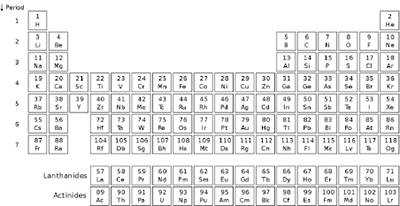
Figure 4.1. The Periodic Table of the Elements.
4. CARBON’S COLLABORATORS
Just as the letters of the alphabet have the potential for endless surprise and enchantment, so, too, do the elements of the kingdom. Unlike an alphabet which has hardly any infrastructure, the kingdom has sufficient structure to make it an intellectually satisfying aggregation of entities. And because these entities are finely balanced, living personalities, with quirks of character and not always evident dispositions, the kingdom will always be a land of infinite delight.
—PETER ATKINS, THE PERIODIC KINGDOM1
THE PREVIOUS TWO CHAPTERS HAVE DESCRIBED MANY ELEMENTS of fitness of the carbon atom for life. But the recipe for life requires more than the carbon atom. Carbon needs collaborators to build the great plenitude of compounds to assemble a complex living system. And, as usual, nature has obliged.
Although most of the atoms in the periodic table are metals and do not form strong covalent directional bonds, carbon’s nonmetal near neighbors in the table, oxygen (O) and nitrogen (N), clustered together near the top right-hand corner, and hydrogen (H), at the upper left, share carbon’s ability to form strong directional covalent bonds (see Figure 4.1). In one of the classic papers arguing for the fitness of nature for carbon-based life, George Wald (awarded a Nobel prize for elucidating the molecular basis of photodetection) emphasized their special fitness for biochemistry:
The special distinction of hydrogen, oxygen, nitrogen, and carbon is that they are the four smallest elements in the Periodic System that achieve stable electronic configurations by gaining, respectively, 1, 2, 3, and 4 electrons. Gaining electrons, in the form of sharing them with other atoms, is the means of making chemical bonds, and so of making molecules. The special point of smallness is that these smallest elements make the tightest bonds and so the most stable molecules; and that carbon, nitrogen, and oxygen are the only elements that regularly form double and triple bonds. Both properties are critically important.2
Not only do hydrogen, oxygen, and nitrogen share with carbon the prime characteristics of fitness for building stable molecules of defined 3-D shape, but their chemical and physical properties are also markedly different from carbon. This diversity is crucial because it allows these elements to introduce into the organic realm carboxyl (COOH), amino (NH2), methyl (CH3), and other groups with novel chemical properties.
If carbon’s nonmetal neighbors in the second row (period 2) of the periodic table, nitrogen and oxygen, had possessed physical and chemical properties similar to carbon (as with most adjacent atoms in most regions of the periodic table), the organic realm would have been much less chemically diverse. Carbon-based life may even have been impossible, or at least restricted to simple unicellular life. But nitrogen, carbon’s near neighbor to the right, and oxygen, the neighbor of nitrogen a step further to the right, are about as different from carbon as can be imagined, being transparent, colorless gases in ambient conditions—very different from a pile of soot or a lump of coal, two common forms of carbon in nature.(Hydrogen, carbon’s other partner, is also a colorless gas in ambient conditions.)
|
Figure 4.1. The Periodic Table of the Elements. |
Also important for life is how different carbon and its period 2 partners are from neighbors outside period 2. Lawrence Henderson stressed this point, noting that these period 2 elements “possess very definite individual properties, which mark them off sharply from other substances.” It’s thus “in the highest degree probable that compounds made from elements of such positive chemical characteristics and very unusual properties will be unlike compounds formed from other elementary substances.” Henderson concludes that this leads “us to believe that other elements are exceedingly unlikely to readily form compounds comparable in number, variety, and complexity with those of organic chemistry as we know it.”3 He further elaborates: “It follows from the peculiarities just explained that the first great factor in the complexity of living organisms as we know them, the complexity and variety of their chemical constituents, depends principally upon the nature of the elements which compose such substances, and is most probably a unique, certainly a very rare characteristic of matter.”4
In a similar vein, Peter Atkins emphasizes that all the neighboring nonmetals atoms in period 2—boron, carbon, nitrogen, oxygen, and fluorine—differ markedly from their homologues immediately below in period 3. Nitrogen, he notes, “is a colorless nonreactive gas,” while its nearest neighbor in the third period “is phosphorus, a colorful reactive solid, while sulfur, directly below the colorless gas oxygen, “is a yellow solid.”5 Indeed, as he further notes, all the atoms of period 2, including boron, carbon, and fluorine are strikingly different from their immediate southern neighbors.6
Only among carbon’s near neighbors in the small, nonmetal region of the periodic table are there atoms able to make strong directional covalent bonds and that possess very different properties, capable of making the right stuff for life when combined with carbon. In this region, the chosen region, as Atkins puts it, “Complexity can effloresce from subtly different consanguinity.”7
In sum, carbon’s genius is to a large degree gifted by its covalent, nonmetal partners, nitrogen, oxygen, and hydrogen, and their differing properties. In fact, it turns out that—consistent with the special fitness of nature for life on Earth—the properties of carbon’s collaborators are precisely what are needed for the chemistry of the cell.8
THE HIGHER an atom’s electronegativity, the greater its attraction for electrons. Of all the similarities and differences among the chemistry of hydrogen, carbon, nitrogen, and oxygen, one of the most consequential involves their electronegativities.
Consider first the carbon-hydrogen bond9 and the chemical consequences of the fact that these two atoms have similar electronegativities.
As a consequence, when hydrogen atoms bond to carbon (C-H), they form what are called non-polar covalent bonds. In these bonds, the electrons are equally distributed between the two atoms. And because of this, there is little or no charge disequilibrium in the resultant bond. In other words, the bonds are electrically symmetrical. There are no electropositive regions (with a deficit of electrons) and no electronegative regions (with a surplus of electrons) around the bond. In C-H bonds, the constituent atoms tug more or less equally on their electrons, the charge distribution remains uniform, and the resulting compounds are non-polar thanks to sharing their electrons equally. As we will see below, one of the effects of this is to render hydrocarbon chains insoluble and hydrophobic, a characteristic which plays a vital role in the life of the cell.
And now we come to something very beautiful, and of the profoundest consequence to all life on Earth. Unlike the bonds that hydrogen makes with carbon, which are non-polar,10 the bonds that hydrogen (H) makes with nitrogen (N) and oxygen (O) are nearly always polar, involving unequal sharing of electrons. This is because, unlike hydrogen and carbon—which are relatively close on the Pauling electronegativity scale (0.35 units apart)—the electronegativity of hydrogen differs markedly from that of nitrogen or oxygen (0.7 and 1.3 units apart respectively). Because of the unequal attraction of the electrons, the oxygen and nitrogen atoms in O-H and N-H bonds are negatively charged while the hydrogens are positively charged.
One of the most important of polar molecules is water (H-O-H), the matrix of life. The polar nature of water molecules is responsible for its great power as a solvent in dissolving charged ions or polar compounds. When water molecules come in contact with a charged ion or polar compound, the water molecules experience electrostatic interactions (charge-based attractions). The negatively charged oxygen atoms are attracted to positively charged ions such as Na+ or regions of electrostatic positivity in a polar compound, while the positively charged hydrogen atoms are attracted to negatively charged ions such as chloride Cl– or regions of electronegativity in polar compounds. We call such ions and polar molecules hydrophilic (“water-loving”).11
As there are nearly always many water molecules relative to solute molecules, these interactions lead to a sphere of water molecules around the solute. These hydration shells allow particles to be dispersed evenly in water and keep the charged solutes apart and unable to combine and precipitate out of the solution and, therefore, soluble. As the majority of organic molecules—sugars, alcohols, fatty acids, amino acids, nucleotide bases—contain charged groups or are polar, they lend themselves to the formation of hydration shells and are readily soluble in water. And as water is the matrix of the cell, the solubility of so many compounds is of great utility.
But what happens when non-polar, long-chain hydrocarbons try to join the electrostatic party? Because of the absence of charged regions, water molecules cannot form hydration shells around the molecules, and the hydrocarbon chains are excluded from the party and forced into insoluble, water-avoiding clusters. For this reason, long-chain hydrocarbons are termed hydrophobic (“water-fearing”). And the force which clumps them together into water-excluded clusters is termed the hydrophobic force.
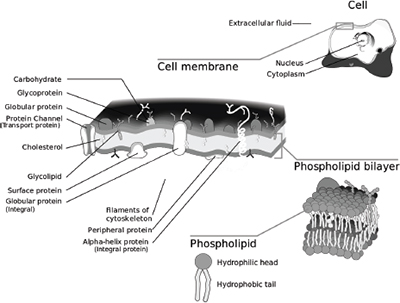
But rather than being a defect in the order of things, the insolubility and hydrophobic character of hydrocarbons is a vital element of nature’s fitness for life. The insoluble hydrocarbons form the core of the lipid bilayer membrane that surrounds the cell and many of the cell’s internal organelles—one of the most important supramolecular structures in the cell, indeed in the entire biological world. It also allows newly synthesized proteins to fold into their secondary structures, one of the most important of all the biochemical processes in the cell.
The basic building blocks of the cell membrane consist of long, insoluble, hydrophobic hydrocarbon chains linked to a polar hydrophilic group or head containing O-H and N-H bonds. Because they contain a hydrophobic group linked to a hydrophilic group, they are termed amphiphilic, from the root words amphi (meaning “both”) and phileo (meaning “to love”). These building blocks are known as phospholipids, because they contain a phosphate group in the hydrophilic head. Because of their hydrophobic character, the hydrocarbon chains or tails are forced to cluster together into two layers in the center of the membrane (away from water), with the water-loving hydrophilic groups clustered at the interface between the hydrocarbon bilayer and the aqueous medium on the inside and outside of the cell.
As John Philip Trinkaus comments in his wonderful book Cells into Organs, “Because water is itself a strongly polar molecule, the polar phosphate of the membrane lipids will inevitably be attracted to the surfaces of the membrane, both external and cytoplasmic. And just as inevitably their nonpolar fatty acid parts will tend to be squeezed into a nonpolar phase in the interior of the membrane.”12 It is this dual character of the phospholipids that enable them to form membranes, without which cells would not exist.
The insoluble hydrophobic hydrocarbon’s other important role in the cell is in protein folding. Many amino acids possess a hydrophobic, insoluble, hydrocarbon side chain (leucine and isoleucine are two examples). However, all amino acids (including those with hydrophobic side chains) also contain the two polar groups COOH and NH2. This means that the hydrophobic amino acids, like the phospholipids in the membrane, are also amphiphilic and soluble to a degree. The same hydrophobic force that causes the hydrophobic tails of the phospholipids to cluster together in the center of the membrane forces the hydrophobic non-polar side chains to cluster into a central core, playing a decisive role in protein folding.
|
Figure 4.2. The lipid bilayer cell membrane. |
In an additional teleological twist, these water-avoiding aggregates that are clustered together in the center of the folded protein provide the cell with tiny, non-aqueous micro-environments vital to the life of the cell. Many of the synthetic and enzymatic processes on which the cell depends can only occur in a chemical micro-environment free of water. So, the relative electronegativities of C, H, and O are responsible for the hydrophobic force, which plays a vital role in the folding and function of proteins.
Charles Tanford was not exaggerating when he said, “The hydrophobic force is the energetically dominant force for containment, adhesion, etc., in all life processes,” adding: “This means that the entire nature of life as we know it is a slave to the hydrogen-bonded structure of liquid water.”13
A Finely Tuned Quartet
THE WAY the different electronegativities of hydrogen, carbon, oxygen, and nitrogen work together towards the formation of the cell membrane and the folding of proteins is amazing. On the one hand, the electrical asymmetry of oxygen-hydrogen bonds leads to the hydrophilic character of water and is the source of the hydrophobic force, which clumps the insoluble non-polar hydrocarbons into the bilayer membranes and clumps the hydrophobic amino acid side chains into the center of proteins. On the other hand, the electrical symmetry of carbon-hydrogen bonds makes the clumping possible by conferring on long hydrocarbon chains their non-polar, hydrophobic, water-avoiding behavior.
This is so elegant. In the face of the wondrous way nature works such a miracle, one can only cite John Keats’s Ode On a Grecian Urn: “Beauty is truth, truth beauty.”14
At the heart of cellular life is an extraordinary reciprocal fitness between the non-polar carbon-hydrogen (C-H) bonds and the polar oxygen-hydrogen (O-H) bonds. This reciprocity gifts life with the cell membrane and the folding of proteins. If the electronegativity of hydrogen, carbon, oxygen, and nitrogen had been the same, unquestionably there would be no carbon-based life on Earth. The cosmos has only come to life because of the different electronegativities of the four collaborators.
And as we have seen, in another teleological twist, it is hydrogen’s electronegativity that is the cause of water’s polar nature (via the electrical asymmetry of the O-H bond) and of the non-polar nature of the long hydrocarbon chains (because of the electrical symmetry of the C-H bond). Hydrogen’s place in Linus Pauling’s electronegativity scale is responsible for both the hydrophobic character of long hydrocarbon chains and for the character of water that renders them insoluble.
Carbon-based life likely would be impossible if the electronegativities of hydrogen (H), carbon (C), oxygen (O), and nitrogen (N) were even slightly different from what they are. So, for example, imagine a world where the electronegativities of these four elements were closer to one another, a world devoid of polar molecules. Alternately, envision a world where C-H bonds were polar and O-H and O-N bonds non-polar. Neither of these imagined worlds would contain carbon-based life, even if all the other properties of these four elements were exactly the same. And not because we lack the imagination to see how life could manage in these counterfactual worlds. Just the opposite. We can assert the impossibility precisely because molecular biologists have done the painstaking work of uncovering the wonder manifest in this unique band of atoms.
The unique capacity of carbon to bond with itself, its capacity to form multiple bonds, the metastability of so many carbon compounds, the directionality and strength of the covalent bonds of carbon and its nonmetal compatriots, the existence of weak chemical forces such as van der Waals forces and weak ionic bonds of appropriate strength for lock-and-key bonding—all these would be useless without the fine-tuning of the relative electronegativities of oxygen, nitrogen, and hydrogen. Only when the whole suite of fitness is complete can the miracle of the cell be actualized.
And as for water’s inability to solubilize oils and fats and other hydrocarbons, this might seem a defect in the so-called universal solvent. But as we have seen, the hydrophobicity of hydrocarbons, and their insolubility in water, is one of the prime elements of nature’s fitness for carbon-based life. An apparent shortcoming of water turns out to provide a previously unsuspected but vital element of nature’s fitness for carbon-based life.
The Fitness of the Cell Membrane: A Checklist
THE MEMBRANE’S many particular properties that make it so fit to form the bounding membrane of the cell are worth looking at individually.
Semipermeability: Perhaps the most important function of the membrane is providing the cell with a semipermeable bounding membrane to separate it from the external environment.
No cell could survive without some sort of membrane that was relatively impermeable to the cell’s constituents, especially small metabolites such as sugars and amino acids but also larger molecules such as proteins and RNAs. The cell needs a way to prevent these from diffusing away into the surrounding fluid. It must have been this way from the beginning. The very first cells must have been enclosed by a membrane to retain the mix of biochemicals within.
The membrane is non-polar, which makes it particularly impervious to polar and charged compounds. This is vital because the small organic molecules in the cell—a mix of sugars, nucleotide bases, metabolic acids, etc.—are nearly all polar molecules. So the lipid bilayer is fit to provide not only a barrier between the inside and outside of the cell, but one that is non-polar—precisely the sort needed to retain the small charged metabolites inside the cell.
At the same time, the membrane isn’t perfectly impermeable. That would make cell survival equally impossible, preventing it from getting nutrients and expelling waste. This problem is solved by the existence in the membrane of special gates or pores through which myriads of small compounds and ions pass in and out of the cell in a highly controlled manner (see below).
Self-Organizing: A second emergent property of the membrane is its self-organizing ability. This remarkable property allows the membrane to form automatically around the outer surface of the cell. As Trinkaus comments in Cells into Organs, “Because of their intrinsic chemical nature… phospholipids naturally and spontaneously self-assemble… to form a bilayer in a watery solution, sequestering their hydrophobic ends in the interior while their polar hydrophobic ends are ‘in solution’ on the surface. It is as it were ‘the nature of the beast’ for them to do so.”15
In addition, any bounding membrane must possess the ability to self-repair so it can readily close any hole or break that would allow the cell’s contents to diffuse away. The membrane needs to be able to adapt to the minute-to-minute deformations in cell form and repair itself to maintain a continuous barrier at all times in the face of all sorts of contingencies. It’s not hard to imagine the huge problems a cell would face if its bounding membrane were not self-mending and did not quickly close any hole or defect in its surface.
The lipid bilayer wonderfully satisfies the necessity for both self-assembly and self-repair. As Trinkaus explains, it can flow in every direction over the cytoplasmic surface so as to maintain a continuous barrier between cell and surroundings, and it manages this in the face “of the ever-changing protrusive activities of the cell surface.”16
Trinkaus describes the membrane as a “two-dimensional liquid”17 and comments, “It is of great significance for cell motile behavior that the lipid bilayer of the plasma membrane has an overall high fluidity (low viscosity) at the normal body temperature of homeotherms.”18
It is therefore not incidental that every cell on Earth has a lipid bilayer forming the skin or boundary layer. Its qualities of impermeability (except at the gates), relatively high fluidity, and spontaneous assembly and self-repair appear to be essential to a cell’s existence. This unique combination of characteristics is only found in the lipid bilayer, another case where a crucial biological function, the bounding of the cell, is carried out by a structure that appears to be both unique and ideal for its assigned role. No other known material could substitute for this particular structure.
The cell would encounter profound challenges if it used a membrane that was not self-organizing or had to be assembled piece by piece. Not only would assembly be a colossal logistical and energy-demanding challenge; the cell also would need a complex sensing system to inform it of the minute-to-minute changes in membrane conformation. Clearly, a mechanical skin that could not self-organize would never work as a barrier surrounding the ever-shifting shape of a living cell.
While crawling, a cell continually changes shape, especially at the leading edge. This is known as the lamellipodium, which is full of actin fibers that grow to propel the cell forward. During crawling, many cells create micro-protrusions or filipods—narrow cylindrical extensions containing actin fibers—which they use like a cat’s whiskers to detect environmental clues, initiate contact with surrounding cells, and convey signals back to the cell body.1920
Filipods are used, for example, during axon growth and guidance by path-finder neurons to find their way in the developing nervous system.21 Plant, fungal, and most bacterial cells, encased in rigid cell walls, have less need for the self-sealing, self-organizing properties of the cell membrane. But in cases where cells undergo continual changes in form, as when an amoeboid white cell chases down a bacterium in the bloodstream or during cell movements in embryogenesis, the self-sealing, self-organizing ability of the membrane is essential.
Without the innate self-organizing property of lipid bilayer membranes, there would be no crawling, no embryogenesis, and probably no thinking beings to contemplate the loss.
Membrane Diversity: The self-organizing nature of the membrane also facilitates the generation of a great variety of emergent structures, including tubes, vesicles, and various types of layered structures.22
The different morphologies of the membranes in the chloroplast and those constituting photoreceptive discs in the outer segments of the photoreceptors in the vertebrate eye testify to the adaptive versatility of this remarkable structure. The same basic membrane structure makes up the endoplasmic reticulum, encloses the nucleus and the mitochondria, and so forth. As I wrote in Biology and Philosophy:
All this plethora of forms arises spontaneously by self-organization from the physical properties of various bilayer lipid membranes of differing chemical composition. Different lipids and proteins can bend and distort the basic membrane form into various globular forms, vesicular forms or tubules. The various forms arise again like the protein folds entirely from the local interactions between membrane constituents. A zoo of vesicular forms can be generated spontaneously from lipid membranes depending on the ratio of lipid and protein constituents. This is the focus of a lot of current work. As Huttner and Schmidt comment: “The shape of biological membranes reflects the shape of its principle constituents—that is membrane lipids and integral membrane proteins, as well as their interaction with each other and with peripherally associated proteins (including the glycoproteins and proteoglyans), the cytoskeleton and the extracellular matrix.”23
And in reporting some fascinating work on a membrane-altering protein dynamin, Wieland Huttner and Anne Schmidt remark, “Dynamin alone is sufficient to change the shape of liposomes, causing either tubulation or vesiculation, depending on lipid composition.”24 Thus, by altering the protein and lipid constituents of membranes, the cell can generate vesicles from planar surfaces, tubules from planar surfaces, and vesicles from tubules. The intrinsic properties of cell membranes generate a vast zoo of emergent membrane structures.
The Right Width: Another crucial property of the lipid bilayer is how remarkably thin it is, only five nanometers.25 If a typical mammalian cell were expanded to the size of a pumpkin, its lipid bilayer membrane would be thinner than a sheet of paper. But despite its thinness, it is highly robust and can maintain its oily integrity in the face of the varied micromechanical forces buffeting it.
The remarkable thinness of the membrane isn’t just a striking curiosity. It’s essential to the cell’s fitness. Proteins average about five nanometers across, about the same as the width of the membrane. And this means that individual proteins can be designed to stretch across the thickness of the membrane. This is important because there are several membrane functions which require individual proteins which can span the width of the membrane. Some make up the ion channels that selectively control the passage of ions—mainly Na+ and K+—across the membrane. Others maintain different ion concentrations on the two sides of the membrane. They also couple the transport of ions with that of small organic molecules,26 relaying signals between a cell’s interior and exterior. Other important transmembrane proteins are those involved in cell-to-cell recognition and selective adhesion.27
Several intriguing elements of fitness determine the width of the membrane. For example, the membrane requires pliable, relatively stable hydrocarbon chains. The chains also need to be sparingly soluble. The chain length of the hydrocarbons used in membranes in the cell is between sixteen and eighteen carbon atoms long. Hydrocarbon chains of more than eighteen carbon atoms long are at ambient temperatures too stiff, insoluble, and wax-like. They cannot be readily mobilized in water and are not sufficiently fluid or pliable for the membrane. On the other hand, chains less than sixteen carbon atoms long are too mobile and unstable. But chains of sixteen to eighteen are just right.28 As Arthur Needham points out, one factor restricts chain-length to some degree, while additional factors further restrict the functional range:
The chains are held in orientation by the London-van der Waals attractions of neighbouring molecules and this is effective only within a certain range of chain-lengths, namely C16 to C36 or so; this is precisely the range of the common biological fatty acids... Beyond this length the molecules topple and tangle, while shorter chains have insufficient attraction… If the lipid is also polar, like the fatty acids, that is if it has a hydrophil group in the molecule, then members shorter than C16 also tend to pass into aqueous solution too readily to form stable membranes.… The C18 fatty acids are much the most abundant in plants and animals… C–even members up to C38 have been discovered in biological material but they only serve to emphasise the unique exploitation of the C16–18 node. Anything shorter does not build stable crystalline layers and anything longer is less soluble in water, protects proteins less well, has too high a melting point, and so on.29
How fortunate it is that these various essential factors overlap in the sixteen-to-eighteen range of chain length. The two C18 hydrocarbon chain layers result in the five-nanometer width of the membrane, just the right width for individual proteins to stretch across.
Membrane Potential: Because the membrane—or more specifically the central hydrocarbon layer—is an electrical insulator, an electric charge can be built up across the membrane. The difference in electric charge across the cell membrane is known as the membrane potential. Cells generate the potential by regulating the movement of charged ions across the membrane through highly selective ion channels.
Resting mammalian cells may use 30 percent of their metabolic energy in generating the membrane potential, which may rise up to 70 percent30 in nerve cells. This may seem like a prodigal use of cellular energy, but the charge serves many important functions, as Crichton explains:
Many transmembrane transporter proteins termed secondary transporters, use the discharge of an ionic gradient to power the “uphill” translocation of a solute molecule across membranes. Coupling solute movement to ion transport enables these secondary transporters to concentrate solutes by a factor of 106 with a solute flux 105 faster than by simple diffusion… Sugars and amino acids can be transported into cells by Na+-dependent symports. Dietary glucose is concentrated in the epithelial cells of the small intestine by a Na+-dependent symport, and is then transported out of the cells into the circulation by a passive glucose uniport situated on the capillary side of the cell.31
The cell’s capacity to generate and maintain a charge across its membrane allows for the transmission of nerve impulses along the axons of neurons in animal nervous systems. The transmission of a nerve impulse depends on what is called the action potential. This is an extremely rapid depolarization of the membrane, which occurs via a sudden influx of millions of Na+ ions into the cell in a fraction of a millisecond.32 The impulse or action potential travels along the axon at speeds up to one hundred meters a second. As Bruce Alberts and his co-authors explain:
In nerve and skeletal muscle cells… Na+ channels… open, allowing a small amount of Na+ to enter the cell down its electrochemical gradient. The influx of positive charge depolarizes the membrane further, thereby opening more Na+ channels, which admit more Na+ ions, causing still further depolarization. This process continues in a self-amplifying fashion until, within a fraction of a millisecond, the electrical potential in the local region of membrane has shifted from its resting value of about –70 mV to almost as far as the Na+ equilibrium potential of about +50 mV.33
The membrane’s electrical insulating character and the resulting membrane potential provides precisely the electrical characteristics required for the transmission of electrical impulses between cells and ultimately for the construction of the nervous system of beings like ourselves. This suggests, as in many other instances, that nature’s fitness is not just for the carbon-based cell but also for advanced multicellular organisms like ourselves.
The generation of membrane potential and the nerve impulse depend critically on the mobility of small ions, which can move rapidly down concentration and electrical gradients. Their mobility and speed moving through ion channels is astounding. Alberts and his colleagues explain that “up to one million ions can pass through one open ion channel each second.”34 Without these tiny, highly mobile inorganic ions, no cell would be able to regulate or generate a membrane potential or a nerve impulse. No other small particles of matter possess charge and such great mobility. Neither proteins nor any of the organic molecules in the cell have the right properties to stand in for the alkali metal ions.
For the first time in this book, we have described a cellular function that involves atoms other than the four partner atoms carbon, hydrogen, oxygen, and nitrogen. Membrane potential and nerve transmission depend on the existence of billions of charged particles in the cell and the surrounding extracellular fluids, and on the very high diffusion rates of sodium and potassium ions.
The Quartet in Review
WE HAVE seen in the previous two chapters that the atoms carbon, hydrogen, oxygen, and nitrogen are impressively fit for the assembly of the vast plenitude of organic compounds with their diverse properties upon which the life of the cell depends. Among other things we saw that because of their directional strong covalent bonds, they form compounds with specific molecular shapes and that the existence of compounds with defined molecular shapes together with the weak chemical bonds enables the formation of large macromolecules of defined 3-D shape, macromolecules that can perform specific biochemical and cellular functions.
And as we have seen in this chapter, these four atoms possess just the right electronegativities to make polar molecules via the O-H and N-H bonds. They also provide non-polar bonds via C-H bonds, which render long-chain hydrocarbons hydrophobic and insoluble in polar fluids such as water, and this leads in turn to the emergence of the self-organizing wonder of the bilayer lipid cell membrane, which as we have seen has a whole suite of equally extraordinary properties, all of which are indispensable to the cell.
In short, these four atoms are mutually fit, in a myriad of improbable ways, to set in motion a succession of unique emergent phenomena, along a pathway stretching from the relative electronegativities of carbon, hydrogen, oxygen, and nitrogen to the nervous system of higher organisms. The way these elements work together to generate this unique path is stunning, conveying an irresistible impression of contrivance.