1 Beyond the Passive/Active Dichotomy: A Spectrum Model of the Brain’s Neural Activities
Introduction
General Background
The way in which investigators think about the brain can have a deep impact on the empirical investigations of neuroscience as well as on the interpretation of their philosophical implications. One model of the brain, favored by the early British neurologist Sir Charles Sherrington (1857–1952), proposed that the brain and the spinal cord were primarily reflexive. Reflexive in this model means that the brain reacts in predefined and automatic ways to sensory stimuli such as auditory or visual stimuli. Stimuli from outside the brain, originating externally in either the body or the environment, are assumed to determine completely and exclusively the subsequent neural activity. The resulting stimulus-induced activity, and more generally, any neural activity, in the brain is then traced back to the external stimuli to which the brain merely passively reacts. One may therefore speak of what I describe as the passive model of the brain.
An alternative view, however, had already been suggested by one of Sherrington’s students, Thomas Graham Brown. In contrast to his teacher, Brown suggested that the brain’s neural activity—that is, within the spinal cord and brain stem—is not primarily driven and sustained by external stimuli from outside the brain. Instead, Brown held that the spinal cord and brain stem do show spontaneous activity that originates internally within the brain itself. Hans Berger, who introduced the electroencephalogram (EEG) early on (Berger, 1929), also observed spontaneous activity in the brain that remained independent of any external tasks or stimuli.
Other neuroscientists, including Bishop (1933), Lashley (1951), and Goldstein (2000), followed Brown’s line of thought and proposed that the brain actively generates its own activity internally, that is, it generates spontaneous activity—or resting-state activity as it is also called in its operational-behavioral version (see Northoff, 2014a; Raichle, 2015a,b; as well as discussion below). Recently, the view of central or spontaneous activity has gained more traction in neuroscience with the observation of spontaneous oscillations (Buzsáki, 2006; Llinás, 1988; Yuste, MacLean, Smith, & Lansner, 2005), spontaneous coherence or connectivity between different regions’ neural activities (Biswal et al., 1995; Greicius, Krasnow, Reiss, & Menon, 2003), and the default-mode network (DMN) (Greicius et al., 2003; Raichle, 2015b; Raichle et al., 2001).
These and other observations point out the central role of the brain’s spontaneous activity for its neural activity including both resting state and task-evoked or stimulus-induced activity (see Huang, Zhang, Longtin, et al., 2017; Northoff, 2014a,b; Northoff, Qin, & Nakao, 2010 for an extensive discussion). This leads me to speak of an active model of the brain. Such an active model of the brain is nicely illustrated in the following quote by the early German neurologist Kurt Goldstein in his book The Organism, which appeared originally in 1934 (Goldstein, 2000):
The system is never at rest, but in a continual state of excitation. The nervous system has often been considered as an organ at rest, in which excitation arises only as a response to stimuli. … It was not recognized that events that follow a definite stimulus are only an expression of a change of excitation in the nervous system, that they represent only a special pattern of the excitation process. This assumption of a system at rest was especially favored by the fact that only the external stimuli were considered. Too little attention was given to the fact that the organism is continuously exposed, even in the apparent absence of outward stimuli, to the influence of internal stimuli—influences that may be of highest importance for its activity, for example, the effect of stimuli issuing from the blood, the importance of which was particularly pointed out by Thomas Graham Brown. (Goldstein, 2000, pp. 95–96)
Recently, the issue of how to model the brain has gained increased traction given the discovery of the DMN (Raichle et al., 2001). The DMN is a neural network that covers various regions predominantly in the middle of the brain, the so-called cortical midline structures (Northoff & Bermpohl, 2004; Northoff et al., 2006). The DMN shows particularly high levels of metabolism and neural activity in the absence of any specific external stimuli, which condition has been termed the resting state (Logothetis et al., 2009; Raichle, 2015a,b; Raichle et al., 2001; see also Klein, 2014, for an excellent discussion in a more philosophical context).
Since its initial discovery, the high levels of resting state or spontaneous activity (for the sake of simplicity I use both terms here interchangeably) in the DMN have been associated with different mental features such as self, consciousness, mind wandering, episodic memory retrieval, time prospection and retrospection, and random thoughts (Christoff, 2012; D’Argembeau et al., 2010a, 2010b; Fazelpour & Thompson, 2015; Northoff, 2012a–c, 2014b; Northoff et al., 2006; Smallwood & Schooler, 2015; Spreng, Mar, & Kim, 2009). Because it is apparently implicated in a wide range of different functions, the exact role of the DMN remains unclear at this point in time.
What is clear, however, is that the nature of the DMN’s activity supports an active model of the brain. Put into a more philosophical context, some authors (Churchland, 2012; Fazelpour & Thompson, 2015; Hohwy, 2014; Northoff, 2012a) have compared the active model of the brain to the model of mind developed by Kant (1781/1998). Briefly, Kant argued against a passive model of mind wherein its activity would be completely determined by external stimuli, which was the view of David Hume. Instead, Kant conceived of the mind as exhibiting spontaneity, entailing an active rather than passive model of mind. The old dispute between Hume and Kant about passive versus active models of mind has thus resurfaced in the form of a quandary in theoretical neuroscience.
Main Aim and Argument
The main focus in this chapter is on discussing empirical evidence in favor of different models of the relationship between spontaneous and stimulus-induced activity. This serves to develop an empirically plausible model of brain activity, the spectrum model (see parts I–III). I argue that the disjunction of passive (part I) and active (part II) models of brain is ill conceived. The brain neither generates its neural activity in a completely passive way as driven by the external stimuli nor in an exclusively active way, that is, entirely driven by its spontaneous activity. Instead, based on empirical evidence, we need to accept a model of brain that undermines the passive/active dichotomy and integrates both in a spectrum that allows for categorizing different forms of neural activity according to the degree of the brain’s participation in generating that activity (part III).
Note that this claim is purely neuronal: I argue that different sorts of neural activity involve resting-state activity to different degrees, with some being more active while others remain rather passive. The spectrum model of brain is not directly concerned with mental features and how they are related to the spectrum of neural activities, although such work may eventually become viable once the relevant neuronal features are more deeply understood. Instead, I here focus exclusively on the brain’s neuronal features, specifically on how its neural activity falls on a continuum or spectrum between purely active and passive modes. I leave open the spectrum model’s relation to mental features such as consciousness, which is discussed in chapter 4.
Definition and Clarification of Concepts
Before continuing, we need to shed a brief light on some terminological issues. First, there is the distinction among different forms of neural activity, spontaneous activity, resting-state activity, and stimulus-induced activity. Spontaneous activity refers to the neural activity that is generated within the brain itself independent of any external stimuli from outside the brain, that is, interoceptive stimuli from the body and exteroceptive stimuli from the world (see Northoff, 2014a; Raichle, 2015a,b). The term spontaneous activity thus denotes the origin of neural activity and is meant in a purely neuronal sense.
This is different from the concept of resting state, which describes a particular behavioral condition: eyes closed or open with fixation and the absence of any specific stimuli or tasks (see Northoff, 2014a; Raichle, 2015a,b). The resting state is often taken as the operational condition that measures the brain’s spontaneous activity. For the sake of simplicity I here use the terms “resting state” and “spontaneous activity” in an interchangeable way to denote the absence of any stimuli from outside the brain (including both interoceptive stimuli from the body and exteroceptive stimuli from the world).
In addition, we also need to distinguish the term stimulus-induced or task-evoked activity. Operationally both resting state/spontaneous activity and stimulus-induced/task-evoked activity can well be distinguished: the resting state is measured with eyes closed or open in the absence of specific stimuli or tasks, whereas stimulus-induced activity is tested for by applying specific stimuli or tasks.
However, the distinction between those two forms of neural activity may no longer be as clear cut when taken in a more physiological sense because it may be that external stimuli simply modulate the ongoing spontaneous activity. This would make the distinction between the forms of activity rather relative (in at least operational terms) if not superfluous (in physiological terms) (see Klein, 2014; Northoff, 2014b; Raichle, 2009, 2015a,b; as well as below for details). The main aim of this volume is to discuss different possible relations between resting-state/spontaneous activity and stimulus-induced/task-evoked activity and to assess their viability according to available empirical data.
Finally, we also need to clarify the concepts of active and passive. In this context the concepts of active and passive concern the degree to which the brain itself provides a contribution to its own neural activity. The passive end of the spectrum would apply to neural activity that is determined by external stimuli, whereas the active end of the spectrum applies when the brain itself exhibits neural activity prior to and independent of external stimuli.
Part I: Passive Model of the Brain
The passive model describes the brain’s neural activity as being dependent on external stimuli it receives from body and environment. The resulting neural activity, stimulus-induced activity, is taken to be determined entirely by the external stimulus; the brain itself just passively processes the external stimuli and does not actively participate in constituting its own neural activity. A passive model of brain can come in two extreme versions, strong and weak, and an intermediate version, the moderate version. I here sketch briefly the two extreme versions.
The strong passive model argues that the brain has no impact at all on how it responds to stimuli. The weak passive model of brain could acknowledge spontaneous activity in the brain but assign it just a modulatory but not a causal role. A causal impact of spontaneous activity on stimulus-induced activity would involve the former causing the latter, so that that there would be no stimulus-induced activity without spontaneous activity even in the presence of external stimuli. In the case of a modulatory impact, stimulus-induced activity would still be present even in the absence of spontaneous activity, which, if present, serves just to modulate the degree of stimulus-induced activity. Based on recent empirical evidence, I argue in the chapters that follow that both versions, weak and strong, as well as the moderate version of the passive model of brain are to be rejected.
Passive Model of Brain Ia: Strong Model—Absence of Resting-State Activity
The strong passive model claims that stimulus-induced activity in the brain can be accounted for entirely by external stimuli. Moreover, a strong passive model of the brain would claim that there is no neural activity at all in the brain unless there is stimulus-induced activity. However, empirical evidence conflicts with these claims. Accordingly, the strong passive model is not empirically tenable and thus merely logically conceivable; a discussion of why it is not tenable, however, is helpful for understanding the other more tenable views.
How can we characterize the brain’s resting state? One should be aware that the concept of the brain’s intrinsic or resting-state activity or spontaneous activity (I use all three terms interchangeably) is a rather heterogeneous one and raises several questions (see also Cabral, Kringelbach, & Deco, 2013; Mantini, Corbetta, Romani, Orban, & Vanduffel, 2013; Morcom & Fletcher, 2007a,b; Northoff, 2014a).
Terminologically, different concepts are used to describe the resting state. In addition to resting-state activity, other terms including baseline, spontaneous activity, or intrinsic activity are also used to describe the internally generated activity in the brain (see Deco, Jirsa, & McIntosh, 2013; Mantini et al., 2013; Northoff, 2014a). Importantly, the brain’s resting-state activity is not restricted or limited to a particular region or network in the brain (Northoff, 2014a; Raichle, 2009). Instead, it is pervasive throughout the whole brain.
The brain is an energy-hungry system. It consumes 20 percent of the whole body’s overall glucose and oxygen budget while accounting for only 2 percent of the body’s weight (Shulman, Hyder, & Rothman, 2014). Most importantly, all that energy is mainly invested into the spontaneous activity itself; neural activity in response to external stimuli, that is, stimulus-induced activity, only uses a tiny fraction of that, amounting to 5 percent of the overall energy budget of the brain (Raichle, 2015a,b). What does the brain’s resting state do with all that energy? The high amounts of glucose and oxygen seem to be mainly invested in neuronal signaling and activity, with 75–80 percent accounting for the latter (Rothman, De Feyter, Graaf, Mason, & Behar, 2011).
These data suggest close coupling between metabolism and neuronal activity, which then is only slightly modified during stimulus-induced activity (considered from an energetic perspective). Metabolic activity can be measured by the cerebral metabolism rates of glucose or oxygen (CMRglc or CMRO2), whereas neuronal activity can be accounted for by measuring the cycles between glutamate and glutamine (as between neurons and glia with the former being converted into the latter by the enzyme glutamine synthetase) as well as between glutamine and GABA (with the latter being synthesized out of the former via glutamate by the enzyme GAD67) (see Hyder et al., 2006; Shulman et al., 2014).
Investigations in both animals (rats) and humans indicate close coupling between metabolic and neuronal activity: the higher the metabolism in the brain, the higher its neural activity, as in spontaneous or resting-state activity, whereas the metabolism is only marginally increased, if at all, during stimulus-induced activity (Hyder, Fulbright, Shulman, & Rothman, 2013; Hyder et al., 2006; Hyder, Rothman, & Bennett, 2013; Shulman et al., 2014).
Taken together, the strong passive model would need to argue that there is no spontaneous activity internally generated by the brain independent of any external stimuli from outside the brain. This is not compatible at all with the empirical data as reflected in the brain’s metabolism and its close coupling to neural activity. Importantly, the high metabolism and its transformation into neural activity are not related to any external stimuli from outside the brain. These factors speak strongly against the strong passive model of brain.
Moreover, a strong passive model of brain would need to presuppose a principally different design of the brain without any metabolism and neural activity independent of external stimuli. Both metabolism and its coupling to resting-state activity would consequently remain absent. This makes it clear that the strong passive model is merely conceivable (on logical grounds) but not a tenable paradigm (on empirical grounds). However, despite being empirically nontenable, the strong passive model can nevertheless teach us about the central relevance of the brain’s metabolism and its coupling to neural activity for the brain’s operation and functioning (even if the empirical details of such neuro-metabolic coupling remain yet to be explored).
Passive Model of Brain Ib: Moderate Model—No Impact of Resting-State Activity on Stimulus-Induced Activity
The advocate of the passive model of brain may now want to argue that the presence of the brain’s resting-state activity even in sensory cortices can well be acknowledged without undermining the basic assumption of the passive processing of stimuli. Specifically, even if resting-state activity is present in sensory cortices, the stimulus-induced activity in these regions can nevertheless be sufficiently and exclusively related to the external stimuli themselves. Resting-state and stimulus-induced activity would then operate in a parallel and segregated way with no interaction (whether causal or modulatory) between them. In that case the resting-state activity in the sensory cortices should have no impact at all on stimulus-induced activity in the same regions entailing that the former is not necessary at all for the latter. That amounts to a moderate version of the passive model of brain, which again is not supported by empirical evidence.
One functional magnetic resonance imaging (fMRI) study focused on the auditory cortex (Sadaghiani, Hesselmann, & Kleinschmidt, 2009). These researchers let subjects perform an auditory detection task and presented broadband noise stimuli in unpredictable intervals of 20–40 ms. The subjects had to press a button when, and only when, they thought they heard the target sound; otherwise, they did not hit the button. This allowed the researchers to compare the neural activity preceding hits with the activity preceding instances where subjects did not hear the target sound.
Interestingly, successful detection was preceded by significantly higher prestimulus activity, for example, resting-state activity, in auditory cortex, when compared to missed detection. Thus, the level of resting-state activity in auditory cortex impacted the degree of perception, such as whether subjects could hear the auditory stimuli.
But what about rest–stimulus interaction in a sensory modality other than the auditory? The same group also investigated rest–stimulus interaction in the visual modality (Hesselmann, Kell, Eger, & Kleinschmidt, 2008). Higher pre-stimulus resting-state activity levels in the fusiform face area were related to subsequent perception of a face rather than a vase in the Rubin ambiguous vase–face figure. Therefore, higher resting-state activity in the fusiform face biases the subsequent perceptual content toward seeing the face, rather than the vase.
Analogous findings were observed with another visual stimulus such as visual motion: the resting-state activity in the visual motion area in the middle temporal cortex (V5/MT) predicted the degree of the subsequent perception of coherent motion (Hesselmann et al., 2008). Hesselmann and colleagues also related pre-stimulus resting-state activity and peak stimulus-induced activity with behavioral performance: the less prestimulus resting-state activity and peak stimulus-induced activity correlated each other, the better the subjects’ subsequent behavioral performance, for example, the motion perception. Hence, better behavioral performance went along with increased distinction of stimulus-induced activity from the preceding resting-state activity.
Does rest–stimulus interaction also hold in regions other than the sensory cortex? Coste, Sadaghiani, Friston, and Kleinschmidt (2011) conducted a Stroop task wherein the names of colors interfered with the color in which the respective color names were presented (the word “green” was, for instance, presented in the color “red”). Subjects had to push a button to determine the color and whether it was congruent or incongruent.
This study again showed that the prestimulus activity in relevant regions such as the anterior cingulate cortex (ACC) and the dorsolateral prefrontal cortex (DLPFC) predicted subsequent behavioral performance, that is, reaction times. The higher the prestimulus resting-state activity in the ACC and the DLPFC, the faster the subsequent reaction times in response to the stimuli.
Whereas this concerns cognitive regions like the ACC and the DLPLFC, the reverse relation was observed in sensory regions involved in color and word processing: the higher the prestimulus resting-state activity in the right color-sensitive area and the visual word-form area, the slower the subsequent reaction times. These data clearly show that rest–stimulus interaction is mediated by both higher-order cognitive and lower-order sensory regions but in different ways: higher prestimulus activity leads to faster reaction times in cognitive regions, whereas it induces slower reaction times in sensory regions. Since it is one and the same stimulus that both sensory and cognitive regions process, the differential impact of high prestimulus activity levels on reaction times can only be due to differential impacts of the regions themselves, for example, cognitive and sensory, on the external stimulus. This entails different forms of rest–stimulus interaction.
What do these findings imply for our argument against the strong passive model? They demonstrate that stimulus-induced activity in sensory cortices and other regions such as the prefrontal cortex is not sufficiently and exclusively related to the external stimulus itself (as either sensory or cognitive stimulus). Instead, the prestimulus resting-state activity level seems to impact the degree or amplitude of stimulus-induced activity. This means that the external stimulus itself is only necessary, but it is not sufficient by itself for determining stimulus-induced activity.
Accordingly, the empirical evidence from these and other studies (see Northoff, 2014a; Northoff, Qin, & Nakao, 2010, for review) speaks against such exclusive determination of stimulus-induced activity by the external stimulus and thus against the moderate passive model of brain. One should be aware that the stimulus being necessary but not sufficient for stimulus-induced activity would still allow for resting activity to have a merely modulatory impact on stimulus-induced activity. That leads us to the weak passive model of brain as it shall be discussed below.
Passive Model of Brain IIa: Weak Model—No Causal Impact of Resting-State Activity on Stimulus-Induced Activity
The proponent of the passive model of brain may nevertheless not yet be ready to relinquish his or her view of the brain as passive. He may strengthen his argument by weakening his claim, stating that the resting-state activity may indeed modulate stimulus-induced activity, whereas it does not causally impact such activity in the same way that the external stimulus does. Only the external stimulus has a causal impact on stimulus-induced activity— meaning that without external stimuli there would be no stimulus-induced activity.
In that case the resting state has at best mere modulatory impact. Modulatory impact means that the resting state does not cause stimulus-induced activity, which therefore would still persist even in the absence of the resting state; the resting state can only modulate or vary the degree of stimulus-induced activity, whereas the latter’s occurrence remains independent of the former. Accordingly, the resting state is not a necessary condition of stimulus-induced activity when it exerts only modulatory impact. This amounts to a weak passive model of brain, which, like its more extreme siblings, conflicts with the empirical data. The data show that the resting-state activity’s impact is not only modulatory but causal.
How can we experimentally demonstrate that resting-state activity causally impacts the stimulus-induced activity? One strategy here may be to vary the overall global level of resting-state activity and then to see how that impacts stimulus-induced activity during particular tasks. This was done in animals in a study by the group led by Robert Shulman. He tested how the baseline or resting-state metabolism impacts subsequent stimulus-induced activity in an animal study (Maandag et al., 2007).
Maandag et al. (2007) induced pharmacologically high and low levels of resting-state activity in rats and measured their neural activity in fMRI during forepaw stimulation. The high level of resting activity condition was associated with widespread activity across the cortex and rather weak evoked activity in sensorimotor cortex during the forepaw movement. This pattern was reversed in the low resting-state activity condition wherein neural activity was stronger in the sensorimotor cortex but more or less absent in other cortical regions. These results demonstrate that the level of resting-state activity causally impacts stimulus-induced activity (see also G. Shulman et al., 2009; R. Shulman, Hyder, & Rothman, 2009; van Eijsden, Hyder, Rothman, & Shulman, 2009, for a discussion of the results by Maandag et al. on a conceptual level).
Passive Model of Brain IIb: Weak Model—Causal Impact of Resting-State Activity on Stimulus-Induced Activity
The empirical data show the causal impact of the resting state on stimulus-induced activity in animals. Is there empirical evidence for analogous causal relationship between resting state and stimulus-induced activity in humans? For that, Qin et al. (2013) devised a clever experimental design by taking advantage of the distinctions between different baselines. They delivered the same auditory stimuli once during eyes open and once during eyes closed. This allowed them to test for the causal impact of two different resting states, that is, eyes open and closed, on the stimulus-induced activity related to the same stimulus.
First, based on a special acquisition technique in fMRI called sparse sampling, Qin et al. determined the impact of the scanner noise on the auditory cortex and compared that condition to the complete absence of any scanner noise. Sparse sampling is an experimental scenario in which the scanner noise is shut down for some seconds while the neural activity, that is, the blood oxygenation level dependent signal (BOLD) response during the period can be recorded after the scanner is turned on again (because the BOLD response shows temporal delay). As expected, this yielded strong activity changes in the bilateral auditory cortex in the comparison of noise versus no noise. These activity changes during the comparison of noise versus non-noise served to determine and locate the auditory cortex’s resting-state activity, albeit indirectly, via the comparison of noise versus no noise. Qin and colleagues then used the auditory cortex as the region of interest for the subsequent analyses.
In a second step, Qin et al. (2013) conducted data acquisition in fMRI during eyes open and closed to investigate the resting-state activity in visual cortex and its modulation by a very basic stimulus, eyes open. Analogous to the noise in auditory cortex, the eyes open condition served to determine the visual cortex’s resting-state activity; this region was then used as a region of interest in subsequent analyses. Data in both eyes-open and -closed conditions were acquired in two different modes, in 20-s periods (block design), which allowed for the generation of BOLD changes, that is, neural activity, and in 6-min periods to determine functional connectivity of the visual cortex to other regions, for instance, the auditory cortex.
Finally, in a third step, Qin (Qin et al., 2012) investigated auditory name perception in two conditions, eyes open and closed, by letting subjects listen to the same names during both conditions: with closed and open eyes. This strategy served to investigate the impact of eyes open and closed, mirroring different baselines (i.e., eyes closed and open), on stimulus-induced activity associated with the same stimulus.
How did the different resting states, that is, eyes closed and open, modulate stimulus-induced activity? During eyes closed, the subject’s own name induced significantly stronger activity in auditory cortex than other persons’ names. Such response difference between one’s own and other names disappeared, however, when the names were presented during eyes open. Because the stimuli were the same in both cases, the absence of any difference in signal change between one’s own and other names during eyes open is most likely to be due to the difference between eyes open and close in the spontaneous activity itself (although an impact from the body and its interoceptive stimuli cannot be completely excluded).
Therefore, the spontaneous activity in the auditory cortex must have undergone some changes when the eyes were opened, thereby apparently changing its sensitivity, especially to the stimuli of the others’ names. Although we currently do not know what exactly changed in the resting state itself during the transition from eyes closed to eyes open, our data nevertheless demonstrate the causal impact of the resting-state activity level on subsequent stimulus-induced activity in auditory cortex.
Our data indicate that there must be some causal interaction between the resting-state activity and the stimulus-induced activity in auditory cortex. Hence, the amount or degree of stimulus-induced activity is determined not only by the stimulus itself but also by the level of the resting-state activity. However, the exact neuronal mechanisms underlying the resting state’s causal impact on stimulus-induced activity remain unclear at this point in time (see also He, 2013; Huang, Ferri, Longtin, Dumont, & Northoff, 2016; Northoff, 2014b; Huang, Zhang, Longtin, et al., 2017).
Different modes of rest-stimulus interaction are plausible. For instance, the stimulus-induced activity may just be added on top of the ongoing resting state activity. Alternatively, the stimulus may elicit a degree of stimulus-induced activity that is either weaker or stronger than the mere addition between the resting-state activity level and the stimulus-induced activity in which case one would speak of nonadditive interaction. Which model holds, the additive or nonadditive model? This determination is subject to ongoing investigation (see chapter 2 for details).
Passive Model of Brain IIc: Causal versus Modulatory Impact of Resting-State Activity on Stimulus-Induced Activity
Taken together, these findings suggest that the resting state has an active causal impact on stimulus-induced activity. The resting-state activity causally interacts with the stimulus, especially a subject’s own name, and becomes therefore a necessary (although nonsufficient) condition of the resulting stimulus-induced activity.
However, a proponent of a weak passive model may be tempted to argue that this example only shows the modulatory, rather than causal, impact of the resting state on stimulus-induced activity. In that case, one would expect the resting state to impact, that is, to modulate all three names in the same unspecific way; but the data do not show this because there are strong differences in how the resting-state activity interacts with the three names. These differences speak in favor of a specific causal impact of the resting state on specific stimuli rather than a nonspecific modulatory effect that remains unspecific to the type of stimulus.
We should, however, be aware that the data presented leave many questions open. One issue is the exact nature of the interaction; even a weak causal interaction must be explained by specific underlying neurophysiological mechanisms. Hence, at this point we cannot be fully clear about the causal nature of the rest-stimulus interaction. The kind of rest-stimulus causality I claim occurs in the example of Qin et al. (2013) is obviously a weaker form of causality than the one that claims a complete absence of stimulus-induced activity in the absence of resting-state activity. Hence, one may want to investigate different forms of causality (e.g., the four forms of causality as distinguished by Aristotle) in the future.
Another issue to consider is that the psychological implications of such rest-stimulus interaction are left completely open. The study by Qin et al. (2013) only focused on the neuronal differences of the stimulus-induced activity related to the own (and familiar and other) name during two different resting-state conditions, eyes closed and eyes open. In contrast, Qin and colleagues did not consider whether the differential neuronal reactivity of the resting state during eyes closed and open to the own name also impacted the psychological features, for example, perception.
For instance, subjects may have heard their own name in a more intense and attentive way during eyes closed when compared to eyes open (due to the additional visual-attentional distraction in the latter condition); this could be probed by reaction times in response to the own name which should then be faster during eyes closed than eyes open. The possible presence of such additional psychological differences may further support the assumption of causal rest-stimulus interaction.
What do these data imply with regard to the characterization of the brain as either passive or active? They suggest that the brain itself, through its resting-state activity level seems to provide an active contribution (that may come in different degrees) to its own neural activity. Such empirically grounded evidence speaks against a passive model of brain in all its versions, strong, moderate, and weak. We therefore shift our focus now to the active model of brain.
Part II: Active Model of Brain
Active Model of Brain Ia: Spatial Structure of Resting-State Activity
Early neuroimaging investigation using techniques such as fMRI and EEG focused on stimulus-induced activity including the brain’s response to sensorimotor, cognitive, affective or social stimuli or tasks. Recently, neuroimaging has shifted focus to the brain’s spontaneous activity and to its spatial and temporal structure. Initially, it was thought that spontaneous activity was contained to a particular neural network, the DMN (see also Klein, 2014). However, it soon became clear that spontaneous activity is pervasive throughout the whole brain.
Spontaneous activity has been observed in many different neural networks including the central executive network, the salience network, and the sensorimotor network (see Klein, 2014). Even in regions as dependent on external stimuli as the sensory cortices, there is spontaneous activity. The continuous neural activity occurring throughout the brain is spatially structured. Specific regions coordinate their ongoing resting-state activity, as measured in functional connectivity, thereby forming neural networks. This suggests that the brain’s resting-state activity can be characterized by a particular spatial structure, which when indexed by functional connectivity describes how two or more regions’ neural activities are synchronized and coordinated across time.
Active Model of Brain Ib: Temporal Structure of Resting-State Activity
In addition to its spatial structure, there appears to be quite an elaborate temporal structure in the brain’s intrinsic activity, which is manifest in its fluctuations in different frequency ranges. In the DMN, spontaneous fluctuations are characterized predominantly by low frequencies (<0.1 Hz). However, low- and high-frequency fluctuations in neural activity have been observed in sensory cortices, motor cortex, insula, and subcortical regions, such as the basal ganglia and thalamus (see Buckner, Andrews-Hanna, & Schacter, 2008; Freeman, 2003; Hunter et al., 2006; G. Shulman et al., 2009; R. Shulman et al., 2009; R. Shulman, Rothman, Behar, & Hyder, 2004; Wang, Duratti, Samur, Spaelter, & Bleuler, 2007).
How are low and high frequencies related to each other in the brain’s resting state? The empirical data suggest that low and high frequencies modulate each other (see the recent reviews by Canolty & Knight, 2010; Fell & Axmacher, 2011; Fries, 2009; Sauseng & Klimesch, 2008). For instance, Vanhatalo et al. (2004) conducted an EEG study of healthy and epileptic subjects during sleep. They used direct-current EEG to record low-frequency oscillations. All subjects showed infraslow oscillations (0.02–0.2 Hz) that were much stronger than during the awake resting state and even stronger than during stimulus-induced activity; these oscillations were detected across all electrodes—and thus the whole brain—without any specific, visually obvious spatial distribution evident.
Most interestingly, Vanhatalo et al. (2004) observed phase-locking or phase synchronization between the slow (0.02–0.2 Hz) oscillations and the amplitudes of the faster (1–10 Hz) oscillations. The amplitudes of the faster frequency oscillations (1–10 Hz) were highest during the negative phases of the slow oscillations (0.02–0.2 Hz). Such phase-locking of fast frequency oscillations’ amplitude to the phases of slower ones can also be described as phase-power coupling (see Canolty & Knight, 2010; Sauseng & Klimesch, 2008, for reviews). Generally, the coupling seems to occur in the direction from slow- to fast-frequency fluctuations (see Buzsáki, 2006; Buzsáki, Logothetis, & Singer, 2013). Such coupling from slow-frequency phase to faster-frequency amplitude represents one of the ways that different oscillatory patterns in neural activity can be related to one another. Such phenomena comprise the temporal structure of spontaneous activity.
Active Model of Brain Ic: Spatiotemporal Structure of Resting-State Activity
How is such temporal structure related to the spatial structure of the brain’s intrinsic activity? In a recent study, de Pasquale et al. (2012) observed that the DMN (and especially the posterior cingulate cortex) shows the highest degree of correlation with other networks in specifically the beta frequency range. The DMN seems to interact much more with the other networks than the latter do with each other. The reasons for that remain unclear but may be due, in part, to the central position of the DMN (and its midline structures) in the middle of the brain.
This fact likely makes the DMN prone to higher degrees of cross-network interaction than the other more laterally situated networks (e.g., the executive network or the sensorimotor networks; see Northoff, 2014a). Such cross-network interaction is dynamic and transient, and therefore it continuously changes. There are, for instance, alternate periods of low and high synchronization between DMN and other networks implying that cross-network synchronization and desynchronization go hand in hand.
These findings suggest that the spatial structure is closely linked to temporal dynamics, that is, oscillations in different frequency ranges (reviewed by Ganzetti & Mantini, 2013). Specifically, different neural networks may show different frequency ranges. For instance, Hipp, Hawellek, Corbetta, Siegel, and Engel (2012) observed that the medial temporal lobe is mainly characterized by theta frequency range, 4–6 Hz; the lateral parietal regions are rather featured by alpha to beta frequency range, 8–23 Hz; and the sensorimotor areas show even higher frequencies, 32–45 Hz. These findings demonstrate the close link between spatial and temporal dimensions in the spontaneous activity.
The tight link between the temporal and spatial structures is well reflected in the empirically informed theories of “nested oscillations” and “nested synchrony,” as observed in the resting-state activity by Monto (2012) using magnetoencephalography. Following him, nested oscillations describe the phase-phase/power coupling between low- and high-frequency fluctuations within one particular region. Neural synchrony extends beyond that, as it refers to the interregional coordination of nested oscillations in one particular region: the phase of a low-frequency fluctuation in one region may mediate the coupling of a high-frequency fluctuation in that region with a high-frequency fluctuation in another region.
In sum, these data show spatial and temporal structure in resting-state activity. The exact mechanisms and features of this spatiotemporal structure remain unclear, but we know that such spatiotemporal structure is highly dynamic rather than static, that is, its spatial and temporal configurations are subject to continuous change.
Active Model of Brain Id: An Active Rather Than Passive Model of Brain?
What do these findings of a dynamic spatiotemporal structure in the brain’s resting state tell us with regard to the model of brain? Let us start with the structure and organization of resting-state activity. I propose, based on empirical data, that in order to be processed, an external stimulus must interact with the resting state and its spatiotemporal structure. The external stimulus is integrated within the resting state and its spatial and temporal features. This allows the single discrete point in time and space of the stimulus to be linked with and integrated within the different time scales of the brain and its spontaneous activity.
How such rest-stimulus integration is related to mental features including consciousness of the respective stimulus remains unclear at this point. Therefore, the current chapter focuses exclusively on the brain and its neural activity independent of its role in mental features such as consciousness, which requires future elaboration in both neuroscientific and philosophical arenas.
At this point one may want to raise the issue of the model of a purely active brain that is not impacted at all by external stimuli . In such a case the spontaneous activity’s spatial and temporal structure should show exactly the same neuronal features during both a resting state and exposure to stimuli and tasks. Such a radical case would mean that the brain and its neural activity would be “self-evidencing” (Hohwy, 2014) rather than “world-evidencing” (as one might say analogously). However, that is not in accordance with the empirical data, which clearly show stimulus-induced or task-evoked activity to differ from the ongoing spontaneous activity.
A more moderate proposal would be to allow for stimulus-induced or task-evoked activity but in such way that the spontaneous activity is not altered. In that case one would for instance expect that the neural networks and the various frequency fluctuations, such as the spontaneous activity’s spatiotemporal structure, would remain the same during both resting state and exposure to stimuli and tasks. That is not the case, however.
Although present during sleep, rest, and tasks, the different neural networks and the frequency fluctuations change their relations with each other. For instance, as indicated above, the infraslow frequency fluctuations are particularly strong during sleep, less strong during awake rest, and even less strong during awake tasks. This suggests that one cannot really account for the brain and its spontaneous activity in terms of a purely active model in either a radical or even moderate form. For that reason I now propose what I call the spectrum model of brain.
Part III: Spectrum Model of Brain
Spectrum Model of Brain Ia: From Passive and Active Models to a Spectrum Model of Brain
Where do these findings leave us in our search for an empirically plausible model of brain? The data clearly do not support a passive model of brain nor are they in favor of an active model. We may thus need to opt for a third model in which the brain can be featured as both active and passive at the same time. I call such model the “spectrum model” of brain.
What do I mean by the concept of spectrum model? Neither the brain’s spontaneous activity alone nor the external stimuli by themselves determine stimulus-induced activity in the brain. Instead, it is their relation and thus their balance, that is, how they stand in relation to each other, that determines stimulus-induced activity. Since there is a spectrum or continuum of different possible balances or relations between spontaneous activity and external stimuli, I speak of a spectrum model of brain.
Specifically, such spectrum or continuum assumes that neural activity in the brain can result from different constellations or balances between internally and externally generated activity. Neural activity is consequently characterized as hybrid, stemming from both internal and external sources. The spectrum model concerns how the brain’s neural activity can involve different degrees of resting-state activity and, still, be shaped by external stimuli. One and the same neural activity level may be constituted by different degrees of resting-state activity at different times.
For instance, if the resting-state activity is rather strong and the external stimuli are weak, as for instance with a very low sound or weak visual feature, neural activity will be predominantly determined by the resting-state activity. If, in contrast, the resting-state activity is rather weak, the external stimuli (even if not that strong themselves) will have a stronger impact in shaping and constituting the brain’s neural activity.
Accordingly, the spectrum model of the brain suggested here is about the balance between the contributions of resting-state activity and external stimuli to the brain’s neural activity. Since various constellations in the balance between the resting state and external stimuli are possible, the brain’s neural activity can best be captured by a spectrum model that has room for configurations between the purely active and purely passive models discussed above.
Spectrum Model of Brain Ib: Middle and Extreme Cases in the Spectrum
The spectrum model carries major implications regarding how we can and cannot bring high-resolution definition to the concepts of resting-state and stimulus-induced activity. Within the context of the spectrum model, both are at best relative (rather than absolute) distinctions. The context of discussion can impact these issues. Whether one is speaking about operationalized concepts deployed in empirical investigations or trying to do justice to the basic physiological facts of the brain can make a difference to how one uses these concepts.
Operationally, we clearly need to distinguish between resting-state and stimulus-induced activities: resting-state activity is measured in a particular behavioral state, for example, eyes closed or open without any specific tasks or stimuli, whereas stimulus-induced activity is tested for by applying specific tasks or stimuli. In contrast, in a physiological context, the distinction between resting-state and stimulus-induced activity seems to evaporate. There is simply neural activity in the brain that may originate from different constellations of different sources.
According to the spectrum model, the brain’s neural activity is by default a hybrid or mixture between resting state and stimulus-induced activity. Neural activity is thus never determined 100 percent by either the resting state alone or by external stimuli alone. Neither extreme case occurs. The healthy brain operates apparently within the middle range of the spectrum or the continuum between different constellations of resting-state and stimulus-induced activity. This leaves open the more extreme cases of the spectrum. I suggest that we can find those in psychiatric disorders: here the constellation or balance between resting-state and stimulus-induced activity seems to shift from the middle of the spectrum toward its more extreme ends.
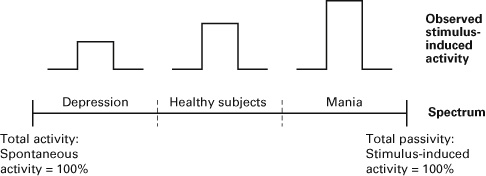
Let us consider schizophrenia and especially the auditory hallucinations that often occur with this disorder. Auditory hallucinations can be characterized by an abnormal increase in the level and functional connectivity of resting-state activity in the auditory cortex (see Alderson-Day et al., 2016; Northoff, 2014c, for recent reviews). In contrast, external stimuli such as auditory stimuli barely induce any activity change anymore in the auditory cortex of these patients. Neural activity in the auditory cortex in these patients is thus predominantly determined by the resting-state activity. Even during external stimulus processing, auditory cortical activity is predominantly constituted by the resting state rather than the external stimulus (see Alderson-Day et al., 2016; Northoff, 2014c; Northoff & Qin, 2011).
Another example occurs with depression. In depression, resting-state activity is abnormally elevated in the ACC. Symptomatically this is apparently related to ruminations and increased focus on the self and internal mental contents at the expense of external mental contents (see Northoff & Sibille, 2014a). Additionally, studies have demonstrated that the degree of activity change elicited by external stimuli is significantly reduced (if it can be observed at all) in depressed patients. The elevated resting-state activity is simply no longer as reactive to change related to external stimuli (see Grimm, Boesiger, et al., 2009; Grimm, Ernst, et al., 2009; Northoff, Wiebking, Feinberg, & Panksepp, 2011). Accordingly, as in the case of the auditory cortex in schizophrenia, neural activity in ACC during depression seems to be predominantly determined by the resting state rather than by the external stimuli.
Let us now consider the opposite extreme, neural activity being determined predominantly by external stimuli with relatively minor impact from the resting state. In healthy subjects this may, for instance, be the case in the presence of an abnormally strong external stimulus such as an extremely loud noise. The extremely strong stimulus will then override the impact of the resting state and determine the neural activity. A pathological instance of such a scenario may be the case of mania, the opposite of depression. In such a condition resting-state activity in ACC is reduced when compared to healthy subjects (Magioncalda et al., 2014; Martino et al., 2016), which seems to predispose it to react abnormally strongly to external stimuli.
What do these cases tell us for the spectrum model? In the case of elevated resting-state activity, the brain itself is highly active and determines its own neural activity, even in response to external stimuli. The opposite is the case when the external stimuli are extremely strong and/or the resting state is rather weak. In that case the external (or internal or neuronal) stimuli predominate in the brain, which by itself is then rather passive with a high degree of receptivity. Accordingly, these instances are cases that are shifted from the middle range of the spectrum toward being closer to the extreme ends of the spectrum between activity and passivity (see figure 1.1).
It is worth emphasizing that these extreme cases are not the norm. They are exceptions and can lead to major mental and psychological changes as in depression and schizophrenia. The healthy brain’s neural activity, in contrast, usually occupies a middle range in the spectrum where neural activity is codetermined by both resting state and external stimuli. The fluctuations in the degrees to which the resting state and external stimuli contribute to neural activity are consequently much more modest if not minor when compared to the major changes observed in the cases that shift more toward the extreme ends of the spectrum as in psychiatric disorders.
In sum, our brain’s neural activity is neither purely active nor completely passive. Instead, we may best account for the brain’s neural activity in terms of a spectrum model where neural activity is assumed to be codetermined by both the resting state and external stimuli in varying degrees and balances. Such a spectrum model entails a continuum between different degrees of activity and passivity in the brain’s neural activity.
Conclusion
In this chapter, we have discussed different models of the brain, passive, active, and spectrum, in the light of recent empirical findings in neuroscience as well as theoretical evidence as presented in the philosophy of science. The passive model of brain assumes that neural activity in general and stimulus-induced activity in particular are accounted for sufficiently (if not exclusively) by external stimuli themselves. Such a brain would only passively receive and process external stimuli but would not really contribute to shaping and constituting the resulting stimulus-induced activity. However, recent empirical evidence does not support such a model since the brain’s own activity, its resting state or spontaneous activity, causally impacts stimulus-induced activity.
The let us turn toward an active model of brain. In this case the brain itself is characterized by neural activity that occurs prior to and independent of external stimuli, the brain’s spontaneous or resting-state activity. Moreover, recent empirical evidence suggests that the brain’s spontaneous or resting-state activity exhibits a certain spatial and temporal structure as, for instance, in its different neural networks and frequency fluctuations.
Does the brain’s resting state or spontaneous activity determine stimulus-induced activity? As I demonstrate in the third section above, stimulus-induced activity seems to result from codetermination by both external stimuli and spontaneous activity, that is, world and brain. Stimulus-induced activity may then be determined by the spectrum or continuum of different possible relationships or balances between spontaneous activity, or brain, and external stimuli, world.
Beyond its empirical and theoretical plausibility, the spectrum model of brain carries far-reaching implications. The brain can no longer be regarded as merely passive in a purely empirical sense. Nor can the brain be regarded as purely active in an idealistic or constructivist sense. The spectrum model of brain builds on both ideas by integrating them into a spectrum or continuum of different relationships or balances between passive and active features. These relations or balances between passive and active features will prove central in our understanding of how the brain can yield consciousness, as is discussed in the second part of this book.