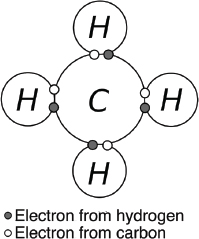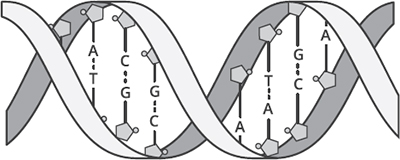
Figure 3.1. DNA’s double helix. The twisting ribbons are the phosphate chains. The letters are the pairings of nucleotide bases—adenine (A) and thymine (T), and cytosine (C) and guanine (G). The pentagons attached to the letters are the sugars.
3. THE DOUBLE HELIX
Twenty angstrom units in diameter, seventy-nine billionths of an inch. Two chains twining coaxially, clockwise, one up the other down, a complete turn of the screw in 34 angstroms. The bases flat in their pairs in the middle, 3.4 angstroms and a tenth of a revolution separating a pair from the one above or below. The chains held by the pairing closer to each other around the circumference one way than the other, by an eighth of a turn, one groove up the outside narrow, and other wide. A melody for the eye of the intellect, with not a note wasted.
—HORACE JUDSON, THE EIGHTH DAY OF CREATION1
CARBON’S UNIQUE CHEMISTRY, MANIFEST IN THE SHEER NUMBER and diversity of its compounds, is ideally fit to provide the cell with a fantastic inventory of small molecular building blocks—sugars, amino acids, nucleotide bases, fats, steroids, and so forth. Yet this abundance cannot by itself explain all of the seemingly miraculous abilities of cells, such as how enzymatic catalysis is carried out or how cells transmit genetic information to their two daughter cells in cell division. These phenomena were still complete mysteries in the early twentieth century. And for many researchers at the time, it seemed these abilities might not be explicable in terms of the laws of physics and chemistry that applied in the inorganic world or the laboratory. Consequently, vitalist notions still found some support among many biologists.
James Watson commented in Molecular Biology of the Gene:
Through the first quarter of this century, a strong feeling existed in many biological and chemical laboratories that some vital force outside the laws of chemistry differentiated between the animate and the inanimate. Part of the reason for the persistence of this “vitalism” was that the success of the biologically orientated chemists (now usually called biochemists) was limited. Although the techniques of the organic chemists were sufficient to work out the structures of relatively small molecules like glucose… there was increasing awareness that many of the most important molecules in the cell were very large—the so-called macromolecules—too large to be pursued by even the best of organic chemists.2
And, as Watson further noted, even the demonstration by James B. Sumner in 1926 that the enzyme urease was a protein and could be crystallized in the lab “did not dispel the general aura of mystery about proteins.”3 These were still undecipherable by the techniques available at the time, so “it was still possible, as late as 1940, for some chemists to believe that these molecules would eventually be shown to have features unique to living systems.”4 The elusive genetic material was widely considered to be made up of proteins (not DNA), which were known to be associated with chromosomes in the cell nucleus. DNA molecules were also known to be a constituent of the chromosomes, but “these were thought to be relatively small and incapable of carrying sufficient genetic information.”5 And as Watson concludes, “The feeling was often expressed that something unique about the three-dimensional organization of the cell gave it its living feature… that some new natural laws, as important as the cell theory or the theory of evolution, would have to be discovered before the essence of life could be understood.”6
Max Delbrück, one of the founding fathers of molecular biology and leader of the celebrated Phage Group7 (whose work led to the discovery that bacteriophage DNA carries genetic information and specifies the assembly of phage capsids in the bacterial cell), commented to Horace Judson during a conversation that Judson reported in his classic The Eighth Day of Creation:
Both proteins and nucleic acids were hopelessly inadequately characterized, in those days. Proteins were characterized a little more, because we knew that there were twenty essential amino acids—or twenty-odd amino acids: one didn’t, after all, know really how many. But whether proteins were built in a regular repetitive way or in a very specific way was still very unknown in the forties. And similarly, DNA—one knew it was a fibrous molecule, one didn’t really know how the nucleotides went in together, at this time, or even whether it had branches in it or not.8
Ideas on the three-dimensional structure of proteins were no less ill defined. Scientists agreed that proteins were big molecules, or macromolecules, probably composed of linear chains of amino acids, but didn’t know how their primary sequences were determined, how they were synthesized, and how they arrived at their native three-dimensional conformations.9 So little was known about proteins or DNA in the 1930s and ’40s that, as mentioned above, most biochemists thought that proteins, rather than nucleic acids, played the primary role in heredity and made up the material structure of the gene.10
In short, there was every reason, as in the early nineteenth century, for vitalistic speculation. As Delbrück pointed out in the 1930s, “Genes at that time were algebraic units of the combinatorial science of genetics, and it was anything but clear that these units were molecules analyzable in terms of structural chemistry. They could have turned out to be submicroscopic steady-state systems, or they could have turned out to be something unanalyzable in terms of chemistry.”11
As Francis Crick recalls about the years leading up to the molecular biological revolution:
Looking back, what was really striking about that period was not, as we take it now rather obviously, that chemistry and physical chemistry, and the physics associated with those subjects, are essential to understanding biology at the molecular level. Not everybody agreed with that, in particular Max Delbrück, who was a physicist who went into biology.… He was hoping to discover new laws of physics which emerged when he looked at these extremely mysterious biological processes of how you get replication, which in those days seemed utterly baffling.12
Erwin Schrödinger also alluded to Delbrück’s quasi-vitalist hope that there might be special laws of physics to account for heredity: “From Delbrück’s general picture of the hereditary substance it emerges that living matter, while not eluding the ‘laws of physics’ as established up to date, is likely to involve ‘other laws of physics’ hitherto unknown, which, however, once they have been revealed, will form just as integral part of this science as the former.”13
In sum, despite the fitness of the carbon atom for the diverse organics required for the cell, this knowledge left unsettled how higher-order phenomena like enzymatic catalysis and heredity were carried out in the cell. There was a gap, and it was not clear how it might be closed.
After Watson and Crick
BUT DELBRÜCK’S hope that new laws might be discovered was not to be. The mid-twentieth-century molecular biological revolution revealed that no laws unique to biology were necessary to account for enzymatic catalysis or the phenomenon of heredity.
It turned out that what biochemist Jacques Monod referred to as the “demoniacal functions”14 of enzymes as well as the storage and replication of genetic information could be largely explained by the same laws of physics and chemistry that applied in the inanimate realm; that something as mundane as molecular shape—the exact positioning of atoms in space—could achieve what was previously attributed to something beyond ordinary chemistry.
During the 1940s and 1950s, knowledge of the molecular structure of the core macromolecules of life increased dramatically due to a series of revolutionary advances brought about by the application of powerful new technologies such as X-ray crystallography and electron microscopy. The advances finally clarified the basic structure and function of the key macromolecules in the cell.
And they revealed something quite astonishing: the atomic constituents of the complex macromolecules in the cell could be arranged in unique, highly specific 3-D conformations, and it was this that enabled macromolecules such as proteins and DNA to carry out highly specific biochemical, enzymatic, and genetic functions previously attributed to mysterious vital forces.
|
Figure 3.1. DNA’s double helix. The twisting ribbons are the phosphate chains. The letters are the pairings of nucleotide bases—adenine (A) and thymine (T), and cytosine (C) and guanine (G). The pentagons attached to the letters are the sugars. |
The protein structures brought to light by X-ray crystallography were the stuff of science fiction. These were objects consisting of up to thousands of atoms arranged into highly specific 3-D arrangements, stranger and more beautiful and more dauntingly complex than any previously conceived aggregate of matter. Max Perutz, quoting J. C. Kendrew’s earlier paper, commented on the 3-D conformation of the atoms in myoglobin, the first protein to have its structure determined in atomic detail: “Perhaps the most remarkable features of the molecule are its complexity and its lack of symmetry. The arrangement seems to be almost totally lacking in the kind of regularities which one instinctively anticipates, and it is more complicated than has been predicated by any theory of protein structure.”15
Rosalind Franklin’s X-ray diffraction work carried out at my alma mater, King’s College in London, led to the discovery that the atoms in DNA are deployed in the form of a double helix. Although the double helix is a far more regular structure than the bewildering forest of atoms in proteins such as myoglobin or cytochrome, the structure nonetheless revealed again how molecules held in specific 3-D conformations could perform very specific functions, in this case the transmission of genetic information.
In the 1960s, shortly after the glory days of the double helix, my doctoral supervisor at King’s College, Henry Arnstein, often referred to the shock he and many other biochemists felt when it became clear that all the atoms in a complex protein could be maintained in precise spatial conformations, and that this specificity of atomic arrangement was key to enabling matter to carry out the seemingly miraculous chemical activities within the cell. A veil had been drawn aside. This was the molecular secret of life.
Judson, in an insightful foreword to the first edition of The Eighth Day of Creation, emphasized that a deeper understanding of high specificity was key to ushering in the revolution:
In the transformation of biology, the great underlying shift of view was the development of the concept of biological specificity. In the mid-thirties, biologists and biochemists certainly spoke of specificity. They had to do so, for many of the phenomena they dealt with—genes (whatever they were in substance), enzymes and antibodies (known to be protein)—were highly specific in action. Yet specificity was really a term almost empty of meaning.… Forty years later, biological specificity is richly stuffed with meaning.16
Although the discovery of the structure of DNA was one of the seminal discoveries of twentieth-century science, the double helix is certainly not the secret of life that Crick claimed it to be in 1953. Nor is any other molecule. Indeed, it is possible to conceive of carbon-based life without DNA, one that uses another self-replicating genetic polymer, perhaps closely related to DNA but built from different nucleotides or perhaps a very different type of polymer built from quite different basic units. A group of researchers at the University of Florida and another group in Oxford working in the field of artificial life are now busy redesigning the genetic code17 and building analogues of DNA.18 It may be that these alternatives are inferior in one way or another to DNA; nonetheless, DNA may turn out to be just one of a number of complex polymers capable of performing the genetic function in carbon-based life forms.
What is unarguable, however, is that the functions of all the macromolecules in current biological systems on Earth depend critically on the ability to deploy multiple atoms (sometimes thousands) in very specific irregular spatial conformations. And one can assume that even artificial life, if we ever invent it—and alien life, if it exists—also will depend on highly specific 3-D molecular conformations of their chemical components.
No chemical life that we can conceive of (and many definitions are given in the literature19) would be feasible without complex molecular machines that can carry out defined tasks. And any sort of molecular machines that can carry out specific biochemical functions would necessarily depend on highly precise and stable 3-D arrangements of atoms. For instance, enzymes catalyze life-essential processes by binding to specific substrates, increasing the rate of conversion to an end product by thousands, or even millions, of times per second.20 No enzyme could manage any such task unless the atoms around the active site were deployed in very exact spatial arrangements to bind the substrate.
THE ARRANGEMENT of atoms in complex bio-macromolecules into highly specific 3-D conformations depends on two types of chemical bonds, strong or covalent bonds (discussed in the previous chapter) and another set of quite different bonds, what are called weak bonds.
Covalent bonds form when atoms share electrons to complete their outer electron shells. Familiar compounds in which the atoms are linked by covalent bonds are carbon dioxide (CO2), water (H2O), ammonia (NH3), and methane (CH4) or marsh gas, shown in Figure 3.2. (For simplicity’s sake I’m describing the Lewis Theory for chemical bonds, which is precise enough for our purposes here. However, the most recent scientific view, which gives a more detailed explanation of chemical bonds, is the Molecular Orbital Theory.)
|
Figure 3.2. Covalent bonds in methane. |
In organic compounds, all the bonds between the constituent atoms, such as C-H, C-O, C-N, and N-O bonds, are strong covalent electron-sharing bonds. The crucial feature of covalent bonds is that they are spatially constrained by the existence of other bonds in the molecule. In other words, the bonds are directional. As Peter Atkins explains about covalent bonding, “The ability of an atom partially to release electrons to form a covalent bond in one direction will affect its ability to release them in a different direction. As a result, the arrangement of atoms in a molecule has a fixed, characteristic geometry… covalent compounds… are discrete, often small groupings of atoms… with characteristic shapes.”21
The fact that the bonds in the molecular building blocks of the cell’s key macromolecules are directional and spatially constrained is of very great consequence. Why? Because a complex macromolecule in which all the atoms must be deployed in stable, specific spatial arrangements, to serve particular biological functions, cannot be assembled from subunits in which the bonds are not directional and spatially constrained.
Ionic bonds, the other type of strong chemical bond, are also strong, but they are not directional and spatially constrained. Ionic bonds involve the donation or acceptance of electrons between two atoms, leaving one atom (the electron donator) positively charged and the other atom (the electron acceptor) negatively charged. Thus, in the formation of salt (NaCl), the sodium atom donates an electron to a chlorine atom, conferring a positive charge on the sodium atom (Na+) and a negative charge on the chlorine (Cl–). Ionic bonds bind the atoms of most inorganic compounds in the mineral world.22 But although ionic bonds are strong—generally more than twice the energy of the covalent bonds that carbon makes with hydrogen, oxygen, and nitrogen—they are non-directional, a fatal defect for the assembly of biomolecules with defined shapes. As Robert E. D. Clark noted, non-directionality is a fundamental reason why organisms could not use ionic bonds:
The attraction between charged atoms is completely devoid of any directive quality: the atoms are only concerned to maintain the same distance between one another and, apart from this, they care nothing for organization.
To make this point clear we may imagine a kind of Alice-in-Wonderland teapot, on the atomic scale. We will consider the teapot to be a “compound” of pot, spout, lid and handle. But when we put the lid, spout and handle on the pot we find, to our consternation, that they do not stay where we put them, but proceed to slip round the pot, beneath it and above!… In short, considered as an organized structure, the teapot fails to fulfill a useful function—its parts might just have well have been joined together by string!
Fantastic as it sounds, the analogy gives us a passable picture of how atoms held together by electric charges [ionic bonds] behave.23
As Atkins points out, the distinction between these two types of strong bonds, ionic (non-directional) and covalent (directional), corresponds with the fundamental division between the inorganic domain and the organic domain, with the latter literally resting on the former:
In general, molecular compounds [made up of covalently bonded atoms] are the soft face of nature, and ionic compounds [inorganic] are the hard. Few distinctions make this clearer than those between the soft face of the Earth—its rivers, its air, its grass, its forests, all of which are molecular—and the harsh substructures of the landscape, which are largely ionic. This is why the upper triangle of the Eastern Rectangle [in the periodic table] is so important to the existence of life, and why all the rest of the kingdom is so important in the formation of a stable, solid platform.24
That the periodic table of elements should contain, in the region Atkins calls “the upper triangle of the Eastern Rectangle,” a set of atoms including carbon (C), nitrogen (N), oxygen (O), and hydrogen (H), as well as phosphorus (P) and sulfur (S), possessing bonds of just the right strength for chemical manipulation in the cell as well as the crucial directional property, is surely indicative of a deep fitness in nature for carbon-based life.
And it is worth noting that these same atoms also form covalent directional bonds (more properly termed coordinate bonds) with a special class of metals—the transitional metals, including notably iron and copper. The directional bonding with the transitional metal atoms is also of vital importance, because it enables the metal atoms to be bound to proteins in unique molecular geometries, which in turn confers on the metal-protein complex many unique chemical properties that underlie specific enzymatic or other activities in the cell.
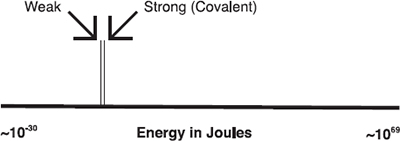
ALTHOUGH THE spatial arrangement of the atoms in the individual basic building blocks—amino acids and nucleotides, sugars, etc.—are determined by strong directional covalent bonds, the higher-order spatial deployment of the building blocks themselves (and their constituent atoms) in the major types of macromolecules is determined by much weaker chemical forces. These include van der Waals forces as well as the weak bonds, which involve electrostatic interactions between atoms and molecules, interactions which do not involve the sharing of electrons. The weak bonds are ten to twenty times weaker than strong or covalent bonds. Where the energy of a C-C covalent bond is about 350 kJ /mol1, the weak bonds have energies of 4–40 kJ/mol.25
Because of their role in determining the 3-D shape of macromolecules, the weak bonds are, as Watson describes them in The Molecular Biology of the Gene, “indispensable to cellular existence.”26 Their “indispensability” was only realized with the molecular biological revolution. Linus Pauling suspected that the weak bonds would prove pivotal, and his suspicions were confirmed by the discovery of the importance of macromolecular shape. As Crick explained in a speech honoring Pauling, chemists before World War II “were much more concerned with the strong bonds,” and it was some time before they became interested in weak bonds. The weak bonds, as it turns out, “are the ones that fit molecules together in a physical sense, in one way or another, with different degrees, of course, of specificity. Pauling believed that that was going to be the key to everything, the way that things fitted together, using these weak forces.”27
It is the weak bonds that fit together two different parts of an individual large macromolecule (like a protein), or two different molecules (as in the case of the two strands of the double helix), conferring unique spatial architectures on the resulting macromolecular complexes.28 Watson nicely captured the nature and biological function of the weak bonds when he described them as conferring “selective stickiness”29 to bio-matter, a stickiness that determines the way complex molecular structures fit together.
The double helical structure of DNA illustrates the way the strong directional and weak bonds work together to determine the overall atomic architecture of complex macromolecules. In the helix it is the strong directional bonds which determine the spatial position of the atoms in each of the nucleotides in each strand, while it is the job of the weak bonds to hold the two stands of DNA together in the classic, higher-order, 3-D double helical conformation. Consequently, the spatial positioning of the atoms in the double helix is determined by these two very different types of bonds working together.
THE DOUBLE helix also illustrates a major and vital characteristic of the weak bonds—that they can be broken relatively easily and are readily reversible. This characteristic enables the cell to pull apart the two strands of the helix during DNA replication and transcription, and enables them to easily reattach afterwards. If the bonds linking the two strands together were strong covalent bonds, the two strands would be irreversibly bound together; replication and transcription would be practically impossible.
Most biological functions not only require stereospecific arrangement of atoms in macromolecules but also (as with DNA) reversible weak binding between various molecular surfaces. And it is the relative weakness of the weak bonds which makes this possible.
Selective bonding between two molecular surfaces can only be achieved by using a number of bonds which collectively form a unique, complementary lock-and-key pattern of electrostatic interactions linking the two surfaces together. While it would be possible to bind two complementary molecular shapes or surfaces together with several strong covalent bonds, it would be difficult to pull the two molecules apart once the strong bonds had been made. That is, it would be difficult to remove the key from the lock. And even if the energetic barrier could be overcome, there would be the additional steric problem of fitting some “bond breaking” molecular device between the lock and the key to break the individual covalent bonds.
If the bonding between the two complementary molecular surfaces is to be both selective (comprising a number of bonds arranged in a unique pattern) and reversible (essential to most cellular functions), then the individual bonds must not be too strong or their collective action would bind the two surfaces into a rigid, immobile structure. Hence selective, reversible bonding could not be achieved using strong covalent bonds. The only way to achieve both highly specific and weak, reversible lock-and-key bonding between two molecules is to use the combined action of several much weaker bonds.
In short, the relative weakness of the weak bonds (compared with the strong bonds) is exactly what is needed for rapid, highly selective association and disassociation of two complementary molecular surfaces—between the two strands of DNA, between two stretches of the polypeptide chain of a protein, between an enzyme and its substrate, etc.
If weak bonds had been, say, ten times stronger, close to the energy of covalent bonds, selective stickiness would remain possible, but it would be irreversible.30 Biochemistry as it occurs in cells would be impossible. Proteins and all the constituents of the cell would be frozen into rigid, immobile structures, including the two strands of the double helix.31
As Watson noted in Molecular Biology of the Gene, the strength of the weak bonds is “not so large that rigid lattice arrangements develop within a cell—the interior of a cell never crystallizes as it would if the energy of… [weak bonds] were several times greater.” This fact “explains why enzymes can function so quickly, sometimes as often as 106 times per second. If enzymes were bound to their substrates by more powerful bonds, they would act much more slowly.”32
On the other hand, if the strength of weak bonds were lower, then it would be impossible to fit enough of them on complementary surfaces to form a strong enough bond to withstand the hurly-burly and continual thermal jiggling of the constituents within the cell. Reversible, specific binding of a substrate to its catalytic site could never occur, nor, for example, could the head of a molecular motor detach from and reattach so readily to an actin fiber. Rob Phillips points out that as things are, disruptive forces in the cell caused by the random collisions between particles are close to the forces exerted by the weak bonds, meaning the weak bonds couldn’t get much weaker and still do their job.33
In short, for reversibly sticking molecules together in the cell in highly specific stereospecific complexes—the vital basis of virtually all biochemical functions—the average energy level of the weak bonds has to be very close to what it is.
|
Figure 3.3. Energy levels in the cosmos vary over a vast range of magnitudes, from low-frequency photons (~2 × 10–30 J) to the Big Bang (~4 × 1069 J). The energy of one hydrogen bond is about 4–40 kJ/mol = 6.7–67 × 10–21 J, and the average energy of the strong bonds is about ten times greater (one order of magnitude more). |
Also, the absolute strength of the weak bonds and their strength relative to that of the strong bonds must be very close to what they are.
Some idea of the exacting nature of this example of fine tuning can be appreciated by plotting energy levels in the cosmos on a logarithmic scale from the energy of low-frequency radio photons to the energy of the explosion which gave rise to the expanding universe, the “big bang”—a range exceeding one hundred orders of magnitude. In doing so, we find that on our graph34 we can only represent the energy range of the strong and weak bonds as two infinitesimally narrow lines.
Materialism Undermined
OVER THE past two centuries there were two major discoveries which undermined previous vitalistic notions. The first was the discovery that the vital properties of organic compounds resided not in vital agency but in the unique chemical fitness of the carbon atom—along with oxygen, nitrogen, and hydrogen—to generate a vast inventory of compounds of diverse chemical and physical properties. The second was the discovery that the seemingly mysterious vital abilities of cells to carry out enzymic catalysis and to replicate genetic information largely depended on the fitness of two types of chemical bonds which uniquely enable the precise deployment of atoms into extraordinarily complex highly specific 3-D conformations.
For many people, the retreat of vitalism in biology in the two centuries following Wöhler has been a grand victory for positivistic thinking and materialism. Watson waxes lyrical in this vein in his book DNA: The Secret of Life. The discovery of the double helix was so important because it “brought the Enlightenment’s revolution in materialistic thinking into the cell,” he writes. “The intellectual journey that had begun with Copernicus displacing humans from the center of the universe and continued with Darwin’s insistence that humans are merely modified monkeys had finally focused in on the very essence of life. And there was nothing special about it. The double helix is an elegant structure, but its message is downright prosaic: life is simply a matter of chemistry.”35
Philosopher Daniel Dennett echoed these sentiments when he crowed, “Vitalism—the insistence that there is some big, mysterious extra ingredient in all living things—turns out to have been not a deep insight but a failure of imagination.”36
But Watson, Dennett, and their fellow materialists should be more circumspect. When they celebrate each retreat of vitalism as a great victory for materialism and mechanism, they overlook a crucial point: each retreat revealed some additional element of intelligent fine tuning in nature—fine tuning for life.
Yes, vitalism retreated, but in its place the wonder of the unique capabilities of the carbon atom and its peerless fitness for biochemistry was revealed. Yes, vitalism retreated with the discoveries which followed in the wake of the molecular biological revolution, but these only highlighted a greater wonder in the fitness of nature for the assembly of complex macromolecules. These were not wonders beyond scientific analysis, but they were wonders nonetheless, and they pointed not to a homunculus in the cell, but to a far greater wonder-worker who finely tuned the very fabric of nature for life on Earth.