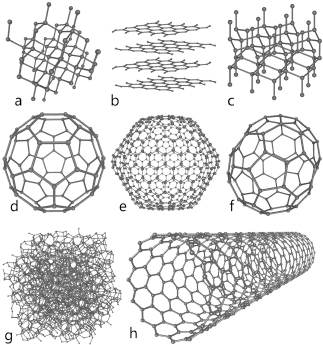
Figure 2.1. A few structures composed of the carbon atom: a) diamond; b) graphite; c) lonsdaleite; d–f) fullerenes; g) amorphous carbon; h) carbon nanotube.
2. THE CHOSEN ATOM
In bodies organised, which true
To type perpetuate themselves.
But through what powers can
Life do this?
Natural or Supernatural?
By properties inherent in
The molecules of which it’s made?…
Or by extraneous, imposed power
A Deus ex Machina force,
Outside the laws of Nature…
This is the problem: this indeed.
—ARTHUR E. NEEDHAM, THE UNIQUENESS OF BIOLOGICAL MATERIALS1
IN THE EARLY MODERN PERIOD RIGHT UP TO THE FIRST DECADES OF the nineteenth century, many biologists were vitalists, believing that the unique behavior, characteristics, and abilities of living things that were not shared by non-living things—including sentience, agency, and the capacity for self-replication—were the result of a nonmaterial, indwelling vital spirit. In the seventeenth century Christina, Queen of Sweden, upon hearing René Descartes insist that organisms are analogous to machines, is said to have retorted by saying of a mechanical clock, “See to it that it produces offspring.”2 Christina’s challenge has yet to be met. Despite extraordinary advances in nano-technology and supramolecular chemistry, no one has assembled a material entity that can mimic the cell’s ability to self-replicate.
In contemplating the astonishing complexity and varied abilities of cells, it is hard to resist the vitalists’ inference that cells are endowed with something beyond the ordinary properties of matter. Especially upon observing activities such as the mating behavior of ciliate protozoa, which mimic in intriguing ways the behavior of many birds and mammals, or watching a white blood cell chasing down bacteria in a blood smear, it’s hard not to imagine some agency or soul bestowing on them their unique abilities. Such abilities are without peer in any known entity in the inanimate realm, including the domain of our own mechanical creations.
But despite its appeal, the inference that there is something at work in living things beyond the laws at play in the inanimate world has a poor track record. The history of biology testifies that from the dawn of organic chemistry in the early nineteenth century to the discovery of the double helix and the molecular biological revolution in the mid-twentieth century, each major advance in knowledge has led to a retreat of vitalist notions. Each new discovery revealed not a vitalist agency gifting cells with this or that unique behavior or characteristic, but rather some extraordinary prior fitness in the properties of matter. In particular, the properties of many of the atoms of the first half of the periodic table are remarkably fit for the assembly of the cell’s chemical constituents and for its physiological functioning.
AS MENTIONED above, in the early nineteenth century, many chemists believed that the unique chemical characteristics of organic compounds derived from some special vital force or agency in the organism. As Lawrence Henderson commented a century later in The Fitness of the Environment, “Many organic substances had been separated from the organism, purified and subjected to the usual experiments of the laboratory… But, as Berzelius [one of the leading chemists of that era] believed, a special vital force had presided over their formation and this, therefore, he supposed to be impossible under any other circumstances.”3
Some chemists at the time imagined the vital agency in very literal terms, like a tiny homunculus in the cell, endowed with the unique ability to assemble atoms into the various complex organic compounds derived from living systems.4 William Prout, for example, a leading physician and chemist in the early nineteenth century and author of the Eighth Bridgewater Treatise (Chemistry, Meteorology, and the Function of Digestion), wrote, “The organic agent… having an apparatus of extreme minuteness, is enabled to operate on each individual molecule separately; and thus, according to the object designed, to exclude some molecules, and to bring others into contact.”5
Although such views seem archaic today, early nineteenth century belief in this form of vitalism is understandable. No one had synthesized an organic compound in the lab. Compared with inorganic compounds, organic compounds were especially fragile and unstable, decomposing rapidly when removed from the body.6 Other known differences included their great variety, diversity,7 and complexity.8 Isaac Asimov summarized some key differences between the inorganic and organic domains thus:
Organic materials are much more fragile and easily damaged than inorganic materials. Water (which is inorganic) can be boiled and the resulting steam heated to a thousand degrees without damage. When the steam is cooled down, water is formed again. If olive oil (which is organic) is heated, it will smoke and burn. After that, it will no longer be olive oil.
You can heat salt (which is inorganic) till it melts and becomes red-hot. Cool it again and it is still salt. If sugar (which is organic) is heated, it will give off vapors, then char and turn black. Cooling will not restore its original nature… Organic substances can be treated with heat or by other methods and converted into inorganic substances. There seemed no way [to the chemists of the early nineteenth century], however, of starting with an inorganic substance and converting to an organic substance.9
The early nineteenth-century chemists did, therefore, have some justification for believing in a unique biological force. The evidence at the time was consistent with the possibility of some mysterious entity in the cell that assembled atoms into organic compounds. Vitalists such as Prout10 accepted that atoms combined together in essentially the same way, according to the same rules, in organic substances as in inorganic. But they believed only living systems could assemble them into organic molecules and actualize the remarkable properties of the substances of the organic domain.
But a major plank in the case for vitalism collapsed in 1828 when, in one of the great breakthroughs of nineteenth-century science, a young German chemist, Friedrich Wöhler, synthesized in his laboratory the compound urea, the major constituent of mammalian urine. It was the first time a chemist was able to synthesize a chemical constituent of a living organism from simple, inorganic compounds. Its synthesis required no vital force and its atomic constituents were combined together in exactly the same way as they would be in an ordinary inorganic compound.
Wöhler obtained urea (CO(NH2)2) by treating inorganic silver cyanate (AgOCN) with another inorganic compound, ammonium chloride (NH4Cl). He wrote triumphantly to his mentor Berzelius, a leading vitalist, “I must tell you that I can make urea without the use of kidneys of any animal, be it man or dog. Ammonium cyanate is urea.”11
As Frances Preston Venable sums up:
It was Wöhler’s brilliant synthesis of urea which finally broke down this barrier, proving the forerunner of many syntheses, and inciting numbers of workers to labor in this lucrative field. It is true that the synthesis had not been made directly out of the elements; but still it was out of substances then regarded as inorganic that he had prepared one of the most interesting and best known of animal products. Of course the dying away of the old belief was slow, but Wöhler’s discovery is commonly pointed to as marking the beginning of organic chemistry as a science.12
In 1845, Hermann Kolbe put another nail in the coffin by synthesizing the organic substance acetic acid from carbon disulfide in the lab.13 After Kolbe’s synthesis, the dam broke, and scientists synthesized more and more organic compounds in the lab. It became clear that at least the basic compounds of living things could be made without some vital agency in the cell.
As knowledge of the chemicals of life grew throughout the nineteenth century, it became ever clearer that not only were the chemical constituents of living things perfectly natural compounds, but also that the carbon atom in conjunction with hydrogen, oxygen, and nitrogen (which make up the bulk of organic substances) possessed a special chemical fitness for the assembly of a vast inventory of complex and diverse organic compounds (acids, sugars, ethers, esters, alcohols) necessary to build complex biochemical systems. As Henderson commented, “The compounds of organic chemistry gradually came to be recognized as different from inorganic substances only in the special characteristics of the elements carbon, hydrogen, and oxygen when in chemical union with one another, just as the compounds of any other elements have their own specific characteristics.”14
Today, a century after Henderson, still no other chemistry is known that can provide such a cornucopia of chemical compounds from which to choose a set of building blocks for a living system and supply all its necessary metabolites. And curiously, the vitalist notion that there was some fundamental difference between the chemicals of life and those of the inanimate realm was retained, even emphasized. But the difference was no longer attributed to a vital supernatural artificer in the cell. It was instead attributed to the unique emergent and natural chemical and physical properties of the carbon atom in combination with hydrogen (H), oxygen (O), and nitrogen (N). Rather than a wonder-working agency in the cell endowing life’s compounds with their unique characteristics, the real wonder-worker was now seen as the unique chemical fitness of certain atoms in the periodic table for life. Agency had been replaced by chemical fitness. A wonder external to nature had been replaced by a wonder immanent in the properties of matter—vitalism by fitness, immediate design by ultimate design, immediate agency by ultimate agency.
It is important to note, however, that although the classic form of vitalism, which postulated a vital agency in the cell to account for life’s chemistry, was abandoned in the nineteenth century as the wonder of carbon chemistry became increasingly apparent, there are still today biological phenomena which are beyond any explanation in terms of the current laws of physics and chemistry. An obvious example is the realm of sentience, mind, and consciousness.15 And whether or not there are laws of nature which apply uniquely to the organic realm remains an open question that we can only hope will be answered by future advances in science.
But whatever future advances of science might reveal regarding the various causal factors at work in living systems, the core claim defended in the chapters ahead is based on an assessment of the scientifically established properties of matter and laws of nature accepted by all biologists today. That there is a profound prior fitness in nature which enabled the actualization of the canonical cell as it exists on Earth is a claim independent of whatever might have been the direct causal factors responsible for the assembly of the first living cell on Earth or the exact physical and chemical steps by which the miracle was actualized. Whether these factors were Darwinian, Lamarckian, vitalistic, or some other is a debate of great interest but largely peripheral to the focus of this book.
The Infinite Inventory
THE DEVELOPMENT of organic chemistry is one of the great episodes in the history of science, and was described by Henderson as “one of the greatest achievements of the nineteenth century.”16 Others concur. Jan Mulder entitled a paper reviewing its development as “Looking Back in Wonder.”17 Many other authors, including Asimov18 and Alfred Russel Wallace, co-founder with Charles Darwin of the theory of evolution by natural selection, have waxed lyrical about the wondrous universe of carbon chemistry.
By the beginning of the twentieth century, more than 100,000 organic compounds had been documented.19 And all the basic compounds of living organisms—the twenty common amino acids used in proteins and the four nucleotides used in DNA, as well as many of the sugars and fats and fatty acids found in living organisms—had been synthesized in the lab.
Of all the elements, carbon stands alone in its ability to form a vast array of complex organic compounds with diverse chemical and physical properties. Indeed, the number of known carbon compounds is currently estimated to be close to ten million, greater than the total of all other non-carbon compounds combined and much larger than Henderson’s estimate from a century ago.
And aside from molecules that include carbon, there are many molecules that contain only carbon. Carbon makes up substances as diverse as coal, diamond (the hardest mineral known), and graphite (one of the softest), as well as complex structures such as fullerenes and nano-tubes. In recent decades chemists announced the discovery of another carbon compound, graphene, which consists of a flat monolayer of carbon atoms packed tightly into a two-dimensional honeycomb arrangement. Its most remarkable characteristic is its strength: it is one hundred times stronger than an equivalent monolayer of steel. Graphene conducts electricity as well as copper does, and conducts heat better than can any known material.
However, the diversity of chemical forms that can be assembled using carbon alone pales against the fantastic diversity of compounds that can be assembled when carbon combines with other atoms.
Carbon and hydrogen combinations form the universe of hydrocarbons. Some hydrocarbons are long, chain-like molecules, such as pentane and butene. Others contain cyclic or ring-like formations, such as benzene. And it is not just the number of chemical structures that dazzles, but also the variety and diversity of properties. Plastic milk jugs, DVD discs, oils, petroleum, kerosene, and naphthalene (moth balls) are all combinations of carbon and hydrogen atoms.
Combining carbon with both hydrogen and oxygen opens another universe of compounds, including alcohols such as ethanol and propanol, aldehydes, ketones, and the carboxylic acids. This combination also creates the vast variety of fatty acids, composed of a long hydrocarbon chain that is attached to a carboxylic acid group at one end. Carbon, hydrogen, and oxygen are also responsible for the sugars, including glucose and fructose. Beyond that, this triad creates cellulose (the hard substance of wood), beeswax, vinegar, and formic acid. All of these belong to this group of carbon compounds.
|
Figure 2.1. A few structures composed of the carbon atom: a) diamond; b) graphite; c) lonsdaleite; d–f) fullerenes; g) amorphous carbon; h) carbon nanotube. |
Throwing nitrogen into the mix leads to a further multiplicity of compounds, including the building blocks of proteins: amino acids. It also creates a set of cyclic compounds known as the nitrogenous bases, some of which are important building blocks of DNA. This combination is found in items as diverse as dyes, antibiotics, explosives, caffeine, and urine.
By the late nineteenth century and the beginning of the twentieth, when the full wonder and uniqueness of carbon’s lavish inventory had been established, Alfred Russel Wallace described the sheer number and diversity of the denizens of this unique domain in both of his works on natural theology, Man’s Place in the Universe and The World of Life: A Manifestation of Creative Power, Directive Mind and Ultimate Purpose. In the first of these, he wrote:
The chemical compounds of carbon are far more numerous than those of all the other chemical elements combined... And the marvel is still further increased when we consider that the innumerable diverse substances produced by plants and animals are all formed out of the same three or four elements. Such are the endless variety of organic acids, from prussic acid to those of the various fruits; the many kinds of sugars, gums, and starches; the number of different kinds of oil, wax, etc.; the variety of essential oils which are mostly forms of turpentines, with such substances as camphor, resins, caoutchouc [natural rubber], and gutta-percha; and the extensive series of vegetable alkaloids, such as nicotine from tobacco, morphine from opium, strychnine, curarine, and other poisons; quinine, belladonna, and similar medicinal alkaloids… all alike consisting solely of the four common elements from which almost our whole organism is built up. If this were not indisputably proved, it would scarcely be credited.20
The total number and diversity of possible chemical structures that may be constructed out of carbon, oxygen, hydrogen, and nitrogen staggers the imagination. Together, these elements form what is in effect a universal chemical constructor kit ideally suited for the construction of the myriads of chemical compounds the cell employs. The need for such a vast inventory of organic compounds is indicated to a degree by published metabolic pathway charts. The charts show the maze of chemical pathways and the huge number of different compounds which undergo chemical transformations in the course of metabolism in a typical cell.
King Carbon
GIVEN THE unparalleled fecundity of the carbon universe, it is somewhat curious that in many ways carbon is not a very prepossessing substance. Think of a lump of coal or a piece of graphite or a heap of soot. Diamond has allure, but on the whole, in most of its elemental forms, carbon does not impress. Its relative un-reactivity21 adds to the impression that it is mundane compared with other more spectacular and reactive atoms like sodium or oxygen. As chemist Peter Atkins comments, carbon seems in terms of reactivity a “particularly mediocre” atom and “easygoing in the liaisons it forms.”22
But carbon does have chemical fecundity, which elevates it into a category all its own, creating the vast array of chemical combinations described above. For this reason, Atkins termed carbon “the King of the Periodic Kingdom.”23 Certainly without the vast inventory of complex molecules of utterly diverse chemical properties gifted to us via the unique properties of this king of atoms, there would be no organic plenitude to satisfy the complex metabolic needs of the cell. In all probability, there would be no chemical life in the universe. Atkins goes so far as to say that the “property we term ‘life’ stems almost in its entirety” from the region of the kingdom containing carbon.24
Carbon-Carbon Bonds: Of all the atoms of the periodic kingdom, including carbon’s three associates which make up the substance of organic compounds—hydrogen (H), oxygen (O), and nitrogen (N), only carbon can bond firmly with itself to form chains of atoms (i.e., C-C-C-C) of almost unlimited length. In this ability, carbon is unique. No other atom in ambient conditions, not oxygen, nitrogen, hydrogen, or silicon, possesses this ability to anything like the same degree as carbon.
More than anything, it is the stability of carbon-carbon bonding that enables organic compounds to grow to almost unlimited size and complexity. In the case of organic molecules containing carbon, as Asimov put it, “Carbon atoms can join one another to form long chains or numerous rings and then join with other kinds of atoms as well. Very large molecules may be formed in this way without becoming too rickety to exist. It is not at all unusual for an organic molecule to contain a million atoms.”25 Large molecules even remotely as complex as proteins or other macromolecules are simply unknown outside the domain of organic chemistry. Many authors have stressed this. As Primo Levi puts it, “Carbon, in fact, is a singular element: it is the only element that can bind itself in long stable chains without a great expense of energy, and for life on earth (the only one we know so far) precisely long chains are required. Therefore carbon is the key element of living substance.”26
Tetravalency: Carbon possesses another element of fitness: it is tetravalent, meaning it can form four electron-sharing chemical bonds with other atoms including itself. By way of comparison, nitrogen can only form three such bonds with other atoms, oxygen two bonds, and hydrogen one. As Arthur Needham comments, “The four bonds of each carbon atom are directed towards the corners of an imaginary tetrahedron, with the carbon atom as its centre, so that by bonding with other carbon atoms an indefinitely extended three-dimensional fabric is possible, similar to that of water… The basis of bioplasm is essentially a fabric of this kind.”27
Its tetravalency further contributes to carbon’s unique fitness for the formation of the vast inventory of organic molecules.
Multiple bonds: A third element of fitness which contributes to the limitless fecundity of the organic universe is carbon’s ability to form multiple, stable bonds with itself and other atoms. This is a consequence of carbon’s relatively small atomic radius, which means the bond distances are short and thus relatively strong.28 The other small, nonmetal atoms in period two, including two of carbon’s partners, oxygen and nitrogen, also share this capacity.
Carbon can form single, double, and triple bonds with other atoms, including itself. Nitrogen can form single, double, and triple bonds, and oxygen can form single and double bonds. The nonmetals right below them in the periodic table (silicon, phosphorus, and sulfur) form such bonds less readily because their larger atomic radii render multiple bonds less stable.29
The Right Strength: The strength of the bonds that link the carbon atoms with other carbon atoms and with the other atoms of organic chemistry—mainly hydrogen, oxygen, and nitrogen—has to be commensurate with their chemical manipulation by the molecular machinery in the cell; otherwise the above ensemble of fitness would be to no avail. Fortunately, the strength of these chemical bonds, and their energy levels—which play a major role in determining the strength of the bonds—are just what they need to be.
To understand why the energy levels of the chemical bonds in organic compounds are indeed in the right range for biochemical manipulations, consider briefly how the cell’s molecular machines carry out chemical reactions. Basically, a combination of two factors are involved. One involves using the energy of molecular collisions to weaken chemical bonds, and the other, specific conformational movements in an enzyme molecule, which strain a particular bond in a particular substrate molecule, lowering the energy level of the bond. In chemical jargon, it decreases the activation barrier, making the bond weaker and easier to break.30
The need for the decrease in the activation barrier is real, because at ambient temperatures the energy imparted by molecular collisions is insufficient to overcome the energy barriers of most organic bonds.31 (And this is why the organic compounds which make up the substances of the body remain chemically stable for relatively long periods of time). By reducing the activation barriers, many more molecular collisions have sufficient energy to break the bonds. Although protein conformational changes involve energy levels significantly less than those of a covalent bond,32 about one tenth, fortunately they are still sufficient in the ambient temperature range to significantly strain particular bonds to lower their activation energy to levels that can be broken by less energetic but more frequent molecular collisions.
If organic bonds were substantially stronger in the ambient temperature range, say as strong as in many inorganic compounds which may be two to three times as strong33 (and which can only be broken by heating to very high temperatures), protein movements could not significantly weaken particular bonds, i.e., decrease the activation barrier for particular reactions. Consequently, the sorts of controlled chemical reactions carried out in living cells would be greatly constrained. Moreover, not only would proteins be unable to exert sufficient conformational strain to significantly weaken particular bonds, but molecular collisions in the ambient temperature range would only very rarely impart sufficient energy to overcome energy barriers and cause bonds to break. On the other hand, if organic bonds were substantially weaker in the ambient temperature range, disruption via molecular collisions would dominate and no controlled chemistry would be possible.
It turns out that the actual strength of organic bonds, as with so many other examples of the fitness of nature for life, is situated in a Goldilocks zone, neither too strong nor too weak, but just right. If the bonds were stronger or weaker by a single order of magnitude, the controlled chemistry of the cell would very likely be impossible. And it is surely an arresting fact, testimony to the prior fitness of nature for carbon-based life, that this Goldilocks zone represents an inconceivably tiny band in the vast spread of energy levels in the cosmos. (Gravity, for example, is at least 1036 times weaker than the strong nuclear force.34)
In short, biochemistry is only possible because carbon compounds in the ambient temperature range are, as described by Needham, uniquely “metastable.”35 As he points out, while carbon compounds are relatively stable in this temperature range and can persist without undergoing chemical change for long periods in the cell, they are “notable for lability as much as for their stability. Few remain unchanged when heated above 300°C, and most are gaseous at that temperature, if not already decomposed. As in so many other respects, carbon seems to have the best of both worlds, in fact, combining stability with lability, momentum with inertia.”36 As Henderson put it in Fitness:
Not less valuable for the organism than the multiplicity of organic substances, and the diversity of their properties, are the great variety of chemical changes which they can undergo, and that characteristic instability which renders such great complexity of chemical behavior easily attainable. In short, organic substances are uniquely fitted not only to provide complexity of structure to the organism, but also, through their instability and manifold transformations to endow it with diverse chemical activities, with complexity of physiological function.37
Similar Affinity: Many authors also have stressed another characteristic of carbon bonds: the energy levels of carbon bonds do not differ much from one partner element to the next. As N. V. Sidgwick notes in his classic The Chemical Elements and Their Compounds, “The affinity of carbon for the most diverse elements, and especially for itself, for hydrogen, nitrogen, oxygen, and the halogens, does not differ very greatly: so that even the most diverse derivatives need not vary much in energy content, that is, thermodynamic stability.”38 Robert E. D. Clark expanded on the same point, writing that carbon “is a friend of all. Its bond energies with hydrogen, chlorine, nitrogen, oxygen or even another carbon differ little. No other atom is like it.”39 Kevin W. Plaxco and Michael Gross concur, and elaborate in their well-known Astrobiology:
Carbon presents a fairly level playing field in which nature can shuffle around carbon-carbon, carbon-nitrogen, and carbon-oxygen single and double bonds without paying too great a cost to convert any one of these into another… Given all this, it’s no wonder that on the order of ten million unique carbon compounds have been described by chemists, which is as many as all of the described non-carbon-containing compounds put together.40
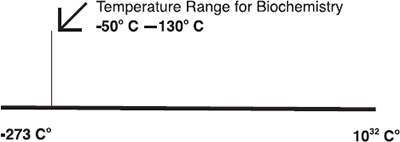
The Right Temperature Range: The necessity for metastable organic compounds to enable the controlled chemistry in the cell has a further intriguing consequence; it constrains the life-friendly temperature range to an extraordinarily narrow band within the immensity of the total range of temperatures in the cosmos.
The upper temperature limit for life is not much above 100°C.41 This is because of the characteristic instability of most organics as temperatures rise beyond that point. Stanley Miller and Leslie E. Orgel noted this in their book The Origins of Life on the Earth.42 The key amino acid alanine, for example, has a half-life of 20 billion years at 0°C, three billion years at 25°C, but only ten years at 150°C, a decrease of more than a billion-fold. And alanine is not exceptional.43 As I pointed out in Nature’s Destiny, “Many vitamins, including vitamin C, folic acid, and some of the other B vitamins—B1 and B6, for example—are rapidly broken down above 100°C.”44 One report in the journal Nature45 showed that the half-life of many of the key organic compounds used by living things—including the amino acids used in proteins, the bases used in DNA, and the adenosine triphosphate (ATP) used for energy metabolism in cells—decompose at rates too fast to measure, or have half-lives on the order of minutes or seconds, at 250°C.46
The lower level for controlled biochemistry has not been ascertained. However, it is known that some organisms can function at temperatures as low as -20°C, below which cell vitrification causes metabolism to cease.47 Whether life could exist at even lower temperatures is not known because no detailed studies have been carried out on the biochemistry of cells at temperatures below -20°C in fluids that are liquid at very low subzero temperatures. But being generous and allowing for a “slow” biochemistry at -50°C (in some fluid other than water) and an upper limit of, say, 130°C, the temperature range fit for biochemistry would still occupy only an infinitesimal fraction of the vast range of temperatures in the universe.48
That by itself is striking. But there’s something else. This temperature range just so happens to be almost the same as the temperature range in which water is a liquid in ambient conditions on Earth,49 surely one of the most extraordinary and consequential bio-friendly coincidences in nature. For if these two independent ranges didn’t happen to overlap, there would be, in all probability, no carbon-based life on Earth or indeed anywhere in the universe.
|
Figure 2.2. Cosmic temperatures, from the temperature of absolute zero to the temperature of the Big Bang. |
Multiple Fitness
IN SUM, carbon is fit in many different ways for the assembly of the complex molecules of life:
1. It forms stable bonds with itself.
2. It forms up to four bonds, being tetravalent.
3. It forms multiple bonds with itself and other atoms.
4. The energy levels of carbon bonds are just right for biochemical manipulation in the ambient temperature range—not too strong and not too weak—described as being “metastable.”
5. The energy levels of the covalent bonds that carbon forms with its other nonmetal partners in organic compounds are similar.
6. The metastability of carbon compounds is in the same temperature range that water is a liquid.
And there is yet another element of fitness which will be discussed in the next chapter: the unique directional nature of the bonds carbon forms with other atoms in organic compounds, which, we will see, plays a vital role in the assembly of complex macromolecules of defined 3-D shapes.
That the carbon atom is uniquely fit for the chemistry of life is not the view of an esoteric minority of researchers or of any special pleading. The peerless fitness of the carbon atom to build a universe of diverse chemicals and fantastically complex macromolecules like proteins and DNA has been recognized by the majority of authors and researchers cognizant of the facts. This has been the case for more than a century.50
By the end of the nineteenth century, all attempts to account for the very remarkable chemical properties of the organic realm in terms of a mysterious “vital force” had been abandoned. Even advocates of design such as Alfred Russel Wallace saw chemical design as inherent in nature and sufficient without augmentation by some mystical vitalistic agency.
While there may be other forms of chemical life, perhaps based on boron or silicon (for which there is at present no empirical evidence), what seems not in doubt is the supreme fitness of carbon for any chemical life form analogous to that on Earth. As Gross and Plaxco confess in Astrobiology, “In the end there may very well be only a single element—carbon... the basis of all life on Earth—that is able to support the complex chemistry presumably required to create a self-replicating chemical system.”51
Finally, what is particularly striking about the properties of the carbon atom is that they appear to be fine-tuned in several different but complementary ways to generate the plenitude of compounds uniquely useful to life. Such a suite of properties, all seemingly arranged to generate a vast inventory of molecules ideal for the biochemistry of living cells, conveys a powerful impression of contrivance. More than a century ago Wallace expressed, in his World of Life:
We see, therefore, that carbon is perhaps the most unique, in its physical and chemical properties, of the whole series of the elements, and so far as the evidence points, it seems to exist for the one purpose of rendering the development of organized life a possibility. It further appears that its unique chemical properties, in combination with those of the other elements which constitute protoplasm, have enabled the various forms of life to produce that almost infinite variety of substances adapted for man’s use and enjoyment, and especially to serve the purposes of his ever-advancing research into the secrets of the universe.52
It turns out that Wallace, peering into the biochemical basis of life, saw what has now become even clearer. Many have believed (and many still do believe) that Darwin drove teleology out of biology forever. But more than a century and half of scientific research since Darwin has shown that the fitness of nature for life on Earth, exemplified so wonderfully in the chosen atom, points irresistibly to purpose and design.