, in which one species is developing within the uterus of another species, might sound pretty straightforward. However, it also requires some careful consideration. Imagine a project to resurrect Steller’s sea cows. Dugongs, the closest living relative of Steller’s sea cows and therefore the most likely surrogate mom, have a thirteen- to fifteen-month gestation period, after which they give birth to a single calf. Newborn dugongs weigh about thirty kilograms and are a bit over a meter long—about one-third to one-half the length of an adult dugong. If the same size ratio applies to Steller’s sea cows, a newborn calf will be somewhere in the range of three to six meters long. Longer, at birth, than his surrogate mom.
To get around this, one might design a giant sea cow artificial womb. Or, perhaps, a better solution is to choose a species for de-extinction with more suitable options for surrogacy.
Will It Be Possible to Move the Resulting Living Organism from Captivity to a Natural Habitat?
Although much of this has been covered in the answers to the first four questions, I’d like to raise a few additional points here that should be considered when selecting a species for de-extinction. I discussed above whether appropriate habitat exists and what might happen to that habitat and ecosystem if an un-extinct species were suddenly reintroduced. Here, I’m thinking of the more technical aspects of reintroduction. How behaviorally hard-wired was the species? How much parental care was involved with rearing offspring? Were their behaviors learned, or were they born already knowing how to survive, find food, and find a mate? How social were they? Although I brought this up briefly when discussing the additional complexities of bringing back a species with no close living relatives, these are challenges that, to some extent, will face any de-extinction. The first unextinct individual of any species will necessarily be all alone in this world. If behaviors have to be learned, from whom will they be learned? Interaction with the surrogate mom or surrogate community might replace some of the missing social interactions. However, if behaviors are learned from these interactions, will they be the same behaviors that the extinct species would have exhibited? And is that important?
We know from ongoing conservation work that some species survive and seem to do well in captivity but fail to thrive once they are released into the wild. The reason for failure to survive in the wild differs among species. Sometimes animals bred and raised in captivity are easier prey when released, never having been trained to sense and flee from predators. Sometimes they lack the social structure in the wild that they need to be successful. And sometimes they simply never learn how to find their own food. In all of these situations, the only way the species would survive in the wild is if the wild was not actually the wild but instead a site that is actively managed by people. The economic cost of such hands-on management, which may not be small and is likely to take resources away from other conservation and wildlife management programs, needs to be weighed against what is gained by the de-extinction.
THE ELEPHANT IN THE ROOM IS A MAMMOTH
At the beginning of this chapter, I raised the question of who gets to decide what the first targets of de-extinction should be. When I asked the students in my de-extinction class which candidate they thought should have that privilege, they responded with complete silence. Eventually, they offered the only solution appropriate for a group of Californian students: it should be a group decision. But what group? And, even groups have to have a leader, someone who decides ultimately how the group will respond.
The truth is that, at least in these early stages of de-extinction research, the decisions about which species to bring back are going to be made by the people with the interest, money, and expertise to make it happen. The European team working to bring the bucardo back to life is just as unlikely to refocus its attention on the Tasmanian tiger as the Australian team working on the Lazarus frog is to lead the de-extinction of the Yangtze River dolphin. Unfortunately, money is probably the most important determining factor in whether a de-extinction project gains traction. After the death of the baby bucardo in 2003, the bucardo project went silent due to lack of funds. In 2013, after the fresh bout of attention to the project that followed the TEDx event in Washington, DC, the Hunting Federation of Aragón allocated new funds, and the team restarted their cloning efforts. Money, rather than any of the ideas discussed above, may also decide which species are selected for de-extinction. In their campaign to raise funds for de-extinction efforts more broadly, Ryan Phelan and Stewart Brand of Revive & Restore are targeting potential donors on Martha’s Vineyard, a wealthy enclave in Massachusetts just south of Cape Cod, asking residents to consider whether they’d like to see Heath hens roaming the island just as they did during the early twentieth century.
And then there is the mammoth. There may be compelling ecological reasons to bring the mammoth back to life, and I will discuss these later. It is also true that mammoths may face fewer technical hurdles in their de-extinction than other species might face. They lived in cold places and many well-preserved bones can be collected and used for DNA analysis. Their closest living relative is the Asian elephant, from which it diverged around 5–8 million years ago, and elephant moms are probably reasonable surrogates for baby mammoths. There is even a place for resurrected mammoths to go: Pleistocene Park is likely to provide a suitable place for mammoths to live, although the steppe tundra that dominated the landscape during the mammoth’s reign is not found anywhere on Earth today. That is not to say that mammoth de-extinction would be without challenges. Elephants reach sexual maturity between ten and eighteen years old and have a nearly two-year gestation, which means that genetic engineering experiments will take a long, long time. Also, elephants are extremely social creatures, and there is no reason to suspect mammoths were not highly social as well. Recreating social contexts into which mammoths can be placed will be key to their survival and an additional challenge to overcome.
What inspired the mammoth de-extinction project was not that it would be easy or hard to accomplish, or that it might be ecologically beneficial to have mammoths roaming around Pleistocene Park (although, as I discuss later, the latter is almost certainly true and has become a motivating force as this research continues). The reason that George Church and his group at Harvard’s Wyss Institute selected mammoths, rather than kangaroo rats, as the focal species for developing the genetic engineering technology necessary for de-extinction is that mammoths are mammoths whereas kangaroo rats are, well, rats.
Stewart Brand says that his motivation to bring back the passenger pigeon is that these birds, in cultural terms, are as iconic as bald eagles. He believes that the highest value of resurrected passenger pigeons would be to inspire people to be more aware of and engaged in conservation. He puts it more poetically: “Flocks in memory, and flocks in prospect, can make the heart sing.” Of course, passenger pigeons are iconic because they formed ludicrously large flocks, which may be difficult to recreate, sustain, or tolerate.
In addition to the challenges of creating and sustaining enormous flocks, passenger pigeon de-extinction faces more (or at least different) technical challenges than does mammoth de-extinction. A high hurdle for passenger pigeon de-extinction is that it is not currently possible to transfer engineered nuclei into bird eggs. There is also no assembled genome sequence yet available either for the passenger pigeon or for its closest living relative, the band-tailed pigeon, but we’re working on that (plate 3). We also don’t know the extent to which the passenger pigeon was a social creature. The enormity of their flocks suggests they may have been highly social, but whether they need these large flocks to survive remains unknown. The Bronx Zoo, part of the Wildlife Conservation Society, is creating a habitat for the captive breeding of passenger pigeons, but as for whether they will ever be released into the wild, that’s also a big unknown. One benefit of choosing passenger pigeons is their generation time. They reproduce every year, so the research necessary to bring them back to life could proceed at a relatively quick rate.
Both the mammoth de-extinction project and the passenger pigeon de-extinction project will require new technologies in order to succeed. But how close are we to seeing either of these species brought back to life? What are the next steps, now that these two species have been selected for de-extinction? Knowing what to do first is easy. First, we have to find the right specimens and extract their DNA.