FIND A WELL-PRESERVED SPECIMEN
One winter morning a few years ago, I met Mathias Stiller and Tara Fulton—two postdoctoral research fellows working in my lab—in a dark, sub-basement hallway of the physics building on our university campus. The dark hallway was home to our ancient DNA lab, which was a purpose-built facility for extracting DNA from poorly preserved samples. Artificial lights flickered ominously overhead as we shed our coats, bags, and shoes, leaving them in the row of outside lockers. Anything that might carry hitchhiking fragments of DNA from the outside world was strictly forbidden to enter the lab. We unlocked the door and moved into the anteroom. The air reeked of the bleach we routinely use to sterilize the floors, surfaces and walls. We dressed in the traditional getup of an ancient DNA scientist: full-body suit, sterile boots, two layers of sterile gloves, hairnet, facemask, and goggles. When we were ready, meaning that no skin or hair or piece of nonsterile clothing was exposed, we moved from the anteroom into the main part of the lab. Tara carried a smoking container of dry ice. Mathias carried an enormous mallet (sterilized, of course). And I carried a tiny plastic bag.
In the plastic bag was a stunning seventeen-million-year-old piece of amber. We had acquired this treasure from my colleague, Blair Hedges, who purchased it for precisely the purpose we had in mind. The amber weighed around eight grams, measured five centimeters long and three centimeters tall, and was a centimeter or two thick at the center. Encased within the amber were hundreds of tiny bees that had become trapped in the sticky tree resin millions of years ago and, to our eyes at least, were perfectly preserved.
We proceeded to the back corner of the lab, where we had installed a thick, sterile stone plate, above which hung a bright white light and movable magnifying glass. We removed the amber from the bag and wiped it down with a bleach solution so that DNA from anyone that may have touched it over the years would be destroyed. We then rinsed it twice with ethanol to wash off the bleach and allowed it to dry for a few minutes. As it dried, we waited in silence.
When we were certain that enough time had passed, Mathias picked up the amber with sterile forceps and placed it gently into the container of dry ice. Then, we waited again.
Although amber is fossilized tree resin, it is still somewhat malleable—anyone who has ever touched amber jewelry will know what I mean. Stabbing amber with something sharp might dent it, but amber is nearly impossible to break or chip. We wanted to get this piece of amber really, really cold, so that it would be hard and rigid. Brittle.
After ten very long minutes, Mathias picked the amber out of the dry ice with the forceps and placed it carefully on the stone. I then raised the mallet and smashed the little glimmering piece of geological history over and over again until it shattered into a zillion tiny, shining, sticky chunks. Then, using the magnifying glass, we sorted the amber from the bees (plate 4). This involved a lot of re-freezing, re-smacking, and skilled operation of tweezers. After a few hours, we had one tube of mostly amber and another of mostly bees. We took the tube of bees and stuck it in the freezer. We were done for the day.
The next morning, Mathias began the process of extracting ancient DNA from the amber-preserved bees. Over the years, people working in the field of ancient DNA have developed highly sensitive DNA extraction protocols for situations like this one. If DNA had survived in these bees, there certainly wouldn’t be much of it left. Mathias opted for the extraction protocol that had proved most successful in recovering very old DNA. We were giving it our best shot.
When the extraction experiments were complete, it was time to send the results off to be sequenced. And then wait. The sequencing results came back three weeks later. We got nothing.
I was disappointed. How incredible would it be to recover DNA from insects preserved in amber? And by incredible, I mean implausible. Far-fetched. Unbelievable. Mathias, I think, was relieved. We both knew that if we did get a result that suggested that millions-of-years-old DNA was preserved, that result would have taken over our lives. We would have had to spend considerable time and energy first convincing ourselves that it was real, and later convincing our colleagues that we had not made a mistake.
When staring into a piece of amber that contains a preserved biological organism, it is hard to understand why the community of ancient DNA scholars would be so skeptical of DNA recovered from that organism. Insects, frogs, and even a 23-million-year-old lizard have been found in fossilized amber, all in perfect physical condition. Why should their DNA not be preserved to the same extent?
The unfortunate truth is that DNA simply does not survive for millions of years. If we did recover authentic ancient DNA sequences from amber, we would have broken all the rules that we’ve come to understand about DNA preservation and decay.
WHAT? NO JURASSIC PARK?
Millions of years before amber is amber, amber is a substance called copal. Thousands of years before it is copal, it is tree resin. Tree resin is a sticky, amorphous, organic substance that is secreted mainly by conifers—pines, cypresses, cedars, sequoias, for example. The resin serves a variety of purposes. It protects the tree from injuries and infections. It may help to heal major wounds, like broken branches. It is also pretty smelly, which might attract curious insects. As the resin oozes out of the tree, bits of plants, insects, and other small animals get trapped and sometimes covered entirely by the sticky substance. Over millions of years, the volatile organic compounds in the resin evaporate, leaving behind only the nonvolatile compounds that make up the amber and anything encased within the amber.
Key to the remarkable preservation of amber-preserved animals is probably the speed in which they are engulfed by resin. If the animal is completely encapsulated and killed almost instantly, that leaves little time for bacteria from the gut or the environment to colonize and start the decomposition process. The tissues also become rapidly dehydrated, killing the enzymes within that would otherwise break down their DNA.
This idea—that amber might provide an exceptional environment for ultra-long-term DNA preservation—is precisely the rationale used by scientists in the early 1990s when they tried this experiment for the first time. Unlike us, however, these scientists claimed success. They shouted it, in fact, in reports that appeared in the most respected scientific journals.
In the early 1990s, the field of ancient DNA was just gaining traction as a serious scientific endeavor. DNA sequences had been recovered from a 170-year-old quagga (an extinct relative of the zebra), from human mummies that were thousands of years old, and from Neandertals and mammoths that were more than thirty thousand years old. Researchers were only beginning to appreciate what this ancient DNA could reveal.
The first applications of ancient DNA were taxonomic: to identify the living species that are the closest evolutionary relatives of extinct species. We now know, for example, that Asian elephants are more closely related to mammoths than are African elephants and that the closest living relative of the dodo is the ornate and beautiful Nicobar pigeon. Some of the taxonomic results from the analysis of ancient DNA have been surprising. In New Zealand, three different species of giant moa (Dinornis) had been described based on differences in the size of their bones. Ancient DNA isolated from these bones showed that, in fact, only one species of giant moa existed on each island. Size, in this case, had nothing to do with taxonomy; the biggest bones were all from female moa and the smaller bones were from males.
As techniques for isolating ancient DNA improved, the field progressed from asking taxonomic questions to asking more detailed questions about the evolutionary history of populations. DNA sequences could reveal cryptic patterns of local extinctions and long-distance dispersals that were invisible in the fossil record. For example, horses—the same species that humans would eventually domesticate—have been around as a distinct taxonomic lineage for at least a million years. Horses originated in North America and dispersed into Asia across the Bering land bridge, which connected the two continents intermittently during the Pleistocene ice ages. Throughout this period, horses dispersed between North America and Asia several times and in both directions, each time establishing new populations and/or hybridizing with populations that already existed. One might even consider the reestablishment of horses in North America by European colonists as the latest in a long history of local extinctions, dispersals, and recolonizations. Feral horses in North America represent, in essence, an unintentional experiment in rewilding that has been extremely successful.
Ancient DNA can identify genes for traits that don’t exist anymore, such as mammoth-specific hemoglobin, which makes red blood cells that excel at carrying oxygen around large bodies when it is very cold outside. Ancient DNA can also reveal precisely which genetic changes differentiate humans from Neandertals. To summarize, ancient DNA has turned out to be a very powerful technique for learning about the evolutionary processes that shaped existing biodiversity.
The research group that was leading discovery in ancient DNA during the late 1980s and early 1990s was Allan Wilson’s Extinct DNA Study Group at the University of California at Berkeley. This group of scientists was pioneering the development of protocols to recover fragments of DNA from the remains of dead organisms and, importantly, to distinguish authentic ancient DNA from contaminant DNA.
The science fiction potential for ancient DNA was very quick to catch on. In fact, Michael Crichton acknowledges the Extinct DNA Study Group as part of his inspiration for Jurassic Park. And, not long after the 1990 book, science fiction appeared to become scientific fact: several groups (but not the UC Berkeley group) reported sequencing DNA from stingless bees, honeybees, termites, and wood gnats that were tens of millions of years old and even a 120-million-year-old weevil. All of these sequences were generated by extracting ancient DNA from the bodies of insects preserved in amber.
It was too good to be true. In 2013, a team of scientists from the University of Manchester in England performed an experiment to see whether it is possible to extract DNA from bees preserved in copal. Copal, remember, is the precursor to amber and is not entirely fossilized. Copal is therefore much younger than amber. The Manchester team extracted DNA from two copal pieces that contained bees. One of the pieces was around 10,000 years old, and the other was less than sixty years old. They extracted DNA using the latest sample-preparation and DNA extraction techniques. In the end, however, they got nothing—just as we had gotten nothing from our 17-million-year-old piece of amber. They even got nothing from the copal specimen that was less than sixty years old.
This Manchester experiment was the second time that scientists tried to extract ancient DNA from copal-preserved bees. In 1997, a team of researchers from the Natural History Museum of London attempted to repeat—and therefore validate—the fantastical results of the early 1990s. These scientists gathered together a variety of amber and copal pieces from their museum’s collection and attempted to extract and sequence ancient insect DNA. They also failed to recover any authentic ancient insect DNA.
The absence of results is always challenging to interpret. It is possible that, if one were to generate more and more sequence data, a result might eventually manifest. However, the weight of evidence suggests that ancient DNA is not preserved in amber. Not much is known about what happens to insects once they become trapped in tree resin. Although they probably become quickly dehydrated, which is good for DNA preservation, other characteristics of amber make it an unlikely source of well-preserved DNA. Amber is permeable to gases and some liquids, for example, which means that the DNA may not be entirely isolated from the forces that destroy DNA over time. Also, fossilized amber might be subjected to very hot or high-pressure conditions over the course of its lifetime, both of which are terrible for DNA survival.
The failure to replicate these early experiments proves that DNA is not preserved in amber. What was it, then, that these researchers had been able to sequence in the early 1990s?
GETTING DNA FROM FOSSILS WHEN NO DNA IS PRESERVED
Insects are the most likely source of the insect DNA that was recovered from ancient amber in the early 1990s. Insects that are alive in the present day, that is.
Although I left this out above, the researchers at the Natural History Museum in London were sometimes able to isolate insect DNA from the amber from their collections. In fact, it is precisely this result that led them to conclude that amber was not a source of ancient DNA. In designing their experiment, they selected some pieces of amber that contained encapsulated insects and other pieces that did not. This provided a control: if the DNA was from the amber-preserved insects, then the amber with no insects should have no insect DNA. Their results did not support this hypothesis. They were equally likely to recover insect DNA from pieces of amber that contained insects as they were to recover insect DNA from pieces with nothing in them. The insect DNA must have been coming from some source other than the animals preserved within the amber.
This result points to a key challenge of working with ancient DNA. In order to recover DNA from specimens that have very little preserved DNA in them, one needs a very sensitive and powerful method for recovering DNA. But, the more sensitive and powerful the method is, the more likely it is to produce spurious results.
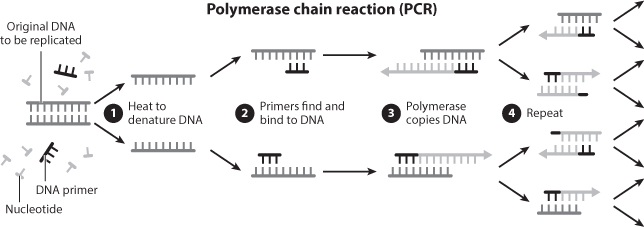
In these experiments, researchers were using a technique called PCR—the polymerase chain reaction—to amplify insect DNA (figure 6). PCR was developed in 1983 by Kary Mullis, who, at the time, was working as a biochemist for a company called Cetus Corporation. DNA-sequencing technologies were making it possible to learn the exact sequence of a fragment of DNA. However, to do so, these technologies required millions of clonal copies of the target fragment. Before PCR, this was achieved by enticing bacteria to capture random fragments of DNA within their genomes. These bacteria were then grown into colonies in which each bacterial cell contained an identical copy of the randomly incorporated DNA fragment: enough copies to sequence. PCR provided a much quicker way to copy DNA and, more importantly, a way to target specific parts of the genome to copy. PCR is now one of the most widely used and essential techniques in molecular biology.
Given its revolutionary implications, PCR is surprisingly simple. To walk through the process, imagine that we want to better understand the genetic differences between domestic and wild chickens. A gene thyroid-stimulating hormone receptor, or TSHR, is thought to have played an important role in chicken domestication by making chickens reproduce more quickly. We propose to use PCR to amplify—make copies of—this gene from DNA extracts of both domestic chickens and from the preserved remains of ancient chickens that lived prior to the time that chickens were domesticated. We will then sequence the results of the PCRs to learn the sequence of this gene and determine whether domestic chickens have different versions than do their wild relatives and pre-domestic ancestors.
Figure 6. The polymerase chain reaction, or PCR. PCR is a common technique in molecular biology that is used to make billions of copies of a DNA sequence by repeatedly heating and cooling DNA in the presence of a DNA-copying enzyme, free nucleotides to build the copied DNA sequences, and DNA primers, which locate the part of the genome to be copied.
First, we need to have some way to target TSHR. We do this by designing two short DNA probes called primers, which will match the sequences of DNA that flank the ends of TSHR. We then make a mixture that contains these primers, the chicken DNA that we already extracted, free nucleotide bases, and a polymerase, which is an enzyme whose job it is to copy DNA. Then we can begin the copying process. We heat the mixture to break the hydrogen bonds that hold together the two strands of DNA. When everything is single-stranded, we cool it back down, which causes the strands to come back together. Because the primers are short and there are a lot of them in the mixture, the first thing that happens is that the two primers find precisely the places in the genome that they were designed to match—the regions flanking TSHR—and form double-stranded DNA with that part of the chicken genome. Finally, the polymerase fills in the missing sequence—the TSHR gene—between the primers, using the single-stranded DNA sequence as a template and the free nucleotides to fill in the missing sequence. When this is complete, the number of copies of TSHR has doubled. To make enough copies to sequence, we repeat the process thirty or forty times over the course of a few hours, eventually generating trillions of identical copies of TSHR.
PCR is extremely sensitive. Theoretically, PCR will work if only one copy of the target DNA sequence exists in the mixture of extracted DNA. On one hand, this is great news for ancient DNA, where very little DNA is expected to have survived. On the other hand, this is a recipe for potential disaster. If DNA can be PCR-amplified from only one fragment of DNA, then it takes only one fragment of contaminating DNA to ruin the experiment. Given this very high sensitivity to contamination, exceptional results, such as DNA from insects preserved in amber millions of years ago, require exceptional proof of authenticity. At the very least, the result should be able to be replicated. In the chicken experiment I described above, identical experiments were performed in ancient DNA laboratories at Durham University in the United Kingdom and at Uppsala University in Sweden. These identical experiments provided identical sequencing results from ancient chicken remains and, therefore, confirmed that the results were real and not due to contamination.
The main source of contamination in ancient DNA research is DNA from organisms that are alive in the present day. DNA is everywhere. It is on the glassware that is used in the lab. It is in the reagents and solutions that are used to extract DNA. It is on the laboratory benches and the floors and walls and ceilings. It is floating in the air in the labs and hallways. Even more problematically, this contaminating modern DNA is in great physical and chemical condition. Whereas pieces of ancient DNA tend to be broken into very small fragments, mostly of fewer than 100 base-pairs strung together (think “cat,” “ant,” “bug”), DNA from living organisms can be millions of base-pairs long (think “supercalifragilisticexpialidocious”). Ancient DNA is also broken. Ancient DNA fragments are often missing bases or have bases that are chemically damaged (“cyt,” “^nt,” “bg”). The polymerase enzymes used in PCR have trouble reading through these damaged sites and end up making mistakes when they copy the sequence (“cut,” “int,” “bog”). Further complicating things, ancient DNA fragments are frequently chemically linked to other pieces of DNA that are present in the DNA extract, forming knotted molecular structures that the polymerase doesn’t recognize as DNA. Because of these problems, the polymerase will preferentially find and make copies of clean, undamaged, freely floating, unbroken, contaminating DNA rather than broken, chemically linked, damaged, ancient DNA. In fact, a single intact fragment of DNA from a living organism can potentially outcompete many hundreds of damaged fragments of ancient DNA during PCR, leading to what looks like, but is not, a real sequence of ancient DNA isolated from a piece of amber. Or from a mammoth bone.
Contamination is not just an idle threat. Contamination comes in many forms, and has played an important role in shaping ancient DNA research. The first and only DNA sequences reported from dinosaurs were (no shock here) contaminants. In fact, many of them were human DNA sequences. Because no one believed dinosaurs were more closely related to mammals than they were to birds or reptiles, and almost nobody believed DNA could be preserved in these very old dinosaur fossils (which are, after all, rocks rather than bones), this result was easily recognized as contamination and rejected.
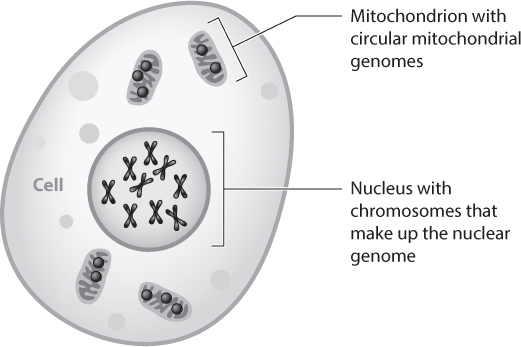
Contamination is often sneakier, however, and this is when it is most dangerous. DNA from modern pigeons (rock doves, the kind that eat leftover fast food and discarded cigarette butts in city centers around the world) somehow contaminated my very first ancient DNA research project, which was to sequence mitochondrial DNA—a type of DNA that is inherited only down the maternal line (figure 7)—from the dodo. The dodo is, as I mentioned, a pigeon, and I was lucky to have spotted the contamination before writing up my conclusions. In this case, spotting the contamination was pretty straightforward. Whereas most of my experiments failed to produce any DNA at all, one particular experiment produced a huge amount of very high quality DNA. This was a dead giveaway that it wasn’t real. I’m still not sure where the contamination came from, but, after that, I started leaving my shoes outside of the ancient DNA lab rather than just covering them up.
Figure 7. Two sources of DNA in our cells. In humans and other eukaryotic animals, each cell contains two types of genome. The nuclear genome, which includes both the autosomes and sex chromosomes, is found in the cell’s nucleus. The mitochondrial genome is found in the mitochondria, which are organelles in the cell cytoplasm. In most eukaryotic animals, mitochondria are inherited only along the maternal line.
In my experience and that of many of my friends and colleagues, there are particular contaminants that pop up from time to time no matter how clean the lab is. DNA sequences of domestic animals and house mice are pretty commonly observed contaminants. This is probably because most of our experiments are designed to amplify DNA from mammals, which, of course, these animals are. Contamination is something we have learned to live with, to expect, and to look for. Because of contamination, we as ancient DNA scientists have developed a healthy wariness of our own data and have set high standards of proof for the authenticity of our results.
This hopefully goes some way to explain the elaborate outfits we wear every time we enter the ancient DNA lab. We are not protecting ourselves from what genetic terrors might be preserved in fossils. Instead, we are protecting whatever DNA might actually be preserved in these fossils from ourselves.
Of course, no matter how careful we are not to contaminate our mammoth bones with our own or any other sources of high-quality DNA, we will likely never find a bone that contains only mammoth DNA. In fact, most of the DNA recovered from a randomly selected mammoth bone will be microbial. Which brings me to our next challenge.
THE SURPRISING DIVERSITY OF DNA IN FOSSILS
Let us assume we find a mammoth bone in Siberia and we want to extract DNA from that bone so that we can sequence the mammoth’s genome. First, we have to protect the bone from contamination. That means we don’t ever touch the bone with our bare hands, because DNA on our hands will get on the surface of the bone, and some of that will be absorbed into the surface layers. We also don’t breathe on the bone, stick it in a bag that hasn’t been sterilized, or allow it to touch other bones. So, we wear gloves and facemasks and hairnets, and we store every sample separately. When we remove a chunk from the sample to take back to the lab (plate 5), we use sterile cutting instruments, work on sterile cutting surfaces, and clean everything with bleach between samples.
When we return to the lab from the field, we do not remove the sample from the sterile bag unless we are in the ancient DNA lab. There, while wearing our sterile and attractive ancient DNA outfits, we smash the bone into powder using sterile smashing equipment and perform DNA extraction using sterile solutions and sterile lab equipment. After we finish the DNA extraction, we have reduced a chunk of mammoth bone to the contents of a tiny, clear tube: an even tinier amount of liquid that looks indistinguishable from water. In that liquid, we should have mammoth DNA.
And bacterial DNA.
And fungal DNA.
And insect and plant and mouse and dog and human and other DNA.
These non-mammoth DNA sequences are, however, not contaminants. More accurately, they are not contaminants in the same sense that my DNA in that sample would be considered a contaminant. The non-mammoth DNA fragments in our DNA extract most likely got into the bone before it was excavated—sometime between when the mammoth died and when we dug up its bone. Bacteria living in the soil, fungi, insects, and plants are all organisms that colonize or grow around bone while it is decaying. Water percolating through the soil will also carry DNA, and this DNA will get into the bone. Even urine carries DNA. A few years ago we showed that sheep DNA can be recovered easily from the same layers of New Zealand soil in which moa DNA is abundant, even though sheep were not introduced into New Zealand until hundreds of years after moa went extinct. A lot of sheep live in New Zealand today. A lot of sheep produce a lot of sheep urine, which leeches through the soil to the deeper layers, commingling with the moa DNA.
Some mammoth bones will have large proportions of mammoth DNA in them relative to the amount of microbial and other exogenous sources of DNA. These are the bones that we prefer to sequence. Unfortunately, it is very difficult to know the ratio of mammoth DNA to other DNA in a sample without going ahead and doing the experiment: extract DNA, sequence it, and see what you get.
Luckily, there are a few general rules about DNA preservation that can be used to guide sample selection. First, cold environments encourage DNA preservation. The chemical processes involved in DNA decay operate more slowly at lower temperatures. Good places to look for bones with well-preserved DNA include the frozen soils (permafrost) of the Arctic and high-altitude caves. Tropical islands are terrible places for DNA preservation, which is bad news for enthusiasts of resurrected dodos (although not all dodos died on Mauritius; some were transported alive to Europe and many of these remains can be found in existing museum collections). Second, ultraviolet light damages DNA. Ultraviolet light causes the same damage to DNA both during life and after death, only dead organisms don’t have the DNA repair mechanisms that keep us from getting terrible skin cancers every time we step into the sunshine. This again points to caves as ideal sources of well-preserved remains, and suggests that remains that are rapidly buried are likely to be better preserved than remains that sit exposed on the surface for many months or years. Third, water is particularly damaging to DNA. Rapid postmortem desiccation and preservation in dry or frozen conditions promotes long-term survival of DNA. Ancient DNA has been recovered from naturally mummified remains of humans, steppe bison, mammoths, and other species. Finally, different tissue types tend to be more or less susceptible to damage and decay. Bone, for example, appears to be a better source of intact DNA than is soft tissue, which perhaps has something to do with the structure of bone matrix or with the bone cells themselves. Hair is another excellent source of well-preserved DNA, as the hydrophobic exterior of the hair shaft limits the amount of water and microbes that might enter the hair and degrade the DNA.
THE TEMPORAL LIMITS OF DNA SURVIVAL
The laws of physics and biochemistry tell us that DNA does not survive forever, even in the best environments for preservation. With that in mind, knowing the age of a sample that we’re considering for a genome-sequencing project is useful in predicting how successful that project will be. While there is no strict rule that states a precise age beyond which DNA will not survive, biochemical modeling suggests an upper limit close to 100,000 years at moderate ambient temperatures. In practice, however, how old we can go varies considerably, and depends on where in the world the sample comes from, what element (hair, tooth, bone, mummified tissue, eggshell) is preserved, and what the preservation history of the sample has been. In warm environments where samples are immersed in water and exposed to UV light, every bit of useful DNA might be destroyed in less than a year. In the Arctic, if a sample is de-fleshed and immediately frozen and then remains underground and frozen from the time of burial to the time of excavation, the DNA within that sample may survive for many hundreds of thousands of years.
It is important to clarify what I mean by “useful” DNA. DNA does not exist as a beautifully preserved and informative molecule on one day and then dissolve into nonexistence overnight on its expiration date. The process of DNA decay includes both the accumulation of chemical damage and the gradual breaking down of the long strands of DNA into smaller and smaller fragments. Once the surviving fragments are shorter than around twenty-five or thirty base-pairs long, they are too short to map to a unique location in the genome and would therefore no longer be useful for genetic research. Fragments of DNA that are one or two base-pairs long may survive for an extremely long time even in very poor environments for preservation, but recovering these would not help us piece together the genome of an extinct species.
I was recently involved in a large international collaboration to sequence the complete genome of an ancient horse—the same kind of horse that runs the Kentucky Derby today, but a very old one. The bone we used was recovered from permafrost soil in the Canadian Arctic. When we found the bone, we knew that it was old. Very, very old. And we were very excited.
In ancient DNA research, it is crucial to know how old the bones are. Knowing the age of each bone provides a way to correlate changes in population size and genetic diversity with environmental changes. For example, horses went locally extinct in North America around 12,000 years ago. As I discussed in chapter 1, the two competing hypotheses to explain horse extinction are that horses fared poorly through the peak of the last ice age around 20,000 years ago or that they were overhunted by humans that arrived in North America after around 14,000 years ago. Knowing that horses were gone by 12,000 years ago is not the same as knowing why horses disappeared. To distinguish between these hypotheses, we need to know when horse populations began to decline. And, to learn this, we have to know the age of each bone.
There are several ways to learn the age of a bone, fossil, or archaeological artifact. In some environmental contexts, such as caves or archaeological sites, these items might be found in a clearly defined layer or stratum within which other items whose age is known are also found. There might be a concentration of fossils that are only found together during a particular time interval, or examples of a prehistoric technology that is only used during a specific period of prehistory. Unfortunately, such layers are not common in permafrost, where most of our horse bones are found.
The age of most permafrost-preserved bones is estimated using a process called radiocarbon dating. Radiocarbon dating measures the ratio of two isotopes of carbon—carbon-14 and carbon-12—in organismal remains and uses this ratio to estimate how long ago the organism died. Carbon-14 is a radioactive isotope of carbon that is created in the atmosphere when cosmic rays collide with nitrogen. Carbon-12 is the normal carbon isotope. Both forms of carbon combine with oxygen to form carbon dioxide, which plants absorb through photosynthesis. Animals then eat the plants, and the carbon from those plants is incorporated into their bones. At any point in time, the ratio of the two forms of carbon is the same in the atmosphere as it is in the organisms living in that atmosphere. Carbon-14 is radioactive and decays at a predictable rate with a half-life of 5,700 years. Because organisms stop taking up carbon after death, we can calculate how long it has been since the organism died based on how much carbon-14 is left in its remains.
Radiocarbon dating is a powerful and pleasantly precise way to estimate the age of permafrost bones. However, the amount of carbon-14 in the atmosphere is very, very small relative to the amount of carbon-12—only about one part per trillion of the carbon in the atmosphere is carbon-14—and the half life of carbon-14 is very short. After around 40,000 years or so, too little carbon-14 will remain in an organism to measure accurately. Radiocarbon dating is useful, therefore, only over this very brief, recent time interval.
Fortunately, there is another way to estimate the age of a permafrost-preserved bone. When volcanoes erupt, they send out a wide fan of very fine dust, which is often referred to as volcanic ash or tephra. The tephra produced by each eruption is unique in its geochemical composition. And, as it turns out, geochemists have developed several ways to learn the ages of these volcanic eruptions. These methods are based on the premise that high heat “resets” the age of the minerals, so that different properties of the minerals can then be measured to estimate when the eruption occurred.
Volcanic tephra is deposited across wide swaths of Alaska and the Yukon Territory and marks eruptions of volcanoes as far to the west as the Aleutian Islands and Alaskan peninsula. When the dust settles, a blanket of white forms on top of the permafrost. As time goes forward, permafrost sediments pile up on top of the layer of volcanic ash, which now clearly delineates fossils buried before the eruption, beneath the tephra, from those buried after the eruption, in permafrost above the tephra. This method is not as precise as radiocarbon dating, but it does provide a means to approximate the ages of bones that are too old to be dated using radiocarbon. This is the method we used to learn the age of our very old horse bone.
HOW OLD IS TOO OLD?
My favorite place to do fieldwork is the Klondike gold-mining region just outside of Dawson City, in Canada’s Yukon Territory. Gold mining, it turns out, is great for ice age paleontology. Most gold miners in the Klondike use a process called placer mining (plate 6). In placer mining, water from the spring snowmelt is collected into holding ponds. After the sun thaws any exposed permafrost, the water is pumped to the active mining site and blasted against the thawed mud. This washes away anything that is not solid ice. The mining then stops for a little while as the warm sun melts away the next layer of frozen mud. Then, the water is turned back on and the freshly melted mud is washed away. This process is repeated until the permafrost is gone and only the gold-bearing gravels remain.
Much to the bemusement of the miners, we are not particularly interested in the gold. We are, however, very interested in the thousands of bones that are unearthed as the permafrost is washed away (plates 7–9). In the Klondike, around 80 percent of these are bones from the extinct steppe bison, about 10 percent are horses, and the rest are mainly mammoths, bears, lions, caribou, wolves, and muskoxen. Crucially, placer mining is slow and methodical, which means that many of these bones can be picked out of the permafrost while they’re still frozen. These bones are impeccably preserved.
We found the really old horse bone in a gold mine near Thistle Creek. The site was special, even among Klondike gold mines. A few years earlier, a team of geologists led by Duane Froese from the University of Alberta discovered that the permafrost near Thistle Creek was very old. In fact, it was the oldest permafrost ever discovered. They knew this because they found a volcanic ash layer called the Gold Run tephra associated with the permafrost mud. The Gold Run tephra was deposited across the central Yukon around 700,000 years ago. So, when we found out that horse bones were preserved within 700,000-year-old permafrost, we could not wait to see whether they contained any horse DNA.
Duane recovered seven bones, all of them larger than those of present-day domestic horses, from the layer of permafrost that was associated with the Gold Run tephra. He made sure that the bones were kept frozen rather than allowed to thaw as they were transported out of the field and into storage. We took subsamples from two of these horse bones for DNA analysis and, to our surprise and delight, were able to recover DNA from both. I repeat: we were able to recover authentic, ancient horse DNA from two 700,000-year-old bones.
The fragments of DNA recovered from these horse bones are the oldest ancient DNA sequences that have been isolated from a specimen whose age is well constrained. However—extraordinary claims require extraordinary proof. Were our results real? We think so. We were extremely careful to make sure the sample was kept frozen and kept away from other samples or other sources of contaminating DNA. The fragments of DNA that we recovered from the bones were short and very badly damaged, as is expected when working with ancient DNA. Our analyses of the sequence data indicated that the old horse was evolutionarily more ancient than living horses. And the results were repeatable. We extracted DNA from these horses in my lab at Oxford and my lab at Penn State, and my colleague Ludovic Orlando and his team at the University of Copenhagen extracted DNA from one of these horse bones several times. The results from all of these extractions were consistent with each other, in terms of both the actual sequences recovered and the damage profile of the recovered DNA. Together, these observations support the authenticity of the very ancient horse DNA.
By the time we finished sequencing ancient horse DNA from this bone, we had generated nearly 12 billion fragments of DNA. We took each of these fragments and tried to match them to the genome sequence of a domestic horse, which had been assembled and published a few years earlier. Around 1 percent of our 12 billion fragments matched different parts of the domestic horse genome, indicating that this tiny component of the DNA recovered from this bone was horse DNA. The other 11.9 billion fragments matched sequences of plant DNA, fungal DNA, bacterial DNA, and other environmental DNA. That is a terrible ratio of horse-to-environmental DNA, and yet we still sequenced the genome of this very ancient horse.
Why did DNA survive in this bone for such an exceptionally long time? We can’t say for certain. The bone was found in the oldest permafrost soil that is known to exist, and the bone probably never thawed between the time of burial and 700,000 years later, when we pulled it out of the frozen ground. Unless older, permanently frozen soils are discovered or fossils are recovered from older ice cores, this may be the age limit of DNA survival in bones.
Exceptional preservation is not limited to the Arctic. Caves have also been found to preserve DNA for a remarkably long time. Most of the Neandertal bones that have been sequenced, for example, were recovered from caves. Recently, DNA was recovered from 300,000-year-old cave bears and a 400,000-year-old hominin from bones preserved in Spanish caves. Environmental stability is known to promote DNA preservation, and caves tend to be consistent in both ambient temperature and humidity, which perhaps explains these examples of exceptionally long-term preservation.
Environmental stability does not, however, seem to be an absolute requirement. We recently pieced together the complete 16,000-base-pair mitochondrial genome of a 100,000-year-old bison bone that was recovered from an ancient lake site in Colorado. The bone belonged to an extinct species of bison called Bison latifrons whose horns spanned an astonishing 2.5 meters in width—five times the width of today’s American bison. The bison bone and the DNA within it somehow survived despite undergoing thousands of seasonal shifts between cold winters and hot summers. The DNA that we recovered from the bone was in terrible condition but, remarkably, remained usable. Would we want to use that particular bison bone as the source of genetic material from which to begin the process of resurrecting Bison latifrons? Not unless we absolutely had to. Less than 0.1 percent of the DNA in the bone was bison DNA, the average fragment length was in the range of thirty base-pairs long, and the sequences were badly damaged. But if this bone was the only bone that we had access to and we really wanted to bring the giant bison back to life, we could use this bone to sequence its genome. We would only get a little bit of bison DNA out at a time, and it would be very expensive. But we would probably, eventually, get the sequence mostly correct.
Fortunately for the mammoth and the passenger pigeon, we don’t have to rely on badly preserved bones with tiny amounts of DNA. Passenger pigeons died out only a century ago, and hundreds of birds are preserved in museum collections around the globe. Well-preserved mammoth remains are even more abundant. If we limit ourselves to the last 40,000 years—which puts us in the range of radiocarbon dating and allows us to know how old the bones we’re working with actually are—there are probably thousands, if not hundreds of thousands, of mammoth remains already in museum and university collections across the world. Most of these are from permafrost deposits, including from the Klondike. Many of these have already been subjects of ancient DNA research, even genome-sequencing projects. We need not be limited, however, to samples sitting on a shelf somewhere at room temperature, subject to faster rates of DNA decay. All we need to do to find an extremely well preserved mammoth bone is get on an airplane, and then a helicopter, and then a boat perhaps, and make our way to the Arctic.