In the second part of chapter 2, I introduced the interaction model of brain that concerns the nature of the interaction between spontaneous activity and stimuli as being nonadditive rather than merely additive or parallel. The interaction model describes the neural activity in the brain including the relation between spontaneous and stimulus-induced activity. In contrast, I left open whether the interaction model is also relevant for consciousness; that is the focus in this chapter.
The general and overarching aims of this chapter are to investigate the relevance of the interaction models of brain for consciousness. The argument is that the interaction model of brain is indeed relevant for consciousness.
Our first specific aim is to discuss the interaction of model of brain and most notably the nonadditive rest–stimulus interaction in the context of recent findings from disorders of consciousness. Based on empirical evidence, I propose that the degree of nonadditive interaction between spontaneous and stimulus–induced activity is directly relevant for the spatiotemporal expansion of stimulus-induced activity, including its association with consciousness. Even stronger, the empirical data suggest that the degree of nonadditive interaction may serve as a neural signature and thus as a neural correlate of consciousness (NCC).
The second specific aim of this chapter entails showing the central role of spontaneous activity for nonadditive rest–stimulus interaction and its relation to consciousness. I argue that the spontaneous activity is a necessary condition of possible consciousness and thus a neural predisposition of consciousness (NPC). Conceptually, the NPC can be enriched by the concept of capacities as understood by Nancy Cartwright—I thus argue for a capacity-based approach (rather than a law-based approach) to the brain and its relation to consciousness.
Part I: Interaction Model and Consciousness
Empirical Findings Ia: Stimulus Differentiation and the Level of Consciousness
Building on a recent study by Qin et al. (2010), Huang (Huang et al. 2014, 2017a,b) from our group investigated stimulus–induced activity in the vegetative state (VS) (i.e., unresponsive wakefulness [URWS] as it is called these days) during self- and non-self-related stimuli. Instead of letting subjects listen to their own (and other) name(s) (as in Qin et al., 2010), subjects now had to perform an active self-referential task wherein they had to refer to themselves, that is, each to his or her own self.
Two types of questions, autobiographical and common sense, were presented via audio. The autobiographical questions asked for real facts in the subjects’ lives as obtained from their relatives. This required subjects to actively link the question to their own selves, a self-referential task. The control condition consisted of common-sense questions whereby subjects were asked for basic facts such as whether one minute is sixty seconds. Instead of giving a real response via button click (because it was impossible with these patients), the subjects were asked to answer (mentally not behaviorally) with “yes” or “no.”
Huang first compared autobiographical and common-sense questions in healthy subjects. As expected on the basis of previous findings about the involvement of midline regions in self-relatedness (Northoff, 2016c,d, 2017a,b; Northoff et al., 2006), this yielded significant signal changes in the midline regions, including the anterior regions such as the perigenual anterior cingulate cortex (PACC) extending to ventromedial prefrontal cortex and the posterior regions such as the posterior cingulate cortex (PCC): the activity changes were significantly stronger in the self-related condition when compared to the non-self-related condition.
What did the brains in the VS patients now show in the very same regions? They showed signal changes in these regions that were reduced compared to those in healthy subjects. More specifically, the degree of neural differentiation between self- and non-self-related conditions was much lower.
How are these signal changes now related to consciousness? As in the study by Qin et al. (2010), a significant correlation in anterior midline regions was observed. The midline regions’ activity including the PACC, dorsal anterior cingulate cortex, and PCC correlated with the degree of consciousness (as measured with the Coma Recovery Scale–Revised). It seems that the more signal changes in these regions differentiated neuronally between self- and non-self-referential conditions, the higher the levels of consciousness that patients exhibited. Thus we have observed a direct relation between the degree of neuronal self–nonself differentiation and the level of consciousness in anterior and posterior midline regions.
Empirical Findings Ib: From Spontaneous to Stimulus-Induced Activity
What about the resting-state activity in the same patients? For our purposes, one wants to know whether the diminished responses to self-specific stimuli are related to changes in the resting state in the relevant midline regions. For that, Huang, Dai, et al. (2014) also investigated the resting state, for example, its functional connectivity and low-frequency fluctuations, in exactly those regions that showed diminished signal differentiation, that is, stimulus-induced activity, during the self-referential task.
As in the previous studies, the VS patients showed significantly reduced functional connectivity from the PACC to the PCC in the resting state. In addition, the neuronal variability of the amplitude in the slowest frequency range (i.e., slow-5 as from 0.01 to 0.027 Hz) was significantly lower in both PACC and PCC in VS when compared to healthy subjects.
Given that we investigated exactly the same regions during both resting state and task, this strongly suggests that the resting-state abnormalities in these regions are somehow related to the earlier described changes during the self-referential task. This was further supported by correlation analysis: the higher the neuronal variability in the slow frequency range (i.e., slow-5) in PACC and PCC, the higher the degree of neuronal signal differentiation between self- and non-self-related conditions in the same regions during stimulus-induced activity.
Taken together, these findings suggest that the stimulus-induced activity is mediated by spontaneous activity and thus affected by the latter’s changes during the loss of consciousness. What remains unclear is their exact interaction, that is, rest–stimulus interaction during, for instance, the loss of consciousness. We discussed in the interaction model that spontaneous activity and stimulus may interact in a nonadditive way. If such nonadditive interaction is central for consciousness, one would expect it to be altered, that is, decreased and ultimately as additive during the loss of consciousness. This is indeed the case, as is supported by additional empirical data that we discuss in the next section.
Interaction Model Ia: Stimulus-Induced Activity and Rest-Stimulus Interaction
What do these findings tell us with regard to stimulus-induced activity and its relevance for the level of consciousness? Let us start with the neuronal side of things. The findings by Huang, Dai, et al. (2014) show the following: (1) there are changes in stimulus-induced activity in VS subjects who are unable to properly differentiate between self- and non-self-related stimuli, in specifically midline regions such as PACC and PCC; (2) changes in the resting state’s spatial structure of VS subjects as reflected in reduced PACC–PCC functional connectivity; (3) changes in the resting state’s temporal structure of VS patients as in decreased neuronal variability in PACC and PCC; (4) correlation of the resting state’s reduced neuronal variability with the decreased neuronal self–nonself differentiation during stimulus-induced activity; (5) correlation of decreased neuronal self–nonself differentiation with the level of consciousness.
Taken together, these findings suggest that the abnormally reduced neuronal variability in spontaneous activity is related to the decreased neuronal self–nonself differentiation during stimulus-induced activity. This suggests abnormal interaction between spontaneous activity and self- and non-self-related stimuli. That, in turn, seems to make impossible the association of stimulus-induced activity with consciousness; this is supported by the correlation between self-related activity and the level of consciousness as observed by Huang, Dai, et al. (2014).
However, my thesis on the effect of abnormal interaction on the loss of consciousness so far relies solely on correlational evidence. Correlation itself does not prove interaction let alone causal relation between spontaneous activity and stimulus-induced activity (as for instance required by both the spectrum and interaction models of brain). The argument of abnormal direct and most likely causal interaction thus rests on rather questionable empirical grounds, that is, correlation. Therefore, we cannot really suppose abnormal interaction between spontaneous and stimulus-induced activity in VS.
In order to support the assumption of abnormal interaction, we therefore need neuronal measures that directly index the interaction between spontaneous and stimulus-induced activity. One such neuronal measure is trial-to-trial variability (TTV). TTV measures the degree of changes in ongoing neuronal variability during stimulus-induced activity in relation to the degree of variability at stimulus onset (or the preceding prestimulus period). By measuring the relative change in neuronal variability, TTV accounts for how the stimulus (and stimulus-induced activity) causally impacts and changes the ongoing variability of the spontaneous activity. Therefore, TTV can be considered an indirect index of causal interaction between the resting state and stimulus.
Let us now consider the TTV in the above discussed data on self–nonself differentiation? Huang et al. (2017a) first investigated a healthy sample with the same paradigm of self and nonself stimuli as in VS. This yielded significantly stronger TTV reduction in specifically PACC and PCC during self-related stimuli when compared to non-self-related stimuli. The same finding was confirmed in a second data set on healthy subjects using the same paradigm. This suggests that self–nonself differentiation is directly related to the interaction between spontaneous and stimulus-induced activity with regard to variability, for example, in TTV. That subject is the focus of the next section.
Interaction Model Ib: Nonadditive Rest–Stimulus Interaction and Consciousness
How is the TTV as an index of rest–stimulus interaction related to the level of consciousness? The data by Huang, Dai, et al. (2014) show that stimulus-induced activity as based on decreased neuronal self–nonself differentiation is related to the level of consciousness by showing correlation (see section directly above). This leaves open, however, the role of TTV and thus of the rest–stimulus interaction itself in consciousness.
That interaction was tested for in the subsequent study in VS and anesthesia subjects (Huang et al., 2017b). Applying the same self–nonself paradigm to VS patients, Huang et al. (2017b) demonstrated significantly reduced TTV reduction in PACC and PCC in VS patients that also correlated with their level of consciousness. The VS patients basically did not show any stimulus-induced change in the ongoing variability—there was no TTV reduction after stimulus onset as is typical in healthy subjects. This was further confirmed in a separate sample of anesthetized subjects who, similar to the VS patients, did not show any reduction in their TTV (see Huang et al., 2017b; see also Schurger et al., 2015, for additional support of lacking global TTV reduction in magnetoencephalography (MEG) in VS).
Taken together, these findings suggest that the direct interaction between spontaneous and stimulus-induced activity as indexed by TTV is not only relevant for self–nonself differentiation but, more important, for indexing the level of consciousness. We need to be more precise however. We have already distinguished between additive and nonadditive interaction between spontaneous and stimulus-induced activity (see part II in chapter 2 for details).
Additive interaction means that the stimulus-induced activity simply supervenes on the ongoing spontaneous activity without showing any direct causal interaction. In that case, stimulus-induced activity is just added on top of the ongoing resting-state activity without the latter impacting the former. In contrast, nonadditive interaction refers to direct causal interaction of the former by the latter (see part II in chapter 2 for details). Here, the stimulus-induced activity is not simply added onto the ongoing spontaneous activity; hence the amplitude resulting from a nonadditive interaction is either higher or lower than the sum total of both spontaneous activity and stimulus-related effects (see part II in chapter 2).
As demonstrated by Huang, Zhang, Longtin, et al. (2017) in a separate study, the degree of nonadditive interaction can be indexed by the degree of TTV reduction: the stronger the TTV reduction, the higher the degree of nonadditive interaction. If conversely, there is no TTV reduction, interaction remains rather additive than nonadditive (see part II in chapter 2). That exactly seems to be the case during the loss of consciousness as in VS and anesthesia: both show lack of TTV reduction in PACC and PCC as well as throughout the whole brain (Huang, Zhang, Longtin, et al., 2017; see also Schurger et al., 2015, for additional support coming from magnetoencephalography.
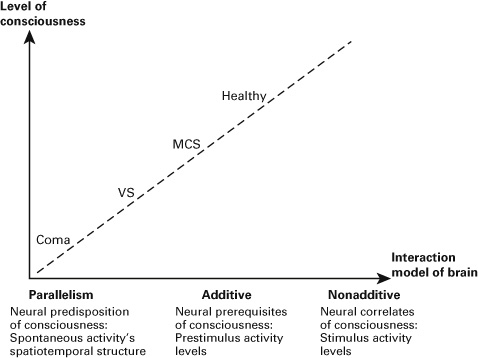
Consequently, I postulate that the degree of nonadditive interaction between spontaneous and stimulus-induced activity (as measured by TTV) can provide a neural signature of the level of consciousness. Nonadditive interaction can thus be conceived as sufficient neural condition of consciousness, that is, as the NCC (Koch et al., 2016). Loss of consciousness, then, may be related to additive rather than nonadditive rest–stimulus interaction as can be seen in the data. Ultimately, both spontaneous and stimulus-induced activity may operate in parallel in patients who have lost their consciousness—nonadditive interaction is then replaced by parallelism as may be the case in coma. This conclusion remains to be tested in the future however (see figure 5.1).
Figure 5.1 Interaction model of brain and the level of consciousness. MCS indicates minimally conscious state; VS, vegetative state.
How sensitive is TTV to different degrees in the level of consciousness? This was explicitly tested by Huang et al. (2017b) in anesthesia when he compared three different levels of consciousness in awake, sedated (e.g., 50% of anesthetic dose), and anesthesia (e.g., 100% of anesthetic dose) states. Again he applied a self–nonself paradigm that elicited strong TTV changes, that is, TTV reduction, in specifically the PCC in the awake state especially during self-related stimuli (and less strong during non-self-related stimuli).
What about the TTV in the sedated and anesthetized state? Most interestingly, lack of TTV reduction in PCC was already observed during the sedated state and did not deteriorate further during the anesthetic state: difference in the degree of TTV was observed in PCC in the sedated state when compared to the awake state. In contrast, the comparison between the sedated and anesthetized states did not show any difference in TTV in PCC any longer. That suggests TTV as an index of nonadditive rest–stimulus interaction, which also seems to be related to the level of consciousness.
Interaction Model IIa: Neural Correlates versus Neural Predispositions of the Level of Consciousness
How are nonadditive rest–stimulus interaction and its relevance for consciousness related to the spontaneous activity? Findings in healthy subjects show direct relation between the spontaneous activity’s temporal structure and the degree of nonadditive interaction (see part II in chapter 2 and Huang, Zhang, Longtin, et al., 2017). How is that related to the level of consciousness? As described in the first part of chapter 4, the spontaneous activity’s temporal structure (as indexed by PLE and neuronal variability) is impaired, that is, reduced during the loss of consciousness.
Huang et al. (2017b) now investigated the PLE and functional connectivity (FC) (i.e., the degree of connection or synchronization between different regions’ neural activities) in the same three stages of anesthesia, awake, sedated, and anesthetic, where he also measured the TTV. Unlike in the TTV, this demonstrated no difference in the spontaneous activity’s PLE and FC during the sedated state when compared to the awake state. In contrast, the findings showed major differences in the spontaneous activity’s global PLE and FC between sedated and anesthetic states (see also Qin et al., in revision; Zhang et al., 2017).
In sum, these findings suggest neuronal dissociation between rest–stimulus interaction and TTV on the one hand and the spontaneous activity’s PLE and FC on the other. During the sedated state and its slightly impaired level of consciousness, the rest–stimulus interaction with TTV is already reduced, whereas the spontaneous activity’s temporal structure (e.g., PLE and FC) is still preserved. The latter only changes once one becomes fully and deeply unconscious as in the anesthetic state. Accordingly, rest–stimulus interaction/TTV and spontaneous activity/PLE–FC seem to take on differential roles for consciousness: rest–stimulus interaction as indexed by TTV seems to be a sensitive and fine-grained marker that is reduced already in sedated states. In contrast, the spontaneous activity’s temporal structure seems to be more robust and immune against such a change—only in the deeply unconscious state as in anesthesia or VS does it change but remains preserved during sedated states.
Taken together, these findings, suggest, albeit tentatively, differential roles of rest–stimulus interaction and spontaneous activity for consciousness. Rest–stimulus interaction as indexed by TTV may be a sufficient neural condition of the level of consciousness and thus be an NCC (Koch, 2004; Koch et al., 2016, for the concept of NCC). Rest–stimulus interaction may thus be central and sufficient for realizing actual consciousness.
In contrast, the spontaneous activity’s spatiotemporal structure (the PLE, neuronal variability, and FC) may be regarded as what recently has been described as NPC (Northoff, 2013, 2014b; Northoff & Heiss, 2015). The concept of NPC refers to those neural conditions that make possible or predispose consciousness while not actually realizing it (as in the case of the NCC). I now propose that the spatiotemporal structure of the spontaneous activity itself predisposes and thus makes possible consciousness. If that very same spatiotemporal structure is altered by itself, consciousness becomes impossible; this is, as suggested by the empirical data, the case in disorders of consciousness (see figure 5.1).
Part II: Capacities and Consciousness
Capacity-Based Approach Ia: Capacities and Causal Structures
What exactly do we mean by the concept of predisposition in NPC? For that, I now turn to a more conceptual discussion about the concept of capacities as introduced and understood by Cartwright (1989, 1997, 2007, 2009); she contrasts capacities and laws that are described briefly in the following paragraphs (without much discussion of philosophical detail).
The traditional view of laws usually focuses on the derivability of theories toward the phenomenon. From this criterion of derivability, observing regularities is essential for the establishment of laws. Regularities of an event can then help scientists to form a hypothesis to explain the causes of the phenomenon. Therefore, laws capable of describing what actually happens are considered fundamental by empiricists. However, Cartwright claims that the capacities of an entity are more basic than laws. Instead of depicting the actual event, capacities indicate the properties of entities that can be fully instantiated without disturbances. Due to the complicated context of the environment, these capacities may not be directly observable in the natural world. Hence, they are abstract and hypothetical in some sense.
Although both regularists and causal structuralists develop their hypotheses from observation, they endow the laws different status. The regularists consider the laws governing the phenomenon: without laws the phenomenon would not exist. But the causal structuralists consider laws the final product of a stable causal structure, being the end point of explanation. The laws are only the attempt of describing the event in an analytic sense, not understanding the essence of the world.
Despite the fact that capacities are not as intuitive as laws during observation, they are not purely logical terms but real causal powers and thus ontological (rather than merely logical) features. Cartwright proposes that these causal powers are the smallest units with which to analyze the phenomenon. They are the subject of the minimal models, and often expressed with the ceteris paribus clause.
When a capacity is put into the context of the environment, it works together with other capacities to form a phenomenon—the phenomenon is the net effect of the causal powers. The combination of different capacities is a composite model. Within it the causal powers form a causal structure. By manipulating each of the capacities in the causal structure, whether removing or adding it to the model, one can understand more about its causal power and its relation with other capacities.
What about causal structure? Causal structure is not only the basis of an event, but a nomological machine to generate laws. The concept of nomological machine regards the law structural and secondary. This contradicts with the regularists’ view, which regards the laws as regular and fundamental. Although one may argue there are regularities in the actual world, they are produced by a stable causal structure, not by themselves as universal laws. These dissimilar views of laws also affect their attitude to their disturbances.
We cannot completely avoid the disturbing factors during experiments in most cases. For the regularists, disturbances are their enemies that interfere with the reproducibility of laws; whereas in the case of a causal structure, disturbances are the factors not yet included in the model. They are part of the environmental background and have their own capacities. Rather than interrupting, they help the researchers to build a more complete causal structure that can better explain a particular phenomenon.
Capacity-Based Approach Ib: Three-Level Model of Nature
In her research, Cartwright comes to a three-level model of nature. There are first the capacities or causal powers. They are the smallest and most basic and fundamental units characterizing nature; we cannot delve any deeper or further in nature. Capacities refer to the most basic existence and reality in nature. Beyond capacities there is nothing that exists and is real in nature. This makes it clear that the concept of capacities is an ontological one that signifies existence and reality in nature, that is, the world.
Such ontological determination of capacities entails that, in our methodological investigation of nature, capacities appear as rather abstract. There is no direct method of observing or measuring them but only an indirect mechanism through the resulting causal structure and the target phenomena. Capacities are consequently the most basic and fundamental units of nature that are revealed once one strips off or subtracts (or abstracts from) all the resulting effects (as manifested in causal structure and target phenomena) of the natural world.
This is reflected in the following quote with regard to capacities in economics: “In methodology, the term of ‘capacities’ is used to mark out the abstract fact of an economic factor: what would be the factor’s effect if it were unimpeded” (Cartwright, 2007, p. 45). Analogously applied to the brain, capacities will mark out a most basic and fundamental factor on which subsequent and directly observable neural activity relies and is based on. We find as we progress in our investigation and discussion that the brain’s spontaneous activity harbors certain capacities that shape and impact subsequent stimulus-induced activity in certain ways rather than in others.
Capacities with their causal power constitute a particular causal structure that results from the interaction between different causal powers (or capacities). This means that due to the capacities different components are arranged in a particular way, thereby forming a causal structure: “A causal structure is a fixed (enough) arrangement of components with stable (enough) capacities that can give rise to the kind of regular behavior that we describe in our scientific laws” (Cartwright, 1989, p. 349; see also Cartwright, 1997). Importantly, without these underlying capacities, such causal structure would not be present at all (or at least would be principally different).
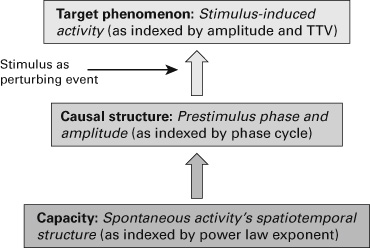
Finally, there is a third level that concerns the target phenomena such as the regular behavior that can be directly observed in empirical–experimental investigation. As indicated in the above quote on causal structure, the target phenomena, such as consciousness for one example, directly result from the causal structure that, by itself, is based on the capacities. How then can we measure and test for whether the target phenomena result from the causal structure? For such determinations, we can use external perturbations such as provoking events to unfold the causal structure and test whether it yields a target phenomenon, for example, consciousness, as in our case (see figure 5.2).
Figure 5.2 Capacity-based model of nonadditive rest–stimulus interaction. TTV indicates trial-to-trial variability.
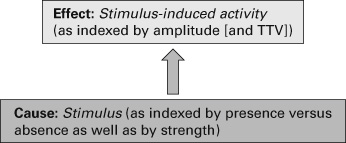
The three-level model distinguishes Cartwright’s capacity-based model of nature from a law-driven model. The law-driven model does not see any need to assume causal structure as a mediating factor: instead there is a direct law-driven relation between the causal powers (that Cartwright attributes to capacities) and the target phenomenon. The causal powers themselves no longer reflect mere capacities but causes, meaning efficient causes that directly (rather than indirectly as in capacities) cause the target phenomenon. The law-driven model thus postulates direct relation between cause and target phenomenon without any mediating causal structure (see figure 5.3).
Figure 5.3 Law-driven model of nonadditive rest–stimulus interaction. TTV indicates trial-to-trial variability.
To understand a capacity-based model of nature and how it distinguishes from a law-driven model, we consequently need to understand how capacities (or causal powers) constitute a causal structure that in turn gives rise to the target phenomenon in question. Importantly, we need to make clear that, without such mediating causal structure, the target phenomenon in question would remain impossible. We are now ready to apply such capacity-based model to the brain and consciousness; this subject constitutes the focus for the remainder of this chapter.
Capacity-Based Approach IIa: Nonadditive Rest–Stimulus Interaction—Causal Structure and Consciousness
How is the example of nonadditive rest–stimulus interaction related to the capacity and the three-level model? The data by He (2013) and especially Huang, Zhang, Longtin, et al. (2017) show that the degree of stimulus-induced amplitude and TTV are dependent on the prestimulus resting-state activity levels in a nonadditive way: lower prestimulus activity levels lead to higher poststimulus-related amplitude and lower TTV. Most importantly, this, as shown by Huang, Zhang, Longtin, et al. (2017), was dependent on the ongoing phase cycle of the resting state at stimulus onset: a negative phase, that is, a trough, led to higher amplitudes and lower TTV than a positive phase, that is, a peak.
The phase and its cycling between positive and negative phases, peaks and troughs, are the means or tools by which the ongoing resting state can impact subsequent stimulus-induced activity: if the stimulus happens to be timed in such a way that it falls into the positive phase, subsequent amplitude will be low and TTV reduction rather high, whereas the opposite pattern occurs when the stimulus timing falls into the negative phase. Finally, the phase and its cycling including the nonadditive interaction itself and its measurement in terms of stimulus-induced activity and TTV were dependent on, that is, they correlated with, the temporal structure of the spontaneous activity itself (as measured by the power law that signifies the relation between lower and higher frequency fluctuations and their power across time) (Huang, Zhang, Longtin, et al., 2017).
How now can we relate these findings to the three-level model of capacities? Let us start from the top, the target phenomenon. The target phenomenon consists here of the observed stimulus-induced activity: this is the neuronal activity we directly observe in response to a stimulus and that we measure in terms of amplitude and TTV. Hence, the amplitude of stimulus-induced activity and its TTV together constitute the target phenomenon.
The target phenomenon is supposed to result from an underlying causal structure that in turn is based on a capacity. Let us start with the causal structure itself. The causal structure consists here of the levels of prestimulus activity. The amplitude at stimulus onset can be either high or low, which directly impacts the target phenomenon, the amplitude and TTV of stimulus induced activity. More importantly, there is the specific phase of the slower ongoing oscillations at stimulus onset (trough or peak) which yield a higher amplitude and stronger TTV reduction when it is a trough compared to when the phase at onset is a peak. Accordingly, prestimulus amplitude and phase and their manifestation at stimulus onset account for the causal structure underlying the target phenomenon.
We demonstrated that the causal structure, as accounted for by prestimulus phase and stimulus-induced amplitude, yields the target phenomenon, the stimulus-induced activity as measured by amplitude and TTV. In contrast, we did not discuss the perturbing event that impinged on and interacted with the causal structure in such a way that it yielded stimulus-induced activity in a nonadditive way. This concerns the external stimulus and its timing with regard to the ongoing phase of resting-state activity, which accounts for the nonadditive nature of rest–stimulus interaction. If in contrast there were no such causal structure with different phase cycles, the interaction would no longer be nonadditive but merely additive.
Imagine if there were to be no phase cycles either positive and negative with low and high excitability, respectively. In that case the causal structure of the prestimulus interaction would no longer yield nonadditive rest–stimulus interaction but merely additive rest–stimulus interaction: the amplitude of the stimulus-induced activity would be merely added on top of the ongoing resting-state activity, and, even more important, the latter’s ongoing variability would probably not be reduced as measured by the TTV. This makes it clear that the underlying causal structure, the prestimulus amplitude and phase cycle, has indeed a direct causal effect on the target phenomenon, the rest–stimulus interaction that consists in its nonadditive nature. Without the causal structure, the prestimulus amplitude and phase cycling, the perturbing event, that is, the stimulus, could only yield additive but no nonadditive rest–stimulus interaction. That, as we have just demonstrated, leads to the absence of consciousness. Hence, the causal structure as second level is central for associating stimulus-induced activity with consciousness.
Capacity-Based Approach IIb: Nonadditive Rest–Stimulus Interaction—Capacities and Consciousness
What about the third level, the capacities underlying the causal structure? The capacity consists in the spontaneous activity itself and more specifically its temporal structure. Huang, Zhang, Longtin, et al. (2017) observed that the temporal structure of the spontaneous activity (as measured in a separate session with the power law exponent [PLE]) that signifies the relation between higher and lower frequency fluctuations and their power across time, specifically, across six minutes as measured) predicted both the prestimulus amplitude and phase cycling and subsequently the degree of nonadditive interaction including the amplitude and TTV of stimulus-induced activity.
How can we account for these findings in terms of capacities? One would assume that the spontaneous activity’s temporal structure, specifically its nestedness/scale-free nature (measured by PLE), provides a capacity for nonadditive rest-stimulus interaction. Specifically, the spontaneous activity’s temporal structure makes possible or predisposes the nonadditive nature of rest–-stimulus interaction. This means that without such temporal structure, as for instance, without lower- and higher-frequency fluctuations being related to each other as measured in PLE, rest–stimulus interaction could possibly not be nonadditive anymore but merely additive.
That seems to be, for instance, the case in disorders of consciousness, which demonstrate the dramatic consequences of a lack of nonadditive interaction, namely the loss of consciousness. Consciousness can thus not only be related to nonadditive rest–stimulus interaction itself but also to its underlying capacities as related to the spontaneous activity’s spatiotemporal structure. We would leave out half of the neuronal mechanisms if we were considering only nonadditive rest–stimulus interaction.
More generally, I here suggest a capacity-based approach to consciousness. This helps us to better understand the distinction between NCC and NPC. The NCC reflect a particular not-yet-fully clear causal structure that allows for nonadditive rest–stimulus interaction and thereby yields the target phenomenon, the association of stimulus-induced activity with consciousness.
In contrast, the NPC concern the capacities, that is, the causal power of the spontaneous activity that first and foremost make possible nonadditive rest–stimulus interaction (and the subsequent association of stimulus-induced activity with consciousness). We saw that that very same capacity, the NPC of the spontaneous activity and its spatiotemporal structure, is lost in disorders of consciousness.
This carries clinical relevance. Changes in the NPC make consciousness impossible so that clinical recovery with return to consciousness is rather unlikely if not impossible—this is, for instance, the case in full-blown coma. Change in NPC must be distinguished from isolated changes in NCC. Isolated changes in NCC still allow for the recovery of consciousness since the underlying NPCs are still preserved. Such isolated change in NCC (and concurrent preservation of NPC) may, for instance, be the case in the minimally conscious state (MCS) and in those VS patients who subsequently wake up (Northoff & Heiss, 2015).
Capacity-Based Approach IIIa: Law-Driven Model of Nonadditive Rest–Stimulus Interaction—Stimulus as Cause
The proponent of a law-driven model of brain may now want to argue that the nonadditive rest–stimulus interaction can be explained in direct causal terms: an underlying cause can directly and sufficiently account for the target phenomenon itself without assuming the mediating effect of a causal structure. Such direct and sufficient cause may be presented in three different scenarios as possible arguments against a capacity-based model of nonadditive rest–stimulus interaction.
The first scenario would be one where everything that I attributed to the resting state could all be related to the stimulus itself with the consecutive assumption of only stimulus-induced activity operating in the brain without any spontaneous activity; the second scenario would acknowledge spontaneous activity but would not attribute any causal relevance to it for rest–stimulus interaction; the third scenario would be one where the cause of nonadditive rest–stimulus interaction would be directly related to the resting state itself, the spontaneous activity.
Let us start with the first scenario. One may assume that the brain itself can only be characterized by stimulus-induced activity while there is no spontaneous activity at all. What is described as spontaneous activity may then be assumed to result from stimuli as well, thus being an instance of stimulus-induced activity rather than reflecting a truly distinct form of neural activity such as spontaneous activity (see, for instance, Morcom & Fletcher, 2007a,b). Although it may seem weird, one could indeed take such a view since the spontaneous activity itself is malleable by external stimuli and thus experience-dependent, as, for instance, reflected in the spontaneous activity’s relation to earlier childhood trauma (see Duncan et al., 2015). However, one can nevertheless distinguish spontaneous and stimulus-induced activity by their different spatiotemporal ranges and scales (see e.g., Klein, 2014, as well as below for details). Let us nevertheless assume for the sake of the argument that there is no distinction between stimulus-induced activity and spontaneous activity so that the brain’s neural operations can exclusively be characterized by stimulus-induced activity.
How would rest–stimulus interaction look in such a case? There would simply be no rest–stimulus interaction since there is no resting state anymore which makes its interaction with the stimulus simply impossible. The observed nonadditive effects could then only be attributed to the stimuli themselves. Different stimuli may elicit different degrees of amplitudes and trial-based variability, for example, TTV, in their stimulus-induced activity. However, due to the absence of the prestimulus amplitude and phase cycle as underlying causal structure, one and the same stimulus should then always elicit the same amplitude and TTV.
This was not the case in our data however, where one and the same stimulus elicited different degrees of amplitude and TTV depending on prestimulus amplitude and phase cycle. The assumption of the law-driven model of directly and causally relating the nonadditive nature of stimulus-induced activity (rather than of rest–stimulus interaction) to the direct causal impact of the stimulus itself is simply empirically implausible and thus argues against a direct and sufficient causal effect of the stimulus itself in a law-driven way.
Moreover, I would argue that, even if the nonadditive nature of stimulus-induced activity is preserved, there would nevertheless be consciousness associated with that stimulus-induced activity. The stimulus must interact with the spontaneous activity and its spatiotemporal structure in order for the nonadditive interaction to be associated with consciousness. Why? Because by interacting nonadditively with the spontaneous activity, the stimulus gets access to the latter’s larger spatiotemporal scale; that, in turn, is important for spatiotemporal expansion and thus for consciousness (see Northoff & Huang, 2017, and part II in chapter 7, for details on the latter point).
What about the second scenario that acknowledges the spontaneous activity but renders it causally impotent? In that case there would be a resting state but its prestimulus amplitude and phase cycles would have no impact on the subsequent processing of the stimulus. The different phase cycles would then no longer reflect different degrees of excitability for the stimulus, which consequently would make impossible the nonadditive nature of the rest–stimulus interaction. Stimulus-induced activity would consequently merely be added on the ongoing resting-state activity with the rest–stimulus interaction being additive rather than nonadditive. This makes impossible associating consciousness to stimulus-induced activity.
The stimulus would still be the sole and sufficient cause for the observed stimulus-induced activity, for example, its amplitude and TTV indicating direct and sufficient causal relation between stimulus and stimulus-induced activity in a law-driven way. That however is not empirically plausible given that the data show clear impact of the prestimulus amplitude and phase cycle on subsequent stimulus-induced activity that is central for consciousness. Accordingly, the second scenario of the law-driven advocate remains empirically implausible as well.
Capacity-Based Approach IIIb: Law-Driven Model of Nonadditive Rest–Stimulus Interaction—“Spatiotemporal Capacities”
What about the third scenario where the resting state is conceived as the sole, direct, and sufficient cause for nonadditive rest–stimulus interaction? In that case one would expect that the degree of nonadditivity would remain always the same during different stimuli, which would then show analogous degrees of amplitudes and TTV. For instance, self- and non-self-related stimuli (as one’s own and other names or auto- and heterobiographical events as phrased in sentences) should then induce the same degree of stimulus-related amplitude and TTV in the same regions.
The last, however, is not the case as a subsequent study by Huang et al. (2016) has demonstrated. He showed that self- and non-self-related stimuli induced different degrees of nonadditive rest–stimulus interaction with subsequently different degrees of amplitudes and TTV’s especially in cortical midline regions (such as the medial prefrontal cortex and posterior cingulate cortex) even though the prestimulus resting state’s distribution of positive and negative, or peak and trough, phase cycles at stimulus onset was the same for self- and non-self-related stimuli.
These data show that the degree of nonadditive interaction is not only upon the resting state itself, that is, its capacity (as reflected in its temporal structure that can be measured with PLE and its ongoing phase cycle at stimulus onset), but also on the content of the stimuli themselves (self- vs. non-self-related, such as one’s own vs. others’ names) and their timing relative to the ongoing phase cycles). The resting state’s temporal structure is consequently not sufficient by itself in a law-driven way as proposed by the advocate of a law-driven model; it is only necessary but not sufficient for nonadditive rest-stimulus interaction.
I therefore propose different roles for resting state activity and stimulus in nonadditive rest–stimulus interaction. The resting state, specifically its temporal structure, provides the capacity for yielding a particular causal structure, that is, its prestimulus amplitude and phase cycle. The resting state’s causal structure has the capacity to interact with different kinds of perturbing events, that is, different stimuli. However, the degree of the actual interaction, that is, the observed degree of nonadditivity, depends not only on the resting state itself but also on the stimulus itself, that is, its content and timing. Together, resting state and stimulus and, more specifically, the degree of their nonadditive interaction yield the target phenomenon, for example, stimulus-induced activity with amplitude and TTV.
What is the role of the stimulus? The stimulus, that is, its content and timing, provides the context for the resting state activity and its capacity—this amounts to what, relying on Cartwright, can be described as “contextual modulation or dependence” (of the resting state’s capacity on the stimulus as context). The assumption of such contextual modulation of the resting state’s capacity by the stimulus is not compatible with and therefore stands opposite to the notion of laws. In the context of a law-driven model, one may want to presume a law that specifically refers to the resting state’s degree of non-additive interaction with stimuli. That law would describe how the resting state itself can mediate different degrees of nonadditive interaction by itself (rather than describing the resting state’s capacity for a certain range of different degrees of nonadditive interaction as in the capacity-based model). Specifically, different degrees in the resting state’s temporal structure would then “translate” one-to-one into different degrees of nonadditive rest–stimulus interaction. Put more philosophically, the resting state itself would exert direct causal impact on rest–stimulus interaction as the resting state itself would then be the sufficient (rather than necessary nonsufficient) condition.
Importantly, such a law describing direct causal impact of the resting state itself on subsequent stimulus-induced activity would exclude possible contextual modulation of the degree of nonadditive rest–stimulus interaction by the stimuli themselves. Such contextual dependence of the resting state’s nonadditive rest–stimulus interaction can only be accounted for by capacities but not by laws. Accordingly, although conceivable on logical grounds, the characterization of the resting state by a law-based model rather than a capacity-based model for nonadditive rest–stimulus interaction must be rebutted on empirical grounds as related to the contextual, stimulus-related dependence of nonadditive rest–stimulus interaction. In sum, the empirical data argue in favor of a capacity-based rather than a law-driven model of brain underlying its propensity for nonadditive rest–stimulus interaction.
Most important, such a capacity-based approach to the brain is central for consciousness. We have seen that the exclusive consideration of stimulus-induced activity as NCC leaves out something crucial, namely the spontaneous activity as neural predisposition of consciousness. The need to consider NPC in addition to NCC urges us to shift from a law-based approach to a capacity-based approach in our explanation of the relation between brain and consciousness. Only when we consider the capacities as described in the NPC, can we understand why and how the brain’s neural activity can be associated with consciousness. The empirical data show that those very same capacities, that is, the NPC, consist in the spontaneous activity’s spatiotemporal structure. Therefore, one may specify the notion of capacities as “spatiotemporal capacities” in the context of brain and consciousness.
Conclusion
I have demonstrated here that the interaction model is not only relevant for the brain (chapter 2) but also for consciousness. There must be nonadditive rather than merely additive interaction between spontaneous activity and stimuli in order for associating stimulus-induced activity with consciousness. Conceptually, the central role of the spontaneous activity is described as “neural predisposition of consciousness.” The NPC concern the necessary condition of possible consciousness as distinguished from the sufficient conditions of actual consciousness, the NCC.
The role of the spontaneous activity of NPC is described further in conceptual terms by the notion of capacities that entail a capacity-based approach. Relying on Cartwright (1989, 1997, 2007, 2009), I attribute such a capacity-based approach to the brain and its relation to consciousness. Specifically, I postulated that the spontaneous activity can be characterized by capacities that predispose consciousness. These capacities seem to exist in the spontaneous activity’s spatiotemporal structure; for that reason, one may specify the brain’s capacities as “spatiotemporal capacities.” I propose that the brain’s spatiotemporal capacities are central for consciousness and further develop and explicate this spatiotemporal model of consciousness in chapters 7 and 8.