Figure 8.1. Cosmic abundance of elements.
8. THE PRIMAL BLUEPRINT
An honest man, armed with all the knowledge available to us now, could only state that in some sense, the origin of life appears at the moment to be almost a miracle, so many are the conditions which would have to be satisfied to get it going.
—FRANCIS CRICK, LIFE ITSELF (1981)1
THERE IS SOMETHING EVOCATIVE AND ENDLESSLY FASCINATING about the idea of a message sent to Earth from space by an advanced extra-terrestrial civilization, revealing information about our own origin, existence, and place in nature. It is the stuff of many popular works, from Carl Sagan’s Contact and Stanley Kubrick’s 2001: A Space Odyssey to Erich von Däniken’s book Chariots of the Gods.
Although not the work of aliens, a very special chemical message regarding the place of carbon-based life in the universe did arrive on Earth on the night of September 28, 1969.2 On that night, the sky over the small southeastern Australian town of Murchison was lit up by an exploding meteorite, which scattered fragments of rock over the nearby countryside. Subsequent chemical analyses of the meteorite’s fragments proved, for the first time, that at least some of the organic building blocks of life are being constantly synthesized in space and exist in vast quantities throughout the cosmos.3 Moreover, very recent analyses suggest the total number of different carbon compounds embedded in the meteorite may number in the tens of thousands and perhaps even millions.4
The Murchison meteorite revealed that the cosmos is seeded with a vast inventory of organic chemicals, including amino acids5 and nucleobases,6 the starting points in the assembly of the two main polymers—proteins and nucleic acids—in carbon-based organisms. Just how many of the basic organic building blocks of life might have been produced abiotically in space and brought to Earth in meteorites (or synthesized in the primeval ocean) is not clear. But more and more are being identified in meteorites and synthesized in the lab in simulated prebiotic conditions.7
More recent spectroscopic analyses of interstellar gas have revealed that the cosmos is seeded not just with some of the basic monomers of life, but also with far more complex carbon compounds. One class of complex carbon compound—the polycyclic aromatic hydrocarbons (PAHs)—is abundant throughout the cosmos, and some may contain up to one hundred carbon atoms.8 Some contain nitrogen (PANHs), forming heterocyclic compounds that have a chemical structure similar to the heterocyclic compounds used in living things, such as the nucleotide bases.9
It is not hard to see a parallel between Friedrich Wöhler’s landmark synthesis of urea in 1828 and the message the Murchison meteorite delivered. Wöhler undercut the need for a vital force in the cell for manufacturing the basic organic constituents of life, but their synthesis still required a cell or a chemist. The Murchison meteorite and subsequent study of other meteorites and spectroscopic analysis of interstellar space imply that the universe is replete with organics and that when the inventory is more fully known, it very well may contain many more of the basic building blocks of life. These have been synthesized, to paraphrase Wöhler, not only without a kidney, but even without a cell or a chemist. And these discoveries have, to a large extent, validated a widespread belief of origin-of-life researchers in the twentieth century, since Stanley Miller’s ground-breaking experiment in 1953, that the basic ingredients of life can be synthesized abiotically in nature.
The Miller-Urey experiment involved sending a spark through an atmosphere thought to mimic the atmosphere of the primeval Earth (water vapor, methane, ammonia, and hydrogen). The result was a complex chemical mix that contained glycine, alanine, and aspartic acid, three of the amino acids used in building proteins in modern organisms.10 Recent studies of his archived material from his original experiments subjected to more advanced analytical techniques show that ten of the twenty biologically important amino acids—lysine, alanine, serine, threonine, aspartic acid, valine, glutamic acid, methionine, isoleucine, and leucine—were present in Miller’s flasks.11
Of course the three amino acids reported by Miller, and even the ten in the later accounting, are a long way from the inventory of monomers needed to assemble a living cell. Much less does it explain how the inventory of monomers were assembled to form the first cell. Nonetheless, Miller-type experiments and evidence from the analysis of meteorites such as the Murchison meteorite do show that at least some of the key building blocks of life can be, and indeed are, synthesized abiotically, and may be common throughout interstellar space. This is no small discovery. For they show that at least the first step towards the cell requires no more than “ordinary chemistry” and suggests that the subsequent steps to the cell might also be explicable in terms of the known laws of chemistry and physics—that nature might be sufficient.
Thus we see that the message brought to Earth that fateful September night, written in the chemistry of a falling star, is highly significant. As well as supporting the claim that life’s emergence might have been the result of entirely natural mechanisms, it also supports the thesis central to this book and the whole Privileged Species series: Carbon-based life as it exists on Earth is no contingent afterthought of nature, no artifactual accident, but an inherent part of the natural order. It is an inherent part of nature’s grand design from the moment of creation.
Cosmic Abundance
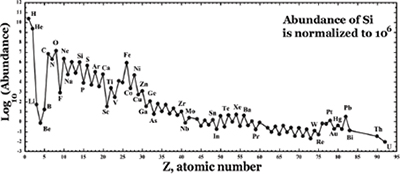
THERE IS further evidence that life may be no contingent cosmic afterthought but an end programmed into the order of things from the beginning. The atoms carbon, oxygen, and nitrogen were among the first atoms synthesized in the stars. These joined hydrogen, already in existence, to make up the universe of organic chemicals, the substances upon which Earth’s whole carbon-based biosphere is built. (When Lawrence Henderson wrote his classic Fitness, the source of carbon, oxygen, nitrogen, and various other elements heavier than hydrogen and helium was a mystery, since the nuclear synthesis of the atoms of the periodic table in the interior of stars was only elucidated by Fred Hoyle and others in the mid-twentieth century.) These four atoms are also, along with helium, commonest of all atoms in the universe.12
|
Figure 8.1. Cosmic abundance of elements. |
Even a cursory observation of the cosmic abundance of the elements reveals an obvious correspondence between the cosmic pattern generated in the heart of the stars and life on Earth, between the cosmic order and the order of life, between man and cosmos. The elements hydrogen (H), carbon (C), oxygen (O), and nitrogen (N), the core atoms that combine to form the molecules of organic chemistry that compose 96 percent of the human body, are respectively the first, third, fourth, and fifth most abundant elements in the cosmos. And their order of abundance curiously corresponds to their abundance in the human body.13 Two of the three most abundant elements, hydrogen and oxygen, make up water (H2O), the matrix of life, which forms more than 60 percent of the mass of the human body. And another key molecule, carbon dioxide (CO2)—the ideal carrier of the carbon atom to all life on Earth—is formed from the third and fourth most abundant atoms: oxygen and carbon.
Other prominent constituents of life are also among the most abundant of elements: magnesium, sodium, calcium, iron, phosphorus, potassium, and sulfur. The overall picture conveys the powerful impression that stellar nuclear synthesis—the atom-building process in the stars—was set up from the beginning to serve the end (the purpose) of life on Earth.
It is important to stress that the selection of the atoms which enable the biochemistry of life is not because of their cosmic abundance, but because they possess the right chemical and physical properties to serve a vast number of highly specific physiological and biochemical functions in the cell.14 Carbon, for instance, possesses the right properties to build a vast diversity of chemical compounds, and this would be true even if it were not the fourth most abundant element in the universe. No other atom possesses the same fitness. In other words, the laws which determine the cosmic abundance of carbon and the other atoms of life are quite distinct from the laws that determine their fitness for life. So here is a genuine coincidence indicative of a deep biocentricity in the cosmic order: the great majority of the most abundant atoms are the most fit for life.
The Elusive Path
DESPITE THE message of the Murchison meteorite—that the cosmos is seeded with the atoms of life and even with many of the core organic molecules, including amino acids and nucleobases—just how or where the transition from soup to cell occurred is an abiding mystery, among the greatest unsolved problems in science.
We do know that banded stromatolite formations generated by mats of bacterial cells very similar to modern blue-green algae first appear in the fossil record about 3.5 billion years ago,15 and there is some fossil and isotopic evidence which implies life might have originated as early as 3.7 billion years ago.16 So we know life has graced our planet for billions of years. But we know virtually nothing about how it originated.
The depth of the mystery is compounded by the fact that we do know, as the previous chapters in this book show, a great deal about the chemical basis of life. We have known since the early nineteenth century that the building blocks of the cell are perfectly natural chemical forms, determined by natural law. After the Murchison discovery and recent astrophysical studies, we know that at least some of these building blocks occur in vast quantities throughout the cosmos. We also know, in astounding detail, the atomic structures and molecular behavior of the key macromolecular constituents of the cell—such as DNA and RNA, proteins, and lipid membranes. We also have known since the 1960s the basic design of the cell, the meaning of the genetic code, and how information flows from the DNA into proteins. More recently, we have uncovered undreamt-of depths of complexity in the genome, including a mushrooming inventory of tiny regulatory RNAs.17
The size of standard textbooks like Molecular Biology of the Cell18 is a testimony to the vast amount of knowledge about life acquired since the 1953 discovery of the double helix (the same year Miller published the results of his famous experiment). Relevant fields outside of biology also have seen major advances, in supra-molecular chemistry for example, which have greatly expanded our knowledge of the behavior of soft matter in the mesoscopic domain.
Yet despite our extensive knowledge of the molecular biology of the cell, we remain at a complete loss as to what may have been the basic steps which led from the Murchison monomers to the cell system in terms of the known laws of chemistry and physics.
Rather than reveal the elusive path from a chemical soup to the last common ancestor of all extant life, the spectacular progress in cell biology and organic chemistry outlined above has revealed just how immense is the chasm between a soup of organic compounds and the cell with its membrane, the necessary complement of enzyme catalysts, the proteins’ synthetic apparatus, genetic information encoded in the double helix, and so forth. Despite many heroic attempts,19 no one has produced any convincing explanation of how nature could have overcome this chasm, the vastness of which was described recently by Stephen Meyer in his Signature in the Cell.20 (See also Chapter 11 in my own Evolution: A Theory in Crisis21 and Brian Miller’s chapter in the newly revised and expanded edition of The Mystery of Life’s Origin.22)
The origin-of-life community has identified various steps to the cell. It is widely accepted that four of these are the formation of basic building blocks such as the amino acids and nucleotides; their polymerization into proteins and DNA; the formation of the first primitive replicating system; and the evolution of the modern DNA and protein cell system with a functioning genetic code and an apparatus for protein synthesis. Only work on the first step has seen substantial progress. How the other steps were accomplished in terms of known laws of nature is a complete enigma. The widely acknowledged reality is that within the entire corpus of twenty-first century science, there is no explanation. Science, it seems, has reached an impasse. The origin of life remains as arguably the biggest unsolved problem in science.
In a critical paper summarizing this impasse, specifically with regard to the problem of how the genetic code and translation system could have emerged, Eugene Koonin and Artem Novozhilov offer the following comment:
At the heart of this problem is a dreary vicious circle: what would be the selective force behind the evolution of the extremely complex translation system before there were functional proteins? And, of course, there could be no proteins without a sufficiently effective translation system. A variety of hypotheses have been proposed in attempts to break the circle but so far none of these seems to be sufficiently coherent or enjoys sufficient support to claim the status of a real theory.23
About proto-protein synthesizing systems that were halfway to the modern cell, they comment, “These and other theoretical approaches lack the ability to take the reconstruction of the evolutionary past beyond the complexity threshold that is required to yield functional proteins, and we must admit that concrete ways to cross that horizon are not currently known.”24
About the transition from an RNA world to the modern DNA/protein world and its associated models, they write that “we are unaware of any experiments that would have the potential to actually reconstruct the origin of coding, not even at the stage of serious planning.”25
Summarizing the state of the art they confess to “considerable skepticism.” As they explain, “It seems that the two-pronged fundamental question: ‘why is the genetic code the way it is and how did it come to be?,’ that was asked over 50 years ago, at the dawn of molecular biology, might remain pertinent even in another 50 years. Our consolation is that we cannot think of a more fundamental problem in biology.”26
Overcoming the Impasse
SO HOW did the transition from soup to cell occur? One obvious explanation is the idea that an intelligent agency assembled the first cell. This is an explanation popular among some supporters of intelligent design. Although it is rejected by most academic biologists, as the evidence stands it is perhaps as convincing an explanation as any available. However, an alternative possibility (my own preferred position) is that there are new laws, or novel properties of matter yet to be discovered, which enabled the path from chemistry to the cell.
For example, one problem that would have to be overcome in any naturalistic framework is what’s known as the clutter problem. In all known prebiotic syntheses, in addition to the desired monomers there is a vast universe of other small, reactive organics. These include a great inventory of closely related amino acids and nucleotides, differing slightly from their biologically important relatives, who are just as likely to join up with the desired monomers, making a polymeric chaos.27 Getting linear bio-polymers made up of a few basic monomers in a prebiotic setting and avoiding multiple unwanted side reactions with various reactive organics is a real poser. How this could have been achieved prebiotically, before enzymes existed, is a huge problem.
As Gerald Joyce comments, “The chief obstacle to understanding the origin of RNA-based life is identifying a plausible mechanism for overcoming the clutter wrought by prebiotic chemistry.”28 Each of RNA’s four components “would have been accompanied by several closely related analogues… which could have assembled in almost any combination.”29 Joyce explains:
The nucleotides (and their analogues) may even have joined to form polymers, with a combinatorial mixture of 2’,5’-, 3’,5’- and 5’,5’-phosphodiester linkages, a variable number of phosphates between the sugars, D- and L- stereoisomers of the sugars, α- and β-anomers at the glycosidic bond, and assorted modifications of the sugars, phosphates and bases. It is difficult to visualize a mechanism for self-replication that either would be impartial to these compositional differences or would treat them as sequence information in a broader sense and maintain them as heritable features.30
In discussing how the clutter problem might be overcome, Joyce continues: “Perhaps there were special conditions that led to the preferential synthesis of activated β-D-nucleotides or the preferential incorporation of these monomers into polymers.” He then elaborates:
For example, the prebiotic synthesis of sugars from formaldehyde can be biased by starting with glycoaldehyde phosphate, leading to ribose 2,4–diphosphate as the predominant pentose sugar… The polymerisation of adenylate, activated as 5’-phosphorimidazolide, yields 2’5’-linked products in solution, but mostly 3’5’-linked products in the presence of a montmorillonite clay. Thus, through a series of biased syntheses, fractionations and other enrichment processes, there may have been a special route to a warm little pond of RNA.31
If the origin of life did occur naturally, in some as yet unidentified way, there must be an underlying special fitness in nature to overcome the clutter problem.
Robert Shapiro also anticipated the discovery of new mechanisms and principles. “Self-replicating systems capable of Darwinian evolution appear too complex to have arisen suddenly from a prebiotic soup,” he wrote. “This conclusion applies to both nucleic acid systems and to hypothetical protein-based genetic systems. Another evolutionary principle is therefore needed to take us across the gap from mixtures of simple natural chemicals to the first effective replicator. This principle has not been described in detail or demonstrated, but it is anticipated, and given names such as chemical evolution and the self-organization of matter.”32
Nobel Prize winner Jack Szostak (Medicine, 2009) also expresses the idea that there must be novel phenomena yet to be discovered to explain how life originated. As he and Itay Budin comment, “The discovery of novel physical mechanisms will be essential for a better understanding of how life could have begun.”33
Paul Davies sounds a similar note in his book The Fifth Miracle, though he lays particular emphasis on how radical he suspects the solution will be. “Real progress with the mystery of biogenesis will be made, I believe, not through exotic chemistry, but from something conceptually new,”34 he comments. He even speculates that perhaps the weird behavior of matter at a subatomic level might have played a role: “Here is a mainstream physical theory that has information at its heart, which it tangles with matter in an intimate way.… Could some sort of quantum-organizing process be just what is needed to explain the origin of informational macromolecules?”35
He goes further, connecting all this to the idea of purpose:
Deterministic thinking, even in the weaker forms of de Duve and [Stuart] Kauffman, represents a fundamental challenge to the existing scientific paradigm.... Although biological determinists strongly deny that there is any actual design, or preordained goal, involved in their proposals, the idea that the laws of nature may be slanted towards life, if not contradicting the letter of Darwinism, certainly offends its spirit. It slips an element of teleology back into nature, a century and a half after Darwin banished it.36
A paper by Tommaso Bellini and his colleagues seems to hint at something similar when they contrast the Darwinian scenarios, which they term “fantastic luck theories,” with fine-tuning (fitness) theories. “Although ‘fantastic luck’ scenarios are not forbidden by natural laws, they appear increasingly unlikely and hence ‘unacceptable’ to the sensitivity of the scientists,” they write. “As we have seen, the direction taken by the [origin of life] research is to propose scenarios where the ‘fantastic luck’ is reduced, and replaced by a stronger degree of necessity. How far this could go, how much our existence can instead be viewed as necessary, woven in the deep structure of Nature, is a question that has always interested scientists.”37
However, at the moment, none of even the most speculative scenarios provides anything beyond the most tentative explanation to avoid the impasse. But this does not mean there will never be a naturalistic explanation. In 1890 no physicist could have conceived of the implications of twentieth-century quantum physics. Which physicist in 1890 could have envisaged wave/particle duality or superposition or the connection of two particles separated by cosmic distances? No one could have imagined the radical implication of the new physics. Given the radical nature of the twentieth-century revolution in physics, the possibility of new discoveries which are currently inconceivable, but which may throw light on the origin-of-life problem and overcome the impasse, cannot be discounted.
IRRESPECTIVE OF what proximate causes led to the origin of the cell, and irrespective of what may have been the enigmatic steps from the monomers of Murchison to the last common ancestor of all extant life on Earth, the creation of the first carbon-based cell here or, indeed, anywhere in the universe was only possible because of the ensemble of fitness in nature described in this monograph—what I have termed the primal blueprint. As we have seen, this ensemble is in effect a detailed blueprint, integral to the emergence of the carbon-based cell, written into the laws of nature from the beginning, long before it was instantiated in material form.
As we have seen, the blueprint includes:
1. The fitness of the carbon atom to form stable covalent bonds with itself and hydrogen, oxygen, and nitrogen, generating the vast inventory of organic compounds (Chapter 2).
2. The fitness of the directional property of covalent bonds that enable the assembly of large macromolecules with defined 3-D shapes capable of specific biological functions—enzymatic, structural, and genetic (Chapter 3).
3. The fitness of weak bonds to form complementary electrostatic surfaces that can reversibly stick together different parts of macromolecules (such as the two strands of DNA), enzymes to their substrates, and molecular motors to actin fibers (Chapter 3).
4. The fitness of the differences in the electronegativities of carbon, hydrogen, and nitrogen, which leads to the non-polar hydrophobic hydrocarbons and confers on water its polar hydrophilic character. The particular differences in electronegativity among these atoms give us insoluble long-chain hydrocarbons, which form the basis of the cell membrane and enable stable folded proteins (Chapter 4).
5. The fitness of the emergent properties of the cell membrane, including semi-permeability, self-organizing ability, and insulating capacity, properties fit to serve many indispensable biological ends. These include selective adhesion and crawling, and electrical properties that enable the nervous systems of higher organisms (Chapter 4).
6. The unique abilities of phosphates to store and use energy in the aqueous medium of the cell (Chapter 5).
7. The fitness of various metal atoms, including iron and copper, for channeling electrons down electron transport chains and for oxygen handling in hemoglobin and cytochrome c oxidase (Chapters 5 and 6).
8. The fitness of the sodium (Na+) and potassium (K+) ions for rapid charge transport across the membrane and the maintenance of the membrane potential (Chapter 6).
9. The fitness of water to serve as the fluid matrix of the cell, including its low viscosity to serve as the medium of circulation, its unsurpassed fitness as a solvent, its hydrophobic force, and its fitness for proton conduction (Chapter 7).
Moreover, the blueprint is not restricted to the generic cell. There are various aspects of the blueprint that appear specifically fit for the cells of higher organisms. For example, aerobic cells of higher organisms depend on the use of biological oxidations to satisfy their energy-hungry metabolism, and this in turn is only possible because of the unique unreactivity of oxygen at ambient temperatures, and the properties of the transition metal atoms such as iron and copper for activating and handling oxygen. The transport of oxygen to the tissues is enabled by the iron atom in hemoglobin. And the transport of CO2 from the tissues to the lungs depends on the unique properties of the zinc atom in the enzyme carbonic anhydrase.
The cells of complex multicellular organisms also depend on water’s low viscosity and the high diffusion rates of solutes in water, which enable the circulatory system as well as metabolically active cells large enough to possess the cytoskeletal machinery to crawl and morph, skills essential in embryogenesis. The high rates of diffusion of solutes in water is also key to the fitness of the small, rapidly diffusing sodium, potassium and chloride ions (Na+, K+, and Cl–) for generating the membrane potential, an indispensable adaptation for transmitting nerve impulses in higher organisms.
Mutual Fitness
WHILE THE properties of the individual atoms are impressively fit for specific biological functions, even more impressive is, as Henderson stressed in The Fitness of the Environment, their mutual fitness to work together to achieve various vital ends. In discussing the way water and carbon dioxide work together, Henderson described their association as so close “that it is scarcely correct logically to separate them at all; together they make up the real environment and they never part company.”38 And again he points out that the fitness of carbon dioxide is “dependent upon water… resting upon solubility and ionization; upon interactions between the two substances.”39 Again he stresses that it is “by many independent and united actions”40 that the ensembles achieve their life-promoting collective properties.
In these pages we have reviewed various remarkable ensembles of reciprocal fitness, many of them discovered in the decades following Henderson’s seminal work. There is the reciprocal fitness of the insoluble non-polar hydrocarbons and the polar hydrophilic nature of water, which gives us the bilayer lipid membrane and the folding of proteins. There is the mutual fitness of the oxygen atom to accept one electron at a time and the ability of transition metals to donate individual electrons, enabling the controlled reduction and activation of oxygen. And there is the mutual fitness of the strong and weak bonds which work together in the formation of the specific shapes and functions of large macromolecules. Those are just three of countless such instances of reciprocal fitness.
Parsimony
FURTHER INTRIGUING evidence of nature’s special fitness for the carbon-based cell is the remarkable fact that the same four atoms—hydrogen, carbon, oxygen, and nitrogen—which combine to form the basic organic substances of the cell also provide, by simple combinations with each other, three simple molecules which are vital to life: water (H2O), the supreme matrix; carbon dioxide (CO2), the unique carrier of the carbon atom to all parts of the biosphere; and ammonia (NH3), the compound via which the nitrogen atom is introduced into the organic domain.
It is not just that simple combinations of the four main atoms of organic chemistry provide these three vital molecules but that they are, as we saw above, among the most abundant atoms in the cosmos. Water is formed from the most common atom, hydrogen, and the third most common atom, oxygen. Carbon dioxide (CO2) is formed from the fourth and third most common atoms in the cosmos, carbon and oxygen. Ammonia (NH3) is formed from the most common and fifth most common atoms in the universe, hydrogen and nitrogen.
There is yet more. Cells require energy, and where do we find an atom to generate energy by chemical reaction with the substances of life, and with unparalleled efficiency? Again, we need look no further than among these same four atoms. One of them, oxygen, the third most common atom in the cosmos, is uniquely fit to provide copious quantities of energy for living systems. No other atom comes close. Additionally, the oxidation of carbon and hydrogen releases greater amounts of energy than any other type of oxidation.41 So two of the same four atoms—carbon and hydrogen—serve as maximally fit stores of metabolic energy.
Has any artifact of man ever transcended the wondrous elegance and parsimony of the ensemble of fitness embedded in these four atoms? Only someone committed to rejecting out of hand all evidence of teleology in nature could fail to see in these ensembles of fitness and in this elegant parsimony evidence of design.
Alien Life
THE PRIMAL blueprint means that nature is uniquely fit for the cell as it exists on Earth, to the almost certain exclusion of any other hypothetical forms of chemical life, natural or artificial. And this is predictive in an important sense: it suggests that all chemical life throughout the cosmos will be carbon-based and resemble life on Earth; or at the very least it suggests that any life found elsewhere in the cosmos that even approaches the sophistication of life on Earth will be based on the same ensemble of carbon and its primary chemical collaborators.
As Henderson noted more than a century ago, “Carbon, hydrogen, and oxygen, each by itself, and all taken together, possess unique and preeminent chemical fitness for the organic mechanism.”42 The fitness of this ensemble, he adds, “results from characteristics which constitute a series of maxima—unique or nearly unique properties of water, carbonic acid, the compounds of carbon, hydrogen, and oxygen…—so numerous, so varied, so nearly complete among all things which are concerned in the problem that together they form certainly the greatest possible fitness… to promote complexity, durability, and active metabolism in the organic mechanism which we call life.”43
He asks, “What are the possibilities of obtaining the same characteristics in other substances?”44 His answer, “No other environment… could possess a like number of fit characteristics” for promoting “the organic mechanism we call life.”45
Anyone familiar with the current state of astrobiology knows that no other set of atoms has ever been discovered or even imagined that could equal the mutual fitness of the members of the ensemble that underlies the existence of carbon-based life.
Astrobiologists are guided by NASA’s motto, “follow the water,” in their search for life beyond Earth. And this amounts to a tacit admission of the lack of a suitable alternative solvent. Astrobiologists Frances Westall and André Brack call liquid water the “perfect solute for carbon-based life.” And they don’t stop at water. They add: “Schulze-Makuch and Irwin (2004) make an excellent discussion about the possibility of life based on other chemical compounds and solutes but none of these combinations presents the same range of properties and advantages as carbon and water and, at least on Earth, most are not stable or are outcompeted by carbon.”46
In the journal Astrobiology, Christopher McKay lists the following as minimum requirements for a life-detection mission: “(1) Liquid water of suitable salinity, past or present; (2) Carbon in the water; (3) Biologically available N in the water; (4) Biologically useful energy in the water; (5) Organic material that can possibly be of biological origin and a plausible strategy for sampling this material.”47
One century after Henderson, no fact has come to light to seriously threaten his conclusion that the laws of nature are finely tuned for life as it exists on Earth. On the contrary, many discoveries and advances unimagined in 1913 have further confirmed it. These include the importance of the hydrophobic force, the utility of strong covalent and weak bonds for the assembly of complex macromolecules, proton conduction, and the fitness of ten or so metal atoms for very specific cellular functions. Further confirming Henderson’s conclusion are advances outside the biological sciences, such as the elucidation of the cosmic abundances of the atoms, the carbon-12 resonance that enables stars to generate carbon, and the fine-tuning of the constants of physics for life.
Darwinism Diminished
ANOTHER IMPLICATION of the primal blueprint is the obvious challenge it poses to the Darwinian world view. The existence of the blueprint specifying the design of the cell long before life emerged on Earth implies that, however the first cell was assembled, whether suddenly through the direct agency of a Divine watchmaker, or gradually by a blind “Darwinian” watchmaker (constrained by natural law and utilizing the natural selection of chance events), or by some other as yet unimagined means, its creation was only possible because the blueprint was already in place.
The fitness of the atoms and the makeup of the primal blueprint for the cell was determined long before the Earth was born. Consequently, even if we posit a blind Darwinian watchmaker as the major player in the actualization of the blueprint, its actualization was only possible because the blueprint was already in place.
In Sum
WE HAVE seen in this monograph that the miracle of the cell is only possible because of a vast prior ensemble of mutual fitness in the unique properties of a set of about one-fifth of the atoms of the periodic table. There is, I believe, no other corpus of evidence anywhere in science which provides more convincing evidence of design in nature and purpose in the universe than the fine tuning of these atoms for the carbon-based cell. On any consideration, the evidence presented in this monograph conveys the irresistible impression that the properties of the atoms have been contrived directly and purposefully to enable the existence of life in the universe.
As far as the actualization of the blueprint is concerned, which remains the deepest of mysteries, I believe that further elements of fitness in nature will be discovered over the coming decades which will finally reveal the fateful path from chemistry to life. And I believe that when the path is finally elucidated, it will turn out to be extraordinary, one of the greatest of scientific wonders, revealing a far deeper teleology in nature than all the elements of natural fitness for the cell and life documented so far.
Even more, I believe that the elucidation of that fateful route will be of far greater intellectual consequence than any other discovery in science since the birth of science in the sixteenth century. Indeed, I believe the path, when discovered, will prove to be so obviously indicative of a profound teleology in the very ground of being that it will prove a watershed in the history of thought.
Conversely, if instead it is eventually established that there is no purely natural path across the great gulf from non-life to life, and that only the additional exertion of an intelligent agent could have assembled the first cell on Earth, that will be equally a watershed in human thought.
Finally, even if the design inference is rejected, the existence of the primal blueprint for the cell, revealed as a result of scientific discoveries over the past two centuries, provides what is surely irrefutable evidence that the cosmos as currently understood is uniquely fit for carbon-based life, and that life as it exists on Earth and beings of our biological design occupy a very special place in nature. Irrespective of any design inference, what science has revealed already confirms the deep intuition of the medieval Christian scholars who believed that “in the cognition of nature in all her depths, man finds himself.”48