The spectrum model of brain refers to the hybrid nature of stimulus-induced activity as resulting from the impacts of both spontaneous activity and stimulus (see chapter 1). Although the spectrum model describes the neural activity in the brain including the relation between spontaneous and stimulus-induced activity, I left open whether that very same relation is also relevant for consciousness. That subject is the focus of this chapter.
Consciousness is not a homogenous entity. Instead, it is rather heterogeneous in that it includes different dimensions. One such dimension is the contents of consciousness such as the computer of which we are conscious (see Koch & Crick, 2003). Yet another dimension is the level of consciousness that refers to the arousal or the state of wakefulness of a person (see Laureys, 2005; see also Northoff, 2014b). The level of consciousness is often investigated in an indirect way. Patients who lose their level of consciousness as in disorders of consciousness such as sleep, anesthesia, or vegetative state (VS) may serve to reveal the neural correlates of the level of consciousness in an indirect way. I also pursue such indirect strategies in this chapter to investigate the relevance of the spectrum model of brain for the level of consciousness.
The general and overarching aim in this chapter is to investigate the relevance of both spectrum and interaction models of brain for the level of consciousness. My central argument is that the spectrum model of brain, on the basis of empirical evidence stemming from disorders of consciousness, is relevant for the level of consciousness. Specifically, I argue that the loss of the hybrid nature of stimulus-induced activity leads to loss of the level of consciousness in disorders of consciousness; this shifts these patients’ stimulus-induced activities from the “hybrid middle” toward the passive pole of the spectrum.
Part I: Stimulus-Induced Activity and the Level of Consciousness
Empirical Findings Ia: Preservation of Stimulus-Induced Activity during the Loss of Consciousness
Adrian Owen (Owen et al., 2006) scanned a patient in VS as featured by loss of consciousness in functional magnetic resonance imaging (fMRI) and let him perform specific cognitive tasks. While lying in the scanner, the VS patient was instructed to perform motor and visual imagery tasks (Owen et al., 2006): the patient was asked to imagine playing tennis. Surprisingly this yielded neural activity in the supplementary motor area (SMA), a region typically associated with the initiation of either physical or imaginary movements, in the VS patient. This region is related to movements as one would imagine or would perform them when playing tennis either mentally or physically. Most interestingly, the same region was activated in more or less the same way in healthy subjects. Hence, the VS patient was apparently able to perform a cognitive task as complex as imagining playing tennis. The same holds, analogously, in yet another task, spatial navigation when imaging walking through the rooms of your house. As in the case of the motor imagery task, the spatial navigation task activated the same region, the parahippocampal gyrus, in the patient as in the healthy subjects). The fact that both patient and healthy subjects recruited and activated the same regions during the two tasks suggests that the patient was able to perform the tasks in the same way as the conscious healthy subjects. Therefore, one can conclude that the patient herself must be conscious since otherwise she could not have performed the tasks and activated the same regions as the healthy subjects. The results were recently replicated in a larger sample by Monti et al. (2010). Analogous paradigms were here conducted in a larger group of fifty-four patients, of whom twenty-three were diagnosed with VS and thirty-one with minimally conscious state (MCS) (Monti et al., 2010). They had to perform the same tasks, imagining playing tennis and imagining walking from room to room in their own house. Five patients (four VS, one MCS) were indeed able to willfully modulate their neural activity during the tasks in a proper way: imagining playing tennis led to activation in the SMA in all five patients.
Moreover, as in the single patient study, imagining walking and spatially navigating in the own house induced neural activity changes in the parahippocampal gyrus in three VS patients and in one MCS patient. These neural patterns were again similar to those in the healthy control subjects. Since then, other investigations of cognitive tasks requiring task-related efforts and willful modulation have been conducted in VS and MCS—they have all demonstrated some preserved neural activity in the relevant regions in these patients (see table 3 in Laureys & Schiff, 2012, for an overview).
Taken together, these findings show that stimulus-induced activity seems to be still present during the loss of consciousness as in VS and MCS (see also Laureys & Schiff, 2012). This is further supported by other findings in anesthesia where, despite the loss of consciousness, subjects still show stimulus-induced activity in response to sensory stimuli and cognitive tasks (see MacDonald et al., 2015, for review).
Empirical Findings Ib: Abnormal Stimulus-Induced Activity during the Loss of Consciousness
The above described studies applied mainly cognitive tasks (and sensory stimuli) in order to elicit stimulus-induced or task-evoked activity. One may also apply a different kind of stimulus, a magnetic stimulus as applied by transcranial magnetic stimulation (TMS), to probe stimulus-induced activity. This was done by Rosanova et al. (2012), who combined TMS-based application of a magnetic pulse with continuous electrophysiological recording through electroencephalography (EEG).
Rosanova et al. combined TMS and EEG in five patients with VS, five patients with MCS, and two patients with locked-in syndrome (LIS) (patients with LIS, who are conscious but cannot communicate with the outside world) (Rosanova et al., 2012). Five of these patients were investigated several times in different stages of improvement, VS, MCS, and a fully conscious state (with only three patients in the last stage). Magnetic impulses were applied via TMS on the right and left medial frontal (superior frontal gyrus) and parietal (superior parietal gyrus) cortex to probe these regions’ neural activity changes in the resting state. The neural effects and especially the temporal and spatial spread and propagation of the magnetic stimulation were measured with simultaneous high-density, 264-channel EEG.
How did the VS patients’ resting state react to the TMS pulse? The VS patients showed a simple positive–negative EEG response that remained local, short, and did not change at all. This contrasted with the MCS patients, for whom the TMS impulse triggered a more complex EEG response that spread both spatially and temporally and also changed over time. The pattern in MCS resembled more closely the pattern in the two LIS patients than the pattern in VS patients. A similar pattern was observed in the longitudinal investigation of the five patients who were investigated several times throughout improvement. Their response patterns became more complex and thus spatially and temporally more propagated in the three patients who recovered from VS over MCS to the fully conscious state. In contrast, neither the more-extended spatial and temporal propagation nor more-complex response patterns could be observed in the two patients who remained in VS.
Taken together, as in the case of sensory and cognitive stimuli or tasks, the findings demonstrate that the TMS pulse still elicits stimulus-induced activity during the loss of consciousness. This is further corroborated by analogous TMS-EEG findings in both anesthesia and sleep (see chapter 15 in Northoff, 2014b, for an overview). However, stimulus-induced activity in response to the magnetic pulse is abnormal in that it lacks proper spatial and temporal distribution: there is neither spatial expansion nor temporal propagation of stimulus-induced activity across different regions and in time. Accordingly, stimulus-induced activity by itself is still present. However, the spatiotemporal features of stimulus-induced activity including its degree of spatial expansion (see chapter 7 for details) change and become abnormal, that is, they are reduced and local rather than expanded and global, during the loss of consciousness.
Empirical Findings IIa: Integrated Information Theory
Before discussing the above described results, I want to introduce briefly the two main theories of consciousness as discussed in current neuroscience, the integrated information theory (IIT) (Tononi et al., 2016) and the global neuronal workspace theory (GNWT) (Dahaene & Changeux, 2011; Dahaene et al., 2014). Both consider a specific neuronal mechanism, either information integration or globalization of neuronal activity, to be central, that is sufficient, for consciousness (see Koch et al., 2016, for an overview).
Edelman (2003, 2004) and Seth, Izhikevict, Reeke, and Edelman (2006) consider cyclic processing and thus circularity within the brain’s neural organization central for constituting consciousness (see also Llinás et al., 1998, 2002). Cyclic processing describes the reentrance of neural activity in the same region after looping and circulating in other regions via so-called reentrant (or feedback) circuits. This is, for instance, the case in primary visual cortex (V1): the initial neural activity in V1 is transferred to higher visual regions such as the inferotemporal cortex (IT) in feed-forward connections. From there it is conveyed to the thalamus, which relays the information back to V1 and the other cortical regions, implying thalamocortical reentrant connections (see also Tononi & Koch, 2008; as well as Lamme, 2006; Lamme & Roelfsema, 2000; van Gaal & Lamme, 2012). Consciousness and its contents are supposedly constituted on the basis of such feedback or reentrant connections that allow for cyclic processing (see also Edelman & Tononi, 2000).
What is the exact neuronal mechanism of the feedback or reentrant circuits? Reentrant circuits integrate information from different sources as associated with the neural activity in different regions and networks. This leads Tononi to emphasize the integration of information as the central neuronal mechanism in yielding and constituting the contents of consciousness. He consequently developed what he calls the “integrated information theory” (IIT; Tononi, 2004; Tononi & Koch, 2008; Tononi et al., 2016).
The IIT proposes the degree of information (as understood in a formal sense as in information theory rather than in common sense) that is linked and integrated central for consciousness: if the degree of integration of different information is low due to, for instance, disruption in functional connectivity between different regions, consciousness remains impossible. This is supported by experimental data that indeed show disruption of functional connectivity between different regions in various disorders of consciousness such as VS (Rosanova et al., 2012), non-REM (NREM) sleep (Qin et al., in revision; Tagliazucchi et al., 2013), and anesthesia (see Ferrarelli, Massimini et al., 2010).
To measure the degree of information integration across, for instance, different regions in the brain, Tononi and others (Seth et al., 2006, 2008; Seth, Barrett, & Barnett, 2011) developed specific quantifiable measures. Neurobiologically, Tononi postulates the integration of information to be particularly related to the thalamocortical reentrant connections. These reentrant connections process all kinds of stimuli from different sources and regions, thus remaining unspecific with regard to the selected content.
Such integration of different contents from different sources and regions is proposed to make possible the contents of consciousness and ultimately even their particular phenomenal quality, that is, qualia. In contrast, unconscious contents do not undergo such cyclic processing through the thalamus and the respectively associated information integration.
Empirical Findings IIb: Global Neuronal Workspace Theory
Another suggestion for the neural correlate of the contents of consciousness comes from Baars (Baars, 2005; Baars & Franklin, 2007) and others including Dehaene (Dehaene & Changeux, 2005, 2011; Dehaene, Changeux, Naccache, Sackur, & Sergent, 2006, for excellent overviews). They postulate global distribution of neural activity across many brain regions in a so-called global workspace central for yielding consciousness. If extended to the neuronal level, that very same global workspace may be found in specific circuits in the brain such as prefrontal and parietal cortex—Dehaene et al. (2014) therefore speak of a “neuronal global workspace theory,” the GNWT (see below in this section and the next).
The information and its contents processed in the brain must be distributed globally across the whole brain in order for them to become associated with consciousness. If, inversely, information is only processed locally within particular regions but no longer globally throughout the whole brain, it cannot be associated with consciousness anymore. The main distinction between unconsciousness and consciousness is then supposed to be manifest in the difference between local and global distribution of neural activity. Hence, the global distribution of neural activity is here considered a sufficient condition and thus neural correlate of consciousness (NCC).
Dehaene and Changeux (2005, 2011) take the assumption of a global workspace of consciousness as starting point and determine it in more neuronal detail when suggesting what they call the global neuronal workspace theory (GNWT). They postulate that neural activity in the prefrontal–parietal cortical network is central for yielding consciousness. More specifically, the prefrontal–parietal cortical network has to be recruited by the single stimulus in order to link and recruit its cognitive function that is central for instantiating consciousness: that makes possible the global distribution and processing of the stimulus, which, in turn, is supposed to be central for constituting the contents of consciousness.
The global workspace theory must be distinguished from other cognitive theories of consciousness. Some accounts link attention and/or working memory closely to consciousness and its contents (see, e.g., Lamme, 2006; Lamme & Roelfsema, 2000; van Gaal & Lamme, 2011). However, recent investigations have shed some doubt on attention and/or working memory being implicated in selecting the contents of consciousness (see Graziano & Kastner, 2011; van Boxtel et al., 2010a,b). This is supported by recent analyses that demonstrated consciousness and attention (and other cognitive functions) to occur independently of each other (see Faivre et al., 2014; Koch et al., 2016; Koch & Tsuchiya, 2012; Lamme, 2010; Tononi & Koch, 2015; Tsuchiya et al., 2015; van Boxtel et al., 2010a,b).
Stimulus-Induced Activity Ia: Spatiotemporal Expansion versus Spatiotemporal Constriction
We are now ready to discuss the above-described results on stimulus-induced activity and the neuroscientific theories of consciousness with regard to the spectrum model of brain. The first question I raise is the following: What do these results tell us about the role of stimulus-induced activity for the level of consciousness? The findings show that stimulus-induced activity as elicited by either cognitive or magnetic stimuli is still present in subjects who have lost their consciousness. This leaves us with two choices with regard to the relation between brain and mental features.
Either one assumes that the presence of stimulus-induced activity is simply not relevant for consciousness—the presence of stimulus-induced activity does not entail the presence of consciousness. Stimulus-induced activity can therefore not be conceived a sufficient condition, that is, an NCC. One would consecutively need to search for other neuronal features beyond stimulus-induced activity that are sufficient for constituting consciousness. Supposing that the patients are indeed unconscious, these neuronal features may then be impaired in disorders of consciousness.
Or, alternatively, one may suppose that the subjects showing stimulus-induced activity are conscious rather than unconscious—the presence of stimulus-induced activity entails the presence of consciousness. The presence of stimulus-induced activity especially in response to cognitive tasks such as motor imagery or spatial navigation (of the patient’s own house) can then be considered a neural correlate of consciousness, that is, NCC: the subjects must have understood the task requirement in order to elicit the observed stimulus-induced or task-evoked activity since otherwise, that is, in case they were unconscious and did not understand the instruction, they could not have elicited the observed neural activity changes.
The assumption that the patients must have understood the task instructions carries major clinical implications. The clinical diagnosis of the loss of consciousness and thus of VS in those patients showing the presence of stimulus-induced activity is simply wrong—neuronal evidence would then override clinical observation. This is indeed the conclusion current neuroscientists such as Adrian Owen and Stephen Laureys, two of the main investigators in the domain of VS, draw. They consider the presence of stimulus-induced or task-evoked activity in response to cognitive stimuli a neuronal marker for the presence of consciousness (see Bayne et al., 2016; Laureys & Schiff, 2012; Monti et al., 2010; Owen et al., 2006).
However, the data on TMS-induced stimulus-induced activity show that matters are not as clear-cut. Stimulus-induced activity is present here too. Does one therefore need to infer the presence of consciousness? Despite the presence of stimulus-induced activity in both conscious and unconscious subjects, there are nevertheless some differences between both groups. These differences concern mainly the spatiotemporal features of stimulus-induced activity.
The stimulus-induced activity in unconscious patients is neither as spatially extended (as to other regions and networks) nor as temporally propagated (as to more distal time points) in unconscious subjects. More generally, this means that stimulus-induced activity operates on a much more limited, that is, a more constricted, spatiotemporal range in the unconscious subjects—there is spatiotemporal constriction rather than spatiotemporal expansion.
Stimulus-Induced Activity Ib: Spatiotemporal versus Cognitive Features
What do I mean by “spatiotemporal expansion and constriction”? Roughly, the concept of spatiotemporal expansion describes the degree to which the stimulus-induced or task-evoked activity goes, that is, reaches, extends, or expands, beyond the specific discrete point in time and space at which the stimulus or task occurs and enters the brain.
Given that the stimulus-induced or task-evoked activity may extend, or expand, beyond the presence of the stimulus or tasks—the actual stimulus-induced or task-evoked activity may still be present even if the stimulus or task itself is already absent (see chapter 7 for more details on the concept of spatiotemporal expansion). If, in contrast, the presence of stimulus-induced or task-evoked activity is more limited in that it is tied and limited to the presence of the stimulus or task itself, one may speak of “spatiotemporal constriction.”
What do the concepts of spatiotemporal expansion and constriction imply for consciousness? The above described TMS-EEG findings suggest that the spatiotemporal features of stimulus-induced activity rather than its mere presence or absence are central for constituting the level of consciousness. Stimulus-induced activity must show a certain yet unclear degree of spatiotemporal expansion in order to allow for assigning a certain level of consciousness to the stimulus. In contrast, spatiotemporal constriction of stimulus-induced activity, even if present, leads to the absence of consciousness. The sufficient condition of consciousness, that is, its neural correlate or NCC, can then be found in the degree of spatiotemporal expansion of stimulus-induced activity rather than in the latter’s mere presence.
The advocate of stimulus-induced activity as NCC may now want to argue that such spatiotemporal characterization neglects its cognitive features. If the subjects can elicit stimulus-induced activity in response to cognitive tasks (see above), the subjects must have understood the cognitive demands, which is possible only if they are conscious. Stimulus-induced activity is taken here as index of cognitive function (Bayne et al., 2016) whose operation is possible only on the basis of consciousness being present.
That inference from the presence of cognitive function and operation to the presence of consciousness, as I say, a “cognitive–phenomenal inference,” is, however, an assumption that is not supported by empirical data. Cognitive functions, even complex ones, can still be present even in the absence of consciousness thus operating in an unconscious rather than conscious mode (see Faivre et al., 2014, Mudrik et al., 2014, for strong empirical support in this direction). One can therefore not infer from the presence of cognitively based stimulus-induced activity the actual presence of consciousness.
The cognitive–phenomenal inference may thus be fallacious, a cognitive–phenomenal fallacy. This puts the supposed attribution of consciousness to those vegetative patients showing cognitively based stimulus-induced activity on rather shaky if not implausible empirical grounds. We must consequently search for neuronal features other than those related to the cognitive features of stimulus-induced activity to reveal the NCC.
Stimulus-Induced Activity IIa: Spatiotemporal Framing of GNWT and IIT
The data suggest that the spatiotemporal features of stimulus-induced activity are central for consciousness. Specifically, stimulus-induced activity must show a certain yet unclear degree of spatiotemporal expansion (as distinguished from spatiotemporal constriction) to serve as NCC. Where and how is the spatiotemporal expansion of stimulus-induced activity coming from? This leads us back to the different neuroscientific theories of consciousness most notably the theories of integrated information theory (IIT) and the global neuronal workspace (GNWT).
The GNWT presupposes spatial expansion of stimulus-induced activity when postulating the need for prefrontal and parietal cortical recruitment during stimulus-induced activity. Moreover, it presupposes temporal propagation when suggesting a later event-related potential such as the P300 to signify the presence of consciousness (Dehaene et al., 2014; Dehaene & Changeux, 2011). Because globalization of neural activity is based on spatiotemporal features, for example the expansion to prefrontal-parietal regions and later potentials such as P300, I speak of spatiotemporal globalization. Importantly, it may be the degree of expansion in both space, that is, to other regions, and time, to later time periods such as P300, that may be central for instantiating consciousness. Instead of the cognitive functions associated with neuronal globalization, I suppose the underlying spatiotemporal features central for consciousness.
The same holds analogously in the IIT. The above described results on the magnetically based stimulus-induced activity serve Tononi to support his IIT (Koch et al., 2016; Tononi & Koch, 2015). Specifically, he supposes that the spatiotemporally constricted stimulus-induced activity in unconscious subjects’ indexes decreased integration of information. I now suppose that the decrease in spatiotemporal extension, that is, spatiotemporal constriction, is central for the loss of consciousness.
What Tononi and the IIT describe as decreased information integration may then be traced to decreased expansion of the spatial and temporal features of stimulus-induced activity, or the activity elicited by the TMS pulse. I thus suppose that information integration is about integration of spatial and temporal features as related to spatiotemporal expansion of stimulus-induced activity. In short, information integration is spatiotemporal integration (see chapter 7 for more details on this point).
Taken together, both GNWT and IIT are well compatible with the here-suggested spatiotemporal characterization of stimulus-induced activity as marker of consciousness. What they describe as globalization and information integration, as based on data, can be framed in terms of the spatiotemporal features of stimulus-induced or task-evoked activity. Therefore, I speak of spatiotemporal globalization and spatiotemporal integration to characterize GNWT and IIT in spatiotemporal terms. This leads me to a spatiotemporal model of consciousness, which will be developed in the second part of the book (chapters 7–8).
Stimulus-Induced Activity IIb: “Argument of Triviality”
The proponent of IIT and GNWT may now want to argue that the spatiotemporal framing is rather trivial. Given that any neural activity operates in time and space, both information integration and neuronal globalization are spatiotemporal by default. The assumption of globalization and information integration consecutively does not add any additional and novel information—the concepts of spatiotemporal globalization and spatiotemporal integration are trivial. This amounts to what I call the argument of triviality.
However, I reject the argument of triviality. The concepts of spatiotemporal expansion and spatiotemporal constriction show that one and the same stimulus or task can be processed in different ways, that is, expanded and constricted. The different ways can be distinguished on spatiotemporal grounds: the stimulus-induced activity can last longer or shorter and be spatially more or less distributed. Such spatiotemporal characterization of stimulus-induced activity seems to remain more or less independent of the stimulus or task itself. Instead, it seems to be dependent on the brain itself and, more specifically, what the brain itself adds and contributes to its own processing of the stimulus or tasks. Let me explicate this in the following way.
I propose that the brain’s own addition to the neural processing of stimuli or tasks consists in the spatiotemporal features of stimulus-induced or task-evoked activity. Specifically, the brain’s spontaneous activity adds a spatiotemporal dimension to its own processing of stimuli or tasks, tasks which then can be processed in either a spatiotemporally more expanded or more constricted way.
By adding the spatiotemporal dimension, the resulting stimulus-induced or task-evoked activity becomes more or less independent of the actual presence of the respective stimulus or task itself—the presence of stimulus-induced or task-evoked activity may then be well compatible with the absence of the actual stimulus or task itself. How is that related to consciousness? Based on the findings above, I postulate that the degree of spatiotemporal expansion of the stimulus-induced or task-evoked activity beyond the actual presence of the stimulus or task itself is directly related, that is, proportional to the level of consciousness.
The argument of triviality can thus be rejected. The brain can process one and the same stimulus in different ways that can be distinguished on spatiotemporal grounds, either constricted or expanded. The possibility of different ways of spatiotemporal processing of one and the same stimulus or task makes the spatiotemporal characterization of stimulus-induced or task-evoked activity anything but trivial. Moreover, the different ways of spatiotemporal processing provide a low-level dynamic neuronal mechanism for what IIT describes as integration (on an informational level) and GNWT as globalization (on a cognitive level). To neglect such a basic underlying level is to neglect that their difference, that is, whether constricted or expanded, leads to different behavioral outcomes (see chapter 7 for the argument of triviality).
Stimulus-Induced Activity IIc: From the “Spectrum Model” of Brain to Consciousness
Where and how does such spatiotemporal expansion of stimulus-induced activity originate if not in the stimulus or task itself? I indicated that the brain’s spontaneous activity contributes the spatiotemporal dimension, which I propose is central for consciousness. One would consequently expect differences in the brain’s spontaneous activity during the presence and absence of consciousness.
We recall that the spectrum model of brain (chapter 1) conceived stimulus-induced activity in a hybrid way. Stimulus-induced activity is neither exclusively and sufficiently associated with the stimulus itself (either a cognitive or a magnetic stimulus) nor with the spontaneous activity. Instead, the spectrum model postulates that stimulus-induced activity must be considered hybrid, that is, a mixture between the impact of the spontaneous activity and the impact of the stimulus.
What does the spectrum model of brain imply for consciousness? Following the spectrum model of brain, we need to search for the spontaneous activity and how it is altered in those unconscious subjects that show spatiotemporally constricted stimulus-induced activity in response to either cognitive or magnetic stimuli. This leads us back to the brain’s spontaneous activity and its temporal structure and how they are altered in disorders of consciousness.
Part II: Spontaneous Activity and the Level of Consciousness
Empirical Findings Ia: Spontaneous Activity and Its Temporal Structure—Power Law Exponent and Cross-Frequency Coupling
We have already discussed that the brain’s spontaneous activity can be characterized by an elaborate spatiotemporal structure (chapter 1). I now point out two temporal features, namely the power law exponent (PLE) and cross-frequency coupling (CFC) that, as we discuss below, will prove relevant for consciousness.
He (He, Zempel, Snyder, & Raichle, 2010) analyzed electrocorticographic data, measuring local field potentials of patients with epilepsy who underwent surgery. She analyzed data during REM sleep, slow- wave sleep (SWS), and in the awake state. First, she analyzed the power spectra (plotted in log-log coordinates) thus showing the PLE that refers to the relation in power between slower and faster frequency fluctuations; it followed the typical distribution with faster frequencies showing higher power and lower frequencies exhibiting lower power than slower frequencies. She also observed different peaks of power during SWS (0.8 Hz, 12 Hz) and the awake state (alpha, beta, theta).
Second, the PLE was estimated for five different subjects in the three conditions (awake, REM, SWS) for low (< 0.1 Hz) and high (>1–100 Hz) frequency ranges. Interestingly, no significant differences in the PLE could be yielded among the three different states (awake, REM, SWS, as calculated by effect of arousal state on the PLE).
Next, He (He et al., 2010) investigated the phase-amplitude coupling between slow and fast frequencies; the strength of such CFC was formalized by a modulation index (MI). The MI was obtained for all possible frequency pairs for all electrodes in 1-Hz frequency steps. Significant values of the MI, and thus nested frequencies (signifying cross-frequency, phase-amplitude coupling), were obtained for all three states (wakefulness, REM, SWS) across the entire frequency range.
She then demonstrated that the preferred phase in the slower frequency for coupling to the amplitude of the higher frequency clustered around the peak and the trough of the phase. The peak and trough reflect the most positive and most negative parts of the phase or cycle duration where it shows the highest and the lowest degrees of excitability.
Taken together, the data show that the brain’s spontaneous activity is not random but rather highly structured by showing strong CFC with multiple phase-amplitude coupling from slow to fast frequency fluctuations. He (He et al., 2010) consequently assumed that this temporal structure operates in the background, against which any subsequent changes in neural activity, as for instance by external stimuli or tasks, occur and can be measured. Moreover, it seemed that the spontaneous activity’s temporal structure as indexed by both PLE and CFC is more or less preserved during the loss of consciousness, as in sleep.
Empirical Findings Ib: PLE and CFC in Sleep and Anesthesia
Is the spontaneous activity’s temporal structure as indexed by PLE and CFC really preserved during the loss of consciousness? Tagliazucchi et al. (2013) conducted an fMRI study in sixty-three subjects during the different stages of NREM sleep. Here the power spectrum as the relation between different frequencies was measured with the Hurst exponent (in a voxel-based analysis) during the transition from wakefulness to the different stages of NREM sleep (N1–N3 with the latter being the deepest).
Interestingly, they observed significant decay of the global Hurst exponent of the whole brain from wakefulness (where it was the highest) over N1 and N2 to N3, the deepest stage of NREM sleep. This means that the power spectrum and thus the degree of scale-free activity (as measured by PLE/DFA) significantly and progressively decrease during the different stages of NREM sleep. Such global decrease of the power spectrum was also observed in anesthesia using fMRI (Zhang et al., 2017). Zhang et al. (2017) observed a global decrease of the PLE across the whole brain in the anesthetic state when compared to the awake state in the same subjects. This went along with a decrease in neuronal variability in the whole brain—the spontaneous activity thus did no longer exhibit as many spontaneous changes anymore in the anesthetic state.
What about the spatial and temporal features of the power spectrum decrease during the loss of consciousness? Let us start with the spatial dimension. More regionally specific effects were tested in a second step in the study by Tagliazucchi et al. (2013). Voxelwise comparison of the four stages (wakefulness, N1–N3) showed significant regional differences in the Hurst exponent in the frontal and parietal regions (that are associated with neural networks such as the DMN and the attention network) during N2 and N3 when compared to wakefulness (and N1). The deepest sleep stage, that is, N3, was associated with more widespread global decrease in the Hurst exponent in the frontal and parietal regions as well as in the occipital cortex and the visual network. In contrast, no major differences in the Hurst exponent could be detected in N1 when compared to wakefulness.
In addition, they also focused on the temporal dimension and, more specifically, on the different frequency ranges. The fMRI measures mainly infraslow frequency fluctuations (IFF) in the range between 0.01 to 0.1 Hz, whereas the EEG records faster frequencies ranging from 1 to 4 Hz (delta) to gamma (30–180 Hz). Tagliazucchi et al. (2013) now related the Hurst exponent as obtained in the infraslow range of fMRI (0.01–0.1 Hz) to the power of the delta frequency band (1–4 Hz) as recorded in a simultaneous EEG. The Hurst index (from fMRI) in the frontoparietal regions associated with DMN and attention networks correlated negatively with delta power (from EEG as averaged across all channels): the lower the Hurst index in the infraslow range (0.01–0.1 Hz), the higher the delta power (1–4 Hz).
These findings suggest a specific significance of the IFF, that is, 0.01 to 0.1Hz, for the level of consciousness (see also Northoff, 2017a, for a recent review). The decrease in the power spectrum (as measured with Hurst) indicates a power decrease in the very slow frequency ranges, such as 0.01 to 0.027 Hz that is featured as slow-5. Is slow-5 power specifically decreased during the loss of consciousness? This was indeed observed in anesthesia by Zhang et al. (2017), who found significant decrease in the power of slow-5 variability (rather than in slow-4, e.g., 0.027–0.073 Hz) during the anesthetic state.
In sum, these studies in sleep and anesthesia show significantly decreased power spectrum with specific power decrease in the infraslow ranges of 0.01 to 0.1 Hz in the whole brain during the loss of consciousness. However, the findings suggest that there is some grading from more local to global neuronal changes in both spatial and temporal dimensions during the transition from the awake to the deeply unconscious state. Most notably, the findings indicate special significance of especially the very slow fluctuations, the IFF as featured by their long cycle durations (with up to 100s), for the level of consciousness (see Northoff, 2017a, for more details).
Spontaneous Activity Ia: Temporal Relation and Integration
What do these findings tell us about the spontaneous activity and its spatiotemporal structure with regard to the level of consciousness? The spatiotemporal structure of the spontaneous activity can by itself be characterized by spatiotemporal relation and integration. We now explore the exact nature of these findings below.
The spontaneous activity shows an elaborate temporal structure. This temporal structure can be measured by the PLE that indexes the temporal relation in the power between infraslow, slow, and faster frequencies. The strongest power can be observed in the very lowest frequency ranges, that is, the IFF, whereas slow and faster frequency ranges show less power. Together, this results in the typical power spectrum that can be characterized as scale-free activity (He, 2011, 2014).
This temporal relation in the power among infraslow, slow, and faster frequencies is changed in altered states of consciousness. The above-described findings suggest that the power in the IFF in general and specifically in their slowest frequency range such as the slow-5 (0.01–0.027 Hz) is diminished in unconscious states such as anesthesia or deep sleep. That leads to decrease in PLE: the decrease in the power of the infraslow frequencies weakens the “temporal basement” upon which neural activity related to faster frequencies stands. The data suggest that that temporal basement is central for consciousness.
Taken together, these findings show that the spontaneous activity’s temporal power relation among infraslow, slow, and faster frequencies is relevant for the level of consciousness. One may now want to argue that these findings only show the relevance of temporal relation but not of temporal integration. The relation in the power among the different frequencies is altered, but that does not yet tell us anything about temporal integration, that is, how the different frequencies are linked and integrated with each other.
For that, we need to investigate how the different frequencies are coupled and linked to each other as in cross-frequency coupling (CFC). The above described data show that there is indeed major CFC in the conscious brain that speaks in favor of temporal integration (that extends beyond simple temporal relation). Is such temporal integration as indexed by CFC also relevant for the level of consciousness? Abnormal, such as decreased CFC within the faster frequency ranges between 1 Hz and 60 Hz has been reported in anesthesia (see Lewis et al., 2012; Mukamel et al., 2011, 2014; Purdon et al., 2013, 2015). This leaves open, however, CFC between the infraslow and slow/faster frequency ranges. That has been tested in multiple simultaneous fMRI-EEG studies in conscious subjects that most often demonstrated a relation between infraslow frequencies (as in fMRI) on the one hand and delta and alpha frequencies (as in EEG) on the other (see Sadaghiani, Scherringa, et al., 2010).
As described above, unconscious subjects in sleep show IFF-delta power–power relationship though in an inverse, that is, negative way (see discussion directly above). However, whether that power–power relation between IFF and delta can be traced to the IFF-phase–delta-amplitude relation as usually measured in CFC remains unclear at this point. If so, one would suppose not just abnormal temporal relation but also a decrease in temporal integration in unconscious subjects. Such decrease in temporal integration may limit and thus constrict temporal continuity of the spontaneous activity across its different frequency ranges. This amounts to what I describe as temporal constriction. That very same temporal constriction of the spontaneous activity’s temporal continuity may be central for the loss of consciousness as the authors of the above study suggest themselves (Tagliazucchi et al., 2013).
Spontaneous Activity Ib: Spatial Relation and Integration
What about spatial relation and integration? Spatial relation is usually indexed by functional connectivity that is based on a statistically based correlation between two (or more) regions’ time series during the resting state. Several investigations have demonstrated reduced functional connectivity in disorders of consciousness such as sleep, anesthesia, and VS (see, e.g., Huang et al., 2014a,b, 2016; Qin et al., submitted; Vanhaudenhuyse et al., 2011). This suggests reduced spatial relation in spontaneous activity during the unconscious state.
Moreover, the findings in both sleep and anesthesia as described above clearly demonstrate the global nature of the PLE/Hurst reduction occurring across all regions and networks during the fully unconscious state. This suggests reduced spatial relation and integration of spontaneous activity in unconsciousness. The findings in sleep demonstrate that these changes in spatial integration occur in a graded way with more local reduction in N2 and stronger more globalized reduction in N3 (see discussion in the previous section).
Overall, the spontaneous activity shows decreased spatiotemporal integration in disorders of consciousness such as sleep, anesthesia, and VS. This suggests that the spontaneous activity’s spatiotemporal relation and integration are relevant for the level of consciousness.
Spontaneous Activity Ic: Spatiotemporal Integration of Stimulus-Induced Activity within Spontaneous Activity
How are the spontaneous activity’s spatiotemporal relation and integration related to the spatiotemporal expansion (or constriction) of stimulus-induced activity? Unfortunately, spontaneous activity and stimulus-induced activity have rarely been investigated together in conjunction in unconscious subjects.
One notable exception is presented by Huang et al. (2014a). They observed that the significant decrease in functional connectivity between the anterior (such as, perigenual anterior cingulate cortex) and posterior (such as, posterior cingulate cortex) cortical midline regions is related to the reduced stimulus-induced activity in response to self-related stimuli (see chapter 4 for details). This suggests decreased interaction and thus decreased spatiotemporal integration of stimulus-induced activity within the spontaneous activity and its spatiotemporal structure.
This assumption of direct relation of impairments in the spontaneous activity’s spatiotemporal relation and integration with changes in stimulus-induced activity is further supported by additional data. These data show decreased nonadditive rest–stimulus interaction in disorders of consciousness (Huang et al., 2017) (see chapter 2 for details on nonadditive interaction). Although rather tentative, these data suggest that the changes in the spontaneous activity’s spatiotemporal relation and integration impact the stimulus-induced activity including its spatiotemporal expansion (or constriction).
One may want to suggest the following hypothesis: the more spatiotemporal relation and integration are reduced in the spontaneous activity, the lower the degree to which the stimulus-induced activity can be spatiotemporally expanded (and the higher the degree of its spatiotemporal constriction). I therefore propose that the low degrees of spatial expansion and temporal propagation of the magnetically based–stimulus-induced activity (during TMS-EEG) in unconscious subjects may be traced to changes in their spontaneous activity’s spatiotemporal relation and integration.
How can we investigate this hypothesis? One could, for instance, investigate the PLE/Hurst and CFC in the EEG resting state and prestimulus periods (e.g., prior to the magnetic pulse/stimulus); they could then be correlated with the degrees of spatial expansion and temporal propagation of the magnetically based–stimulus-induced activity. Lower values in the spontaneous and prestimulus activity’s PLE and CFC should lead to lower degrees of spatial expansion and temporal propagation of the subsequent magnetically based–stimulus-induced activity. Demonstration of that, however, remains to be done in the future.
In sum, I hypothesize that spontaneous and stimulus-induced activity are closely related to and integrated within each other on spatiotemporal grounds. I therefore speak of spatiotemporal integration between spontaneous and stimulus-induced activity that is ultimately based on the spatiotemporal relation and integration within the spontaneous activity itself. Most important, the data suggest that such spatiotemporal integration between spontaneous and stimulus-induced activity is relevant for and, even stronger, proportional to the level of consciousness.
Spontaneous Activity IIa: Spectrum Model of Brain and Consciousness—Passive Model
What does the need for spatiotemporal integration between spontaneous and stimulus-induced activity imply for the relevance of the spectrum model of brain for the level of consciousness? The main point of the spectrum model of brain is the hybrid nature of stimulus-induced activity that supposedly results from the impact of both spontaneous activity and stimulus. The data suggest that the hybrid nature of stimulus-induced activity seems to be central for constituting the level of consciousness. Let us specify that in the following discussion.
Stimulus-induced activity is still present in the absence of consciousness. What, in contrast, is not present during the absence of consciousness is the spatiotemporal expansion of stimulus-induced activity. That very same spatiotemporal expansion seems to be based on the spontaneous activity and its spatiotemporal relation and integration. If the latter two are altered, as seems to be the case in disorders of consciousness, stimulus-induced activity can no longer be spatiotemporally extended; this, apparently, makes it impossible to assign consciousness to the respective stimulus (as either cognitive, magnetic, or otherwise).
The changes in the spontaneous activity in the disorders of consciousness entail that its (altered) spatiotemporal structure cannot impact subsequent stimulus-induced activity anymore. There is less spatiotemporal integration in spontaneous activity, which makes stimulus-induced activity less hybrid during the loss of consciousness. Instead of being codetermined by both spontaneous activity and stimulus, stimulus-induced activity is solely (and exclusively in most extreme cases) determined by the stimulus itself: the hybrid balance between spontaneous activity and stimulus is shifted toward the latter, which makes stimulus-induced activity less hybrid and less spatiotemporally extended.
How does that relate to the spectrum model of brain? The spectrum model of brain postulates that stimulus-induced activity consists of different components including contributions from both stimulus and the brain’s spontaneous activity. Hence, stimulus-based coding is hybrid by default. However, the balance between both components, stimulus and spontaneous activity, may vary. If the contribution from the stimulus predominates, the resulting stimulus-induced activity shifts more toward the passive pole of the spectrum. If, in contrast, the contribution of the spontaneous activity predominates, the stimulus-induced activity can be located more on the active end of the spectrum.
I distinguished among different models on the passive end of the spectrum of stimulus-induced activity: (1) the weakly passive model supposes modulatory but not causal impact of spontaneous activity on stimulus-induced activity; (2) the moderately passive model suggests no impact of spontaneous activity on stimulus-induced activity; and (3) the strongly passive model assumes that there is no spontaneous activity at all any more (see chapter 1 for a generalized discussion).
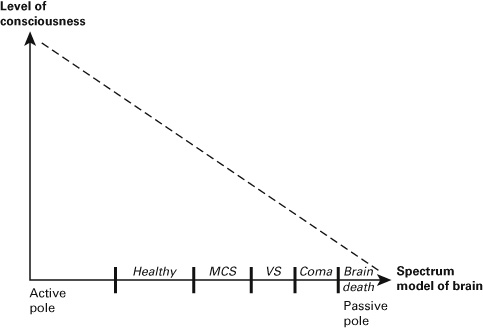
I now postulate that the different passive models of stimulus-induced activity are well in accordance with the data on the different degrees of spatiotemporal expansion (as in the TMS-EEG study) stages during the loss of consciousness as experienced in MCS, VS, and coma. The above-described TMS-EEG results show that magnetically based–stimulus-induced activity is more spatiotemporally expanded in the MCS subjects and less so in the VS patients. Stimulus-induced activity in the MCS subjects may thus correspond to the weakly passive model.
In contrast, stimulus-induced activity in VS subjects may rather presuppose the moderately passive model. Finally, the even more extreme cases of coma and especially of brain death show no spatiotemporal expansion of stimulus-induced activity at all anymore resulting in spatiotemporal constriction or total absence of stimulus-induced activity—they may thus come close to the strongly passive model.
What does this tell us about the level of consciousness in the healthy subject? The hybrid nature of stimulus-induced activity and ultimately the spectrum model of brain are relevant for the level of consciousness: the more hybrid the stimulus-induced activity as resulting from the impact and spatiotemporal integration between spontaneous activity and stimulus, the higher the level of consciousness (see figure 4.1).
Figure 4.1 Spectrum model of brain and the level of consciousness.
Spontaneous Activity IIb: Spectrum Model of Brain and Consciousness—Active Model
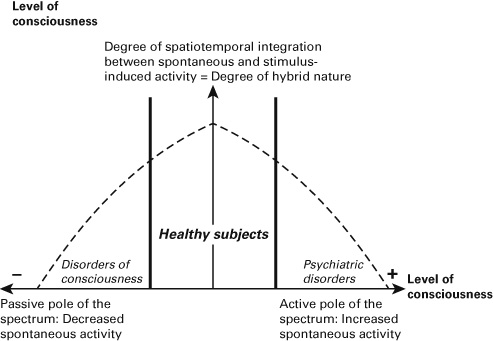
One may now want to argue, however, that the assumption of the hybrid nature of stimulus-induced activity neglects the opposite end, the active pole of the spectrum model. In that case, stimulus-induced activity is determined solely by the spontaneous activity itself without any impact by the stimulus anymore. One then reverts to the opposite extreme and neglects the middle ground, which, as I suggest, is central for consciousness.
Analogously to the passive pole, one may want to distinguish among different stages or degrees on the active end of the spectrum. In the case of a weakly active model, the spontaneous activity can no longer be affected by stimulus-induced activity in a causal way; however, the former can still be modulated by the former in a modulatory way. That very same modulation disappears in the moderately active model: the spontaneous activity can no longer be modulated by stimulus-induced activity even in a noncausal way. Finally, the strongly active model postulates the absence of any stimulus-induced activity with the spontaneous activity taking over its role (see figure 4.2).
Figure 4.2 Spectrum model of brain and its relation to consciousness.
How can we relate the different active models to specific disorders? We have seen that the disorders of consciousness entail a passive model where the impact of the spontaneous activity on subsequent stimulus-induced activity decreases and vanishes—this, as I suppose, is directly proportional to the degree to which consciousness is lost. What about the opposite end, the active models? I postulate that psychiatric disorders are a perfect test case here.
Depression can be characterized by increased spontaneous activity, specifically in the midline regions and the DMN (see Northoff, 2015a, 2016a,b, for review), but the stimulus-induced activity in these regions is decreased and, it can be hypothesized, may no longer affect the spontaneous activity in these regions in a causal way. One may thus want to assume a weakly active model in depression. Symptomatically, the lacking causal impact of stimulus-induced activity on spontaneous activity may, for instance, be reflected in the fact that, behaviorally, these patients no longer react properly to external stimuli and continue ruminating on their own thoughts (even when confronted with external stimulation).
Bipolar disorder can be characterized by mania and depression that show opposite motor behavior with psychomotor retardation in depression and psychomotor agitation in mania. The manic patient runs around, whereas the depressed patient sits in the corner. That seems to be related to neuronal variability in the sensorimotor network: manic patients show increased resting state neuronal variability in this network, whereas the same measure is decreased in depressed subjects (Martino et al., 2016; Northoff et al., 2017). Importantly, the psychomotor behavior cannot be modulated by any external stimuli at all. Neuronally, this may be related to the fact that the depressed patients’ spontaneous activity in the sensorimotor network is no longer modulated in either a causal or noncausal way by the external stimuli. The depressed patients’ spontaneous sensorimotor cortical neuronal variability may thus correspond to the moderately active model of stimulus-induced activity.
Yet another example is displayed in schizophrenia. Here the patients can experience auditory hallucinations that are related to increased activity in the resting state (Alderson-Day et al., 2016; Northoff & Duncan, 2016). Despite the absence of any stimulus-induced activity, patients nevertheless experience voices in the external world, which, in the healthy subjects, are usually associated with stimulus-induced activity. Hence, the schizophrenic patients’ spontaneous activity takes over the role of stimulus-induced activity. This corresponds to the strongly active model of stimulus-induced activity where it disappears while its role is taken over by the spontaneous activity itself.
One may now want to argue that the psychiatric examples fit well with the active end of the spectrum model of brain. In contrast, they are not relevant for consciousness because it is the case at the opposite end of the spectrum, its passive pole. However, such supposition is to neglect that psychiatric disorders also show changes in consciousness. Depressed patients show increased awareness of their own inner thoughts and beliefs, and the same holds for schizophrenic patients with regard to their auditory hallucinations.
One may therefore tentatively suggest that the level of consciousness in these patients is increased (rather than decreased). Hence, rather than rejecting the relevance of the spectrum model of brain for consciousness, psychiatric disorders support the model by showing the opposite extreme in the level of consciousness (Northoff, 2013, 2014b).
Conclusion
Is the spectrum model of brain relevant for the level of consciousness? I have demonstrated that stimulus-induced activity shows decreased spatiotemporal expansion during the loss of consciousness. That, as it may be hypothesized, is closely related to changes in the spontaneous activity and its degree of spatiotemporal relation and integration. Therefore, the spontaneous activity can no longer impact stimulus-induced activity any more as in the healthy subject.
The resulting stimulus-induced activity consequently shifts more toward the passive pole of the spectrum: it is more strongly dominated by the stimulus itself than by the brain’s spontaneous activity. Different models of stimulus-induced activity on the passive end of the spectrum, weak, moderate, and strong, may thus correspond to different degrees in the loss of consciousness such as MCS, VS, and coma. Accordingly, the spectrum model of brain conforms well to the data during the loss of consciousness and can therefore be considered relevant for consciousness.
Finally, the data and their explanation by the spectrum model of brain suggest that stimulus-induced activity and spontaneous activity take on different roles for consciousness. The stimulus-induced activity and its underlying nonadditive rest–stimulus interaction may serve as a sufficient neural condition of actual consciousness, that is, as NCC. In contrast, the spontaneous activity may rather provide the necessary condition of possible consciousness, a neural predisposition of consciousness (Northoff, 2013, 2014b; Northoff & Heiss, 2015, Northoff & Huang, 2017). That neural predisposition of consciousness, however, needs to be explained in greater detail. For that, we now shift our focus to yet another model of brain, the interaction model and how it relates to consciousness.