
![]() The outer cell wall and the plasma membrane are barriers that regulate what can enter a cell.
The outer cell wall and the plasma membrane are barriers that regulate what can enter a cell.
![]() The cell membrane has tunnels that connect all the cells in a plant.
The cell membrane has tunnels that connect all the cells in a plant.
![]() Aquaporins and transport proteins embedded in the membrane assist water and nutrient ions in entering the cell, while keeping unwanted materials out.
Aquaporins and transport proteins embedded in the membrane assist water and nutrient ions in entering the cell, while keeping unwanted materials out.
![]() The cytoplasm is the liquid part of the cell along with the organelles, the structures in a cell that perform special jobs.
The cytoplasm is the liquid part of the cell along with the organelles, the structures in a cell that perform special jobs.
![]() Mitochondria are the powerhouses of the cell.
Mitochondria are the powerhouses of the cell.
![]() Chloroplasts are the sites of photosynthesis.
Chloroplasts are the sites of photosynthesis.
![]() Protein molecules are constructed on ribosomes located on the endoplasmic reticulum or floating in the cytoplasm.
Protein molecules are constructed on ribosomes located on the endoplasmic reticulum or floating in the cytoplasm.
![]() The central storage facility, the vacuole, is surrounded by another membrane, the tonoplast.
The central storage facility, the vacuole, is surrounded by another membrane, the tonoplast.
![]() Lysosomes and peroxisomes are the recycling centers of the cell.
Lysosomes and peroxisomes are the recycling centers of the cell.
![]() The nucleus is the command center where the DNA is housed.
The nucleus is the command center where the DNA is housed.
![]() Microtubules and actin filaments serve as the communication centers and transportation infrastructures.
Microtubules and actin filaments serve as the communication centers and transportation infrastructures.
THERE ARE LOTS of places we could start. However, I am a big believer in setting up the stage before introducing the players. When it comes to how plants eat, the stage is the basic plant cell. The Cell Theory holds that the cell is the smallest unit of life and that all living things are made up of one or more cells. Moreover, all cells come from pre-existing cells.
Most gardeners studied some cellular biology in school, and many of us have a basic understanding of how cells divide. In recent years, however, scientists have made many new discoveries by studying DNA and gene markers and using bigger and more sophisticated electron microscopes and a whole host of other wonders of modern technology. These discoveries have resulted in a much clearer picture of the plant cell and what goes on outside and inside it.
Plant cells are incomprehensibly small and thus outside the purview of a gardener’s day-to-day thinking. It probably doesn’t help to note that most plant cells are in the range of 10 to 100 micrometers (0.0003937 to 0.003937 inches). Micrometers (or microns) and nanometers (there are 1000 nanometers in a micron) are the measurements used at the cellular level. However, it is helpful to at least try and put the size of a plant cell into some perspective.
First, let’s forget about inches from here on, except to note that 1 meter is about 39 inches. A centimeter is one-hundredth of a meter and getting a lot closer to cell size. It is just smaller than the diameter of an American dime. A millimeter is one-thousandth of a meter and is the thickness of that dime. A micron is one-millionth of a meter and is the thickness of a cell.
Cell components are measured in nanometers, or billionths of a meter. Nanometers are used to measure atoms and molecules. Despite all of these numbers, these small sizes are nearly incomprehensible. Let’s try. A human hair is about 100,000 nanometers wide. You can see one without a microscope and unaided with the naked eye. A water molecule is less than 1 nanometer wide. You can’t see that with a microscope in a high school biology class. Anything smaller than 500 nano-meters can only be seen by using an electron microscope.
Still not getting how ridiculously small this is? If each person on Earth were 1 nanometer tall, we would all fit into a commercial seed packet. That is more than 6.5 billion very tiny people, by the way. Or consider the period at the end of this sentence; it is about 500 to 600 microns in diameter. Five to fifty plant cells would fit into this area. Don’t forget they are three-dimensional.
Another way to appreciate these sizes is to pretend that a single atom is the size of a small pea. A pea pod, then, is the size of a simple molecule. A semi-trailer truck full of pea pods is the size of a complex molecule, and an ocean-going transport tanker that carries a whole bunch of pod-filled semi-trailers is the size of a cell.
Size may seem like a limiting factor when it comes to cells. One might think that for efficiency and economy cells should be large. However, a smaller sphere has relatively more surface area per unit inside area. If a cell were to get too large, there would not be enough membrane surface to supply the cell with sufficient nutrients to survive. In any case, there is a practical limit on the size of cells, and small is not a bad thing.
The number of cells in a plant is equally stunning. A 20-year-old birch tree is made up of something like 15 or 16 trillion cells. Probably more, but who can count that high? Again, these plant cells are not flat objects, as they are so often depicted in school science texts. That tree is decidedly three-dimensional, as are every one of its trillions of component cells. They have thickness and are filled with equally three-dimensional parts composed of three-dimensional atoms, the smallest particle of an element.
Everything from here on should be viewed in three dimensions. By the end of this chapter, you should able to mentally shrink down to a half a micron and wander around a cell, inspecting its parts and getting a molecule’s eye view of things. At that size, our plant cell looks like a factory as large as a football stadium.
Plant cells are surrounded by a cell wall, a strong, lattice-like structure. The cell wall is what distinguishes plant from animal cells, which lack cell walls. Averaging around 20 nanometers thick, its primary function is to contain and protect the cell.
The cell wall is mostly made up of cellulose, which is what makes the fibers of the pages of this book (unless it is electronic). About one-third of all vegetative matter is composed of cellulose, making it the most common biologically produced substance on Earth. Cellulose is a complex molecule made up of many simpler glucose molecules, a type of sugar, that when linked together become polysaccharides (meaning “many sugars”). The structure of polysaccharides allows them to be woven closely to form long strands. These fibrils overlap each other to create layers, which gives plant cell walls great strength. Lest the image of a solid wall confuse things, the cell wall is actually extremely porous. Imagine the cell wall as a luffa sponge.
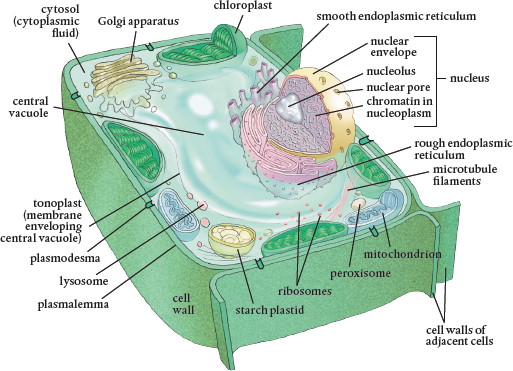
A typical plant cell and its internal organelles. Cells may be small, but they are still three-dimensional and should always be visualized in that form.
Plants are not the only organisms to produce cellulose. Some bacteria are able to make it as well. This fact gave rise to the concept that bacteria were ingested and eventually incorporated into plant cells with great success, for the bacteria and the cells, the result of which is that all plant cells now produce cellulose.
Special protein complexes produce the cellulose fibers of the cell wall. These proteins move along the outer membrane of the cell, which lies just inside the cell wall. These proteins spool out fibrils almost like the spindle on a knitting machine or a cotton candy machine. The supplies that make up the cell wall (including cellulose, hemicelluloses, and pectins) are all produced inside the plant using nutrients (such as boron) that are aggregated there from the soil. The nutrients are transported to the cell membrane for the construction of the cell wall on the other side of the membrane. This is quite an engineering feat, one of many accomplished by these tiny cells.
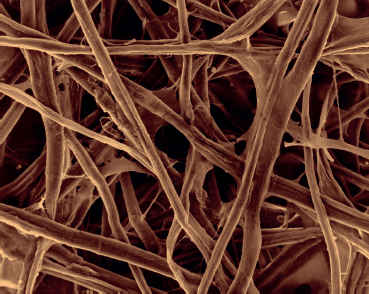
This scanning electron micrograph shows cellulose fibers, which give great strength to the cell wall.
Consider that there are about 3000 molecular units linked together in just one of the polysaccharide chains that form the microfibrils that are woven into cellulose and cell walls. The synthesis of cellulose in the cells requires enzymes, nitrogen-based molecules that vastly speed up the chemical reactions that occur in a cell. If gardeners had to wait for the right sugar molecules to come along and collide with another 3000 times to make just one strand, there is no way we would live to see a plant grow.
Cell walls are not very wide, 0.1 to 10 microns is the norm, but they are very strong and protect the cell. They’re also flexible to a point, just like a reed mat or a wicker chair. Thus, when water flows into a cell, pressure builds inside and the cellulose weave expands a bit. However, it can stretch just so far. This serves as a constraint to the buildup of internal water pressure. When the wall can’t stretch, the inflow of water stops. This is turgidity. The full cell is turgid.
In addition to cellulose, other polysaccharides are produced and utilized in the cell wall. Synthesis of these requires a different set of enzymes. These help bind the cellulose together, forming cross beams, as it were. Pectin, the familiar Jell-O–like substance, is one such polysaccharide. The weaving together and gluing of the cellulose fibrils gives the cell wall great strength. Still, it is porous to small molecules, such as water.
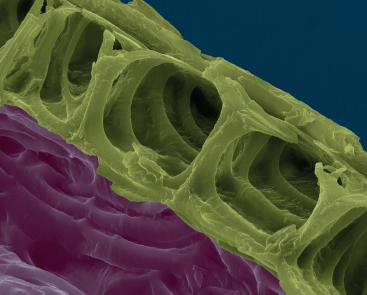
A scanning electron micrograph of cell walls
The polysaccharide fibers that are exposed inside cell walls have negative electrical charges on them, and these attract positively charged particles, known as cations. Some important nutrients held in the soil are positively charged, and they are attracted to the negative charges on the polysaccharide fibers in the cell wall, which adsorb them (that is, hold them on the surface). These nutrients then become available to the plant.
Herbaceous plants just have a thin primary cell wall, whereas woody plants also possess a thicker secondary cell wall that provides additional support. This is a lignin-based layer. Lignin is another long-chained carbon molecule, and it is the major component of wood. A secondary wall sometimes sits up against the primary cell wall and usually contains suberin, a waxy, waterproof substance (again synthesized inside the cell). Cork is essentially suberin.
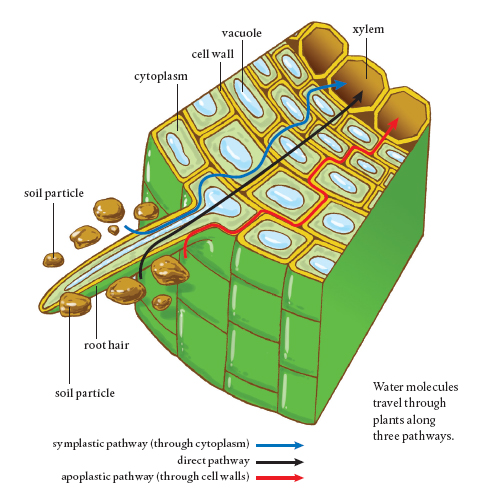
In both herbaceous and woody plants, there is a special set of cells located in the interior of the roots that separates the epidermis and the cortex from the area that protects the root’s vascular tissue. This is known as the endodermis. Cells in the endodermis have walls with a strip of suberin and other waxes, which forms a barrier that prevents water from passing any further through that cell wall. A single layer of these cells encircles the center of the root, and the clogged portions of the cell walls line up to form what is known as the Casparian strip. It prevents water from flowing any further through the root and into the plant. To get there, water has to take a different pathway, one that will regulate what the water can bring into the cells along with it.
Water and gas molecules are small enough to travel around inside the porous cell wall and to flow through it into the adjoining cell walls (until the Casparian strip is encountered). This passageway into a plant through cell walls is called the apoplastic pathway, and it plays a key role in plant nutrition. Water in a cell wall or anywhere along the apoplastic pathway can move through the wall into plants without ever crossing over the cell membrane and entering a plant cell.
In sum, the cell wall is a porous, structural support for the plant cell. It is about 0.2 nanometer thick and sits up against the outer cell membrane. Because it is porous to water and gases, these small molecules can travel into plants by staying within the connected cell walls and not further entering the cells. Once water (and the molecules dissolved in it) encounters the waterproof Casparian strip in roots, however, it either meets the screening criteria of the next layer of the cell, the plasmalemma, and enters the interior of a cell or it travels out of the plant the same way it came in, via the apoplastic pathway.
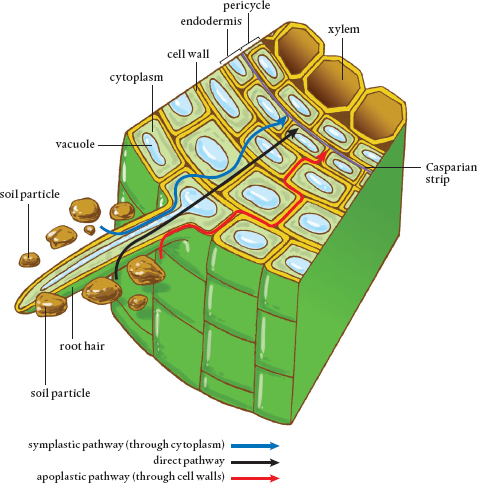
Water can move into a plant through root cell walls via the apoplastic pathway. However, once water encounters the Casparian strip, it must enter the cell across the cell membrane or move out of the root via the Casparian strip.
Water moves through plants along three different pathways. The apoplastic pathway is located in the cell walls, which are porous and connected. Water can travel from wall to wall, but when it encounters the waterproof Casparian strip, it is prevented from crossing into the central area inside the root known as the stele. Water then either must leave the root via the cell wall system or move into a cellular pathway inside the plant.
The symplastic pathway is through the interior parts of cells. Water moves from the cell wall through the adjacent cell membrane and into the cytoplasm of a plant cell. Each cell of a plant is connected to its neighbors by tiny tunnels called plasmodesmata. The symplastic pathway is entirely inside plant cells, with molecules able to move from one cell to the next and then the next, throughout the plant. Cell organelles in the cytoplasm are not part of the symplastic pathway. They get in the way, and molecules must move around them if they are to stay on the symplastic path.
Finally, there is the intercellular or direct pathway. The chemical properties of water molecules and their small size allow them to move directly through cell walls and cell and organelle membranes, regardless of whether the structure acts as a barrier to other molecules. Thus, water can take the short route to the plant’s vascular system, which is where all three pathways are headed.
Water is essential for the movement of nutrients through the plant. It is important to note that water molecules travel along all three pathways simultaneously, which helps to explain the tremendous volume of water that travels through a plant.
The plasmalemma is the outer cell membrane that sits against the inside of the cell wall. A permeable membrane allows things to pass through. Because only certain things can pass through the plasmalemma, however, it is a semi-permeable membrane, screening what can enter and exit a cell.
The plasmalemma is a barrier holding in the liquid cytoplasm and organelles that make up a cell. The plasmalemma holds all of this inside, while keeping some other things from entering the cell. This is not just about getting nutrients in to the plant. The flow through the plasmalemma is bidirectional. How else would plant-produced exudates move out into the soil?
Plasma is matter that is not quite solid or liquid or gaseous, but rather in a phase that is something of all three. Many of you may have a plasma television set. Try moving your fingers around the screen when the set is on to see how plasma operates. Plasma fills fluorescent lamps and neon signs when electricity flows through them. Lightning is plasma, as are flames and the aurora borealis displays (hey, I am an Alaskan). The point is that the plasmalemma is a not a solid, rubber-like barrier. Rather, it is a semi-fluid barrier, much like a thick oil or half-jelled gelatin. Its constituent molecules are moving around and shifting places. It is essentially made up of two sheets of phospholipids, phosphorus-based molecules, punctuated with proteins and carbohydrates that have special functions.
Phospholipids are two-part molecules. The head is a negatively charged phosphate ion that is water-soluble. This means the heads dissolve in water. On the other end, the lipid tail is a carbon-based chain that lacks an electrical charge. It is not soluble in water. In fact, this end of the phospholipid is hydrophobic: it lumps together with other hydrophobic molecules rather than dissolving in water. This is exactly what happens to the fat in chicken stock or oil poured into water. It lumps together, rather than dissolving.
The plasmalemma consists of a double sheet of phospholipid molecules arranged so the water-loving heads point outward to the aqueous extracellular environment and inward to the aqueous cytoplasm. The two sets of hydrophobic tails form an inner, middle layer that is not water-soluble. Imagine two pieces of bread each slathered with peanut butter, made into a sandwich. The peanut butter is the hydrophobic lipid tail layer and each piece of bread is a sheet of the water-soluble heads.
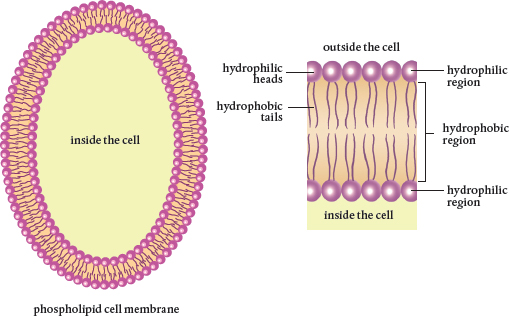
Phospholipid molecules that make up the double-layered cell membrane have phosphate heads and lipid tails. They form a barrier that protects the cell.
Certain molecules can easily cross through the plasmalemma. These are either small enough to fit between the phospholipid molecules or to dissolve in water and move through it in dissolved form. Molecules of oxygen, carbon dioxide, and nitrous oxide pass through the plasmalemma by diffusion, the natural movement of molecules to areas of lower concentration. Water molecules pass through the plasmalemma via osmosis, which is simply the diffusion of water.
How do other molecules get into or out of a plant cell if the plasmalemma is such an effective barrier? The answer lies in special proteins and carbohydrates embedded in and transversing the plasmalemma. These afford different kinds of molecules passage from the apoplastic pathway to the symplastic one, through the plasmalemma. They are key to how nutrients enter and wastes and exudates leave plant cells.
 Plasmodesmata Tunnels Plant cell walls are all connected via the apoplastic pathway. The area inside a cell membrane is connected to the inside of adjacent cells, forming the symplastic pathway, which is made possible by the presence of tunnels, plasmodesmata (or plasmodesma, when referring to just one). Because each and every cell of a plant is connected, if you were small enough you could start out in a root cell and eventually find yourself on the top of a leaf.
Plasmodesmata Tunnels Plant cell walls are all connected via the apoplastic pathway. The area inside a cell membrane is connected to the inside of adjacent cells, forming the symplastic pathway, which is made possible by the presence of tunnels, plasmodesmata (or plasmodesma, when referring to just one). Because each and every cell of a plant is connected, if you were small enough you could start out in a root cell and eventually find yourself on the top of a leaf.
During cell division, bits of the plasmalemma get trapped between the two new cells and form the tunnels that connect the two. If you visualize a cell as one of those long balloons that can be twisted into an animal, you will note that bits of the balloon get twisted whenever another section is created. The balloon is like a mother cell’s plasmalemma, and the twist becomes the connection between the two new cells. This happens every time a cell divides, such that all of the cells in a plant are connected internally. Therefore, it is possible for water and other molecules of the right size to travel from one cell to the next through these tunnels via the symplastic pathway. Because all the cells in a plant are internally connected by plasmodesmata, it is important that the plasmalemma screen out what should not be traveling into the symplastic pathway. Once a substance gets into one cell, there is a way to get to all other cells without being so regulated.
The analogy to the balloon twisting breaks down when you learn that an individual plant cell can have from 1000 to 100,000 plasmodesmata. These tunnels are 50 nanometers in diameter and up to 90 to 100 nanometers long. (Remember, a water molecule is about 1 nanometer in diameter.) Each has a smaller, tubular connection to the endoplasmic reticulum, a cellular organelle that serves as the pathway for transport of cellular materials inside a cell.
The size of a plasmodesma limits the size of the molecules that can pass through cells. At each end of these interconnecting tunnels (and sometimes inside them) are proteins that act like sphincters. These are responsible for regulating what molecules are allowed through. Again, there can be 100,000 of these little machines in a single cell membrane connecting it to other cells.
As for the differing numbers of plasmodesmata per cell, their numbers depend on the function of the neighboring cells. If there is a need for a lot of transport between cells, there can be as many as 15 plasmodesmata per square micron. (Many cells only have one plasmodesmata per square micron.) That’s a cellular screen window, for sure.
Aquaporins Water molecules are small enough that they can sneak through those lipid tails in the phospholipid membranes by osmosis, the movement of water molecules to where there is a lower concentration. When scientists measured the flow and amounts of water into and through cells, however, they realized that something other than simple water movement was at play. Aquaporins are special proteins embedded in the plasmalemma of plant cells that transport water molecules—and only water molecules—across the phospholipid membrane.
Aquaporins cover as much as 10 percent of the plasmalemma surface. Given the need for water in plants, this high proportion of surface area makes sense. As one might begin to suspect, plant cells involved in water absorption have more aquaporins than those that aren’t. (Plants develop with amazing specificity that often passes by those who only observe the flowers.)
Aquaporins employ an electrical field to transport water from outside the cell through the plasmalemma, one water molecule at a time. Each water molecule has a positive end and a negative end, and the molecules can link up in chains. This polarity works with the electrical charge of the aquaporin protein. (Beam me up, Scotty!) Among other functions, aquaporins work with the plasmalemma and plasmodesmata to control the turgidity of the cell.
Transport Proteins Play a Crucial Role Unlike plasmodesmata, which are actually part of the membrane, aquaporins are distinct and separate proteins embedded in the plant cell membrane. However, aquaporins are so strongly embedded in the membrane that it is extremely difficult to separate them from it. As such, they move with the plasmalemma. (It’s plasma, remember? Imagine colors moving on a soap bubble.)
Aside from aquaporin, there are many kinds of transport proteins that are embedded in the plasmalemma. Each allows transit through the membrane of specific molecules. Together, these are known as integral membrane proteins (IMPs). What once appeared to be a smooth membrane has been discovered to resemble a rock climbing wall at the gym, punctuated with hand and foot holds. These IMPs are the entryways for plant nutrients.

This atomic force micrograph shows the phospholipid bilayer plasma membrane and its proteins (spikes). The proteins either span the whole membrane (transmembrane proteins) or sit on its surface (peripheral proteins).
A tremendous amount of a cell’s energy and the DNA blueprints it carries is devoted to making these transport proteins. Without each of them, a plant cannot take in the necessary building blocks for what it produces, and a cell makes everything it needs. That means there are a lot of different IMPs, as well as DNA to regulate their production and replacement. However, IMPs are key to any new resource intake, so the expenditure is well worth it—vital, in fact.
Integral membrane proteins can be grouped into three broad categories depending on how they transport specific molecules. The first group, channel proteins, serves as tunnels. Mechanisms at either end of the protein act as gates and regulate the flow of molecules by changing size or shape, thus giving rise to the name gated channels. Transit through channels does not require the addition of energy. It all depends on the natural propensity of molecules to move from areas of higher concentration to those of lower concentration, as well as the attraction between oppositely charged molecules. Because no external energy is required, this is known as passive transport. Don’t confuse this with slow transport. Water and other specific ions pass through channel proteins, one molecule at a time, at the incredibly high speed of 10 million molecules per second.
The next group of IMPs consists of pump proteins. As the name suggests, these serve as miniature pumps. The most prevalent kind pumps positively charged hydrogen ions (H+) out of the membrane, and these accumulate on the outer surface. Other molecules located there can use the energy of these hydrogen ions to move into the cell. There are sodium and potassium pumps. Because energy is used to run these pumps, their process is known as active transport.
Cotransporters or carrier proteins represent the third group of IMPs. These proteins bind to molecules, causing changes in the protein shape that move the bound molecules across the membrane. This is called facilitative transport because the transported molecule is helped or facilitated across the membrane. This is a form of passive transport because it does not require the input of energy.
Receptor proteins in the membrane transport signals across cell membranes. These membrane proteins cause chemical changes inside the cell as a response to extracellular stimuli. They act in concert with carbohydrate signaling mechanisms also found in the plasmalemma, forming infrastructure that connects all cells to one another and to the external environment. In many instances, receptor proteins create the chemical conditions inside or outside the cell that cause nutrients to pass into plants and they direct the nutrients’ intracellular path.
Integral membrane proteins make up 50 to 75 percent of the surface of the plasmalemma. Remember, this membrane is already covered with plasmodesmata, as much as 10 percent of its area. These proteins provide part of the answer as to how nutrients get into plants and exudates and metabolites flow out. To move through an IMP, a molecule must be charged—that is, it must be an ion. Thus, plant nutrients are almost always in ionic form. The exception is boron (and organic molecules that apparently can be taken up by a small group of tundra plants).
Sometimes, there are materials that need to be moved through a cell membrane for which there isn’t a specific transporting protein. Perhaps the molecule is too large. Endocytosis is the process whereby a membrane envelops something outside the cell, pinches off, and brings it into the cell. Conversely, exocytosis occurs when a membrane opens to the outside and dumps material out of the cell. Organelles inside plant cells and vesicles (something like a bubble) have membranes of the same material as the plasmalemma, enabling them to fuse to it. The material carried in a vesicle can be dumped outside of the cell by exocytosis without having to go through an IMP.
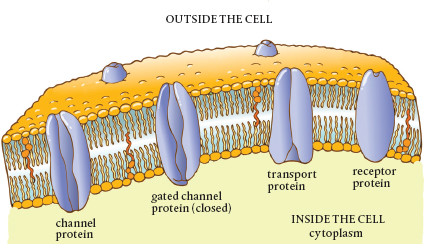
Cross section of the plasmalemma showing various types of integral membrane proteins
The Importance of the Plasmalemma The plasmalemma is the regulator of what comes into and exits a plant cell. It enables the cell to keep a balance between the outside and inside, pumping water and ions in and out to ensure the proper conditions for life. The plasmalemma is the source of communication and signals between the cell and the outside world as well as with other cells. It helps to maintain and adjust cellular turgidity by regulating the amount of water held inside the cell. Finally, the plasmalemma is the site where the machines that build the cell wall operate.
The cytoplasm is all of the stuff inside the cell membrane, except for the nucleus. The purpose of the membrane is to hold the cytoplasm in and keep unwanted things out. Cytosol, a clear, jelly-like fluid, is by far the major component of cytoplasm. It holds the cell’s organelles, the sites of specific biochemical functions within the cell. The cytosol also holds vesicles that serve as tiny cellular warehouses. Some of these store building blocks such as carbon, oxygen, and proteins. Others act as cellular trash-cans containing wastes and sequestering toxins until they can be disposed of outside of the cell, mostly by exocytosis.
Mitochondria are organelles that produce energy for the cell from the sugars made during photosynthesis. Because cells require so much energy, there are hundreds and even tens of thousands of mitochondria in each plant cell. The mitochondrion is the site of cellular respiration, the breaking down of sugar using oxygen. The energy formed in the process is stored in adenosine triphosphate (ATP) molecules, the universal currency for cellular energy.
Mitochondria are bound by their own double phospholipid membranes, complete with embedded transport proteins. However, the mitochondrion’s inner membrane is folded over and over like ribbon candy, producing numerous attached compartments, or cristae. This folding provides more surface area so more energy-producing chemical reactions can take place.
Mitochondria are miniature generators, the powerhouses of the cell. They are the sites of respiration, the process that produces energy from the sugars made elsewhere. A typical plant cell can have tens of thousands of these cellular generators. The number appears to depend on the cell’s function in the plant.
The cylindrical mitochondrion has a double-layered phospholipid membrane, complete with embedded transport proteins like the plasmalemma of the plant cell. The big difference is that the inner membrane is folded to increase the surface area and to create compartments, much like ribbon candy. These compartments are cristae, the sites where the necessary chemical reactions take place. The energy created by these biological generators is stored in the phosphate bonds of ATP molecules.
Mitochondria not only have a double phospholipid membrane typical of plant and animal cells, but they also have their own DNA and divide just like bacteria. In fact, it appears that these organelles once were freeliving bacteria that entered into such successful symbiotic or parasitic relationships with plant cells that they were finally biologically assimilated into them.
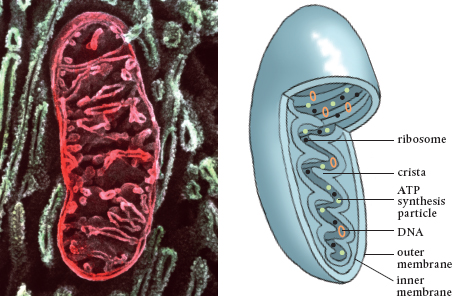
This colored scanning electron micrograph and accompanying diagram each show a single mitochondrion.
Inside the cristae, enzymes strip electrons from glucose molecules and use them to add phosphorus atoms to adenosine diphosphate (ADP) molecules, making ATP molecules. The energy in the new phosphorus bond is what makes ATP so valuable. Breaking this bond releases the energy. As you might expect, cells that require more energy have more mitochondria to produce more ATP molecules.
Producing energy is not the only role of mitochondria, however, They also synthesize the phospholipids that are so necessary for cellular membranes. Mitochondria also help to store calcium, which is used in intercellular signaling and directing traffic in the symplastic pathway.
To the studied eye, mitochondria look a lot like bacteria. They carry their own set of DNA, which is similar to that of bacteria. They divide like bacteria and have a double membrane, which is just like that of bacteria. Scientists now believe that mitochondria were once free-living bacteria that somehow got into the plant cell. There is an argument as to whether these bacteria entered into a symbiotic relationship with the plant cell after being ingested by endocytosis, or whether the remnant DNA that shows the ancestral bacteria had a flagellum (a moving tail that provides locomotion to certain bacteria) means the early relationship was more parasitic.
In either case, there is little argument that eons of assimilation have made mitochondria an integral part of plant cells. Without them, there simply wouldn’t be enough energy to get things done and no plasma membranes to hold, protect, and regulate cellular content. Fortunately, you can find them all over the cellular factory, generating power for plant life (and generating phospholipids in the off hours). Mitochondria account for up to 20 percent of a cell’s total volume. Life, no matter how tiny, runs on energy.
Plastids are miniature factories and storage facilities that are found throughout the plant cell. There are three groups of these organelles: chloroplasts, leucoplasts, and chromoplasts. Chloroplasts are the sites where photosynthesis takes place. They are filled with chlorophyll, a green pigment that absorbs energy from sunlight. The light excites electrons, and this energy is captured. Chlorophyll gives plants their green color. Leucoplasts are colorless (or white) and are found in parts of the plant not exposed to light, such as the roots and seeds. They are where starch grains, lipids, and proteins are formed and stored. Leucoplasts are also found in the white or colorless parts of variegated leaves, suggesting that they may be mutated chloroplasts. Amazingly, some leucoplasts can convert into chloroplasts when exposed to light. Chromoplasts contain the red, yellow, and orange pigments found in flowers, fruits, and some roots. These colors are useful in attracting pollinators and seed dispersers.
Chloroplasts are the most familiar of the plastids and are shaped like tiny lenses (2 to 8 microns long). An average plant cell contains 10 to 100 of them. The average leaf (if there is such a thing) would have a whopping 500,000 chloroplasts per square millimeter. Like mitochondria, chloroplasts have a double phospholipid membrane. The inside membrane is also folded to add a lot of surface area for the photosynthetic reactions to take place. The concept of a conveyor belt comes to mind.
Inside each chloroplast are thylakoids, membrane-bound structures that are flattened, hollow discs, stacked one on top of another. Attached to these discs are green, chlorophyll-filled antennae that absorb light. When light hits the chlorophyll molecules’ electrons, they jump to an outer orbit (I will discuss electron orbits in the next chapter), which increases their energy level. The souped-up electrons flow like electricity through the thylakoid and accumulate in one membrane compartment.
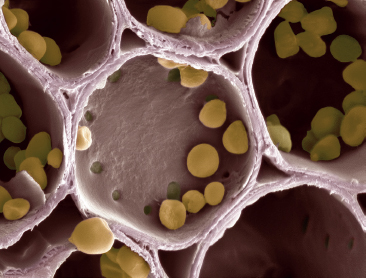
This scanning electron micrograph shows starch grains within amyloplasts in the cell wall compartments of potato tuber cells. Starch is synthesized from glucose, a sugar formed in the leaves during photosynthesis and transported to the tuber.
When the concentration of these electrons inside the thylakoid compartment becomes greater than outside, the laws of thermodynamics take over and they fight to move out and away from each other. They can do this only by passing though a special channel protein in the membrane, the enzyme ATPase. The movement of electrons through the channel causes phosphates to be added to ADP molecules, creating ATP, which is the currency of energy in biological systems. Removing the phosphate yields ADP again, plus the release of energy.
The electrons then get a second dose of light and are picked up by nicotinamide adenine dinucleotide phosphate (mercifully abbreviated as NADP). The ATP created in the first go around is used to form sugar molecules. Each chloroplast can create thousands of these every second, provided there is light to excite electrons in the chloroplast. Remember, there are half a million chloroplasts in a square millimeter of leaf. That is quite a sugar factory.
There is a close similarity between chloroplasts and cyanobacteria, chlorophyll-filled photosynthesizing bacteria. Both have a double membrane and DNA and both divide. Again, this similarity suggests that some ancient, free-living ancestors were ingested into plant cells or entered as parasites. It is evident that the cell incorporated parts of the cyanobacterial DNA into its own, and chloroplasts are now an integral part of plant cells.
Ribosomes are bead-shaped organelles that serve as the sites of protein synthesis, which requires much of the energy produced by a cell. They are the smallest organelles in a plant cell (25 to 30 nanometers). Ribosomes can be attached to the membranes of the rough endoplasmic reticulum (another organelle described later in this chapter) or floating free in the cytosol. Although some cells have thousands of ribosomes, most usually only contain about a thousand. Perhaps it is due to their size that ribosomes were not discovered until 1953.
Ribosomes are constructed in the nucleus and transported out into the cytoplasm. Ribosomes are interchangeable, and their numbers fluctuate depending on the need for proteins. The free-floating ribosomes make proteins for use in the cell, whereas those on the endoplasmic reticulum synthesize proteins that are exported out of the cell for use elsewhere in the plant. There are no lipids in ribosomes, no membranes. Ribosomes also carry a negative charge.
There are typically forty to fifty chloroplasts in most plant cells. Here, each chloroplast is shown as if cut lengthwise. Each is surrounded by an external double membrane, which provides protection for the operations inside. The area surrounded by the inner layer is known as the stroma. This is where light reactions occur during photosynthesis. Located here are the grana (yellow), stacks of flattened thylakoids. Only grana contain the chlorophyll pigments that give plants their green coloration and convert sunlight to energy. Light energy absorbed by the chlorophyll in the chloroplast is used to convert carbon dioxide and water into sugars and starches. The large amounts of starch produced during photosynthesis appear as dark circles within each chloroplast.
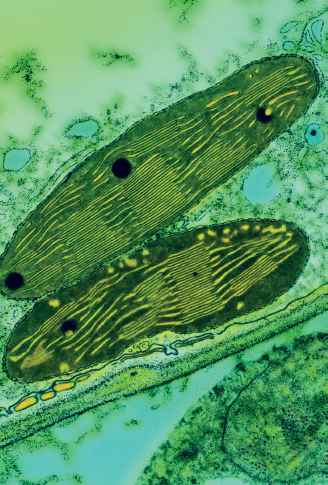
This colored transmission electron micrograph shows two chloroplasts in the leaf of a pea plant.
Imagine the ribosome as a very busy knitting machine, one that you pull balls of wool through and out comes a woven strand. Into the ribosome go strings of amino acids, the building blocks of proteins, and out they come connected in just the right order to make a specific protein. A ribosome can make various kinds of proteins depending on the chemical signals it receives. The particular combination and order of the amino acids (there are twenty) determines the characteristics of the protein.
Deoxyribonucleic acid (DNA) and ribonucleic acids (RNAs) are the molecules involved in replicating the genetic code used to build everything in a cell. The instructions for the particular protein are delivered by messenger RNA (mRNA), and the sequence is copied in the ribosome. The amino acids that are made into proteins are brought to the ribosome by transfer RNA (tRNA). A well-fed plant will have many times more tRNA than mRNA, so there is never a need to wait for supplies to construct a protein.
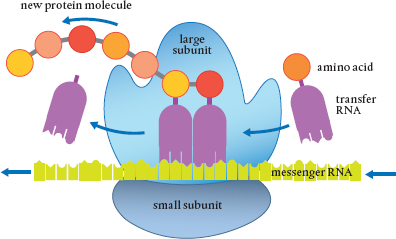
Ribosomes are the sites where messenger RNA sequences (copied from DNA in the nucleus) are read and proteins are synthesized from combinations of the twenty amino acids. The amino acids are brought to the ribosome by transfer RNA.
Don’t be fooled by the tiny size. Each ribosome is a real assembly line, precise in its actions, with all the equipment necessary to connect amino acids in the proper order to synthesize proteins. They are small, but the synthesis of proteins uses much of the energy produced by the cell. In any case, some of these proteins are used inside the cell, and others are exported to different parts of the plant. Once a protein chain is created in the ribosome, it is released and transported to the Golgi apparatus, another organelle inside the plant cell, for final processing before being taken to the cell membrane or used inside the cell.
The endoplasmic reticulum is a long membrane folded over itself again and again to create the lumen, a maze of cavities. Many ribosomes are attached to a section of these folds, making it look bumpy, hence it is called the rough endoplasmic reticulum.
The number of ribosomes thin out along the smooth endoplasmic reticulum, and it is from here that the proteins produced in the ribosomes are shipped to other parts of the cell. Nucleotides are added to the ends of the chains to serve as the address for delivery. The smooth endoplasmic reticulum is also the site where proteins have sugars added to them to become glycoproteins needed for certain cellular functions and reactions. In addition, lipids are made in the smooth endoplasmic reticulum. Another important function of this organelle is changing toxins to more benign substances so they can then be safely transported out of the cell. Vesicles form by endocytosis around waste toxins here, and vesicles also transport proteins from the endoplasmic reticulum for further processing.
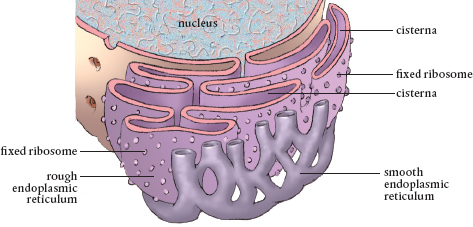
The endoplasmic reticulum extends from the nucleus to the cell membrane.
The endoplasmic reticulum extends from the nucleus throughout the cell to the plasmalemma. The extensive network of the endoplasmic reticulum allows molecules to travel an easier path than having to slog through the jelly-like cytosol and dodging all manner of intercellular molecular traffic of synthesized and synthesizing molecules.
The endoplasmic reticulum is connected to the Golgi apparatus (or Golgi complex), membrane-enclosed organelles named after an Italian histologist, Camillo Golgi. (I can’t think of anyone else whose name graces a cell organelle.) These are the packaging and shipping centers in a cell. Each Golgi apparatus is surrounded by tiny tubules that start at the endoplasmic reticulum and radiate throughout the cell. These and less visible axial fibers are used as tracks along which loaded vesicles move. The vesicle fuses with the membrane of the Golgi apparatus and deposits its contents into the assembly line, where they are modified.
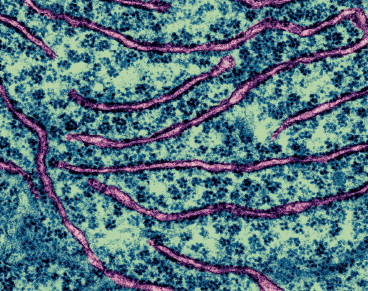
Golgi apparatus in a plant cell. Stacks of cisternae and vesicles are present.
The Golgi apparatus is really a number of Golgi bodies, stacks of membrane-enclosed discs. These contain enzymes that modify the materials sent to the complex. Once finished, the Golgi apparatus ensures these materials reach their ultimate destination, which can be inside or outside the cell. What kind of materials? Proteins are finished in the Golgi apparatus, and the polysaccharides (long sugar chains) needed for making cellulose and used in root exudates are assembled there as well.
There are five to eight Golgi apparatuses in a plant cell. This is not a lot when you realize that almost all the molecules in a cell pass through one of them. Each Golgi body within a complex contains separate sets of enzymes. Inside the Golgi bodies are long passages where molecules can be finished, sorted, chemically tagged with a destination code, and loaded into vesicles for final transport. The cisternae in the sacs contain lots of tiny transport vesicles to which specific proteins and polysaccharides bind for the transport out. Each vesicle’s phospholipid membrane merges with the cell membrane and the contents are emptied. Those vesicles containing waste material fuse with the vacuole.
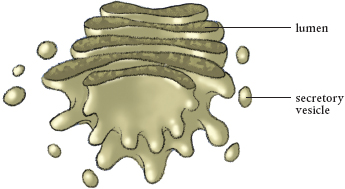
The Golgi apparatus is where the final packaging is done, and then molecules are shipped out in vesicles.
The vacuole is a single-membrane organelle filled with cell sap, a watery mixture of molecular compounds. This sap helps maintain pressure (turgor) in the cell. Cell sap mostly contains water, as well as sugars, enzymes, proteins, organic acids, and the pigment molecules responsible for the red, blue, or purple of flowers and the red of leaves. It also contains cellular wastes. It is slightly acidic, meaning it has a pH below 7.
Young plant cells may have several small vacuoles. These individual vesicles have the same kind of plasma membrane, so they can merge, just like smaller soap bubbles merge to become larger ones. Mature cells have only one large vacuole. The vacuole’s membrane is the tonoplast. Like the plasmalemma, the tonoplast is embedded with specialized transport proteins that allow materials to move across the membrane without having to deal with the barriers created by the phospholipid molecules. Transport of materials across the tonoplast most often requires the input of energy.
Each plant cell usually has one main vacuole, which can make up as much as 90 percent of the cell’s volume. The vacuole fills and increases the turgor, or pressure, within the cell. A full vacuole is one reason plant stems can support heavy fruits and store water. When the vacuole shrinks enough due to a shortage of water, the plasmalemma pulls away from the cell wall. Collectively, this causes plasmolysis or wilting.
The tonoplast helps keep the pH in the cytosol at the proper level. The pH is a measure of the concentration of hydrogen ions (H+) in a solution: the more of these, the lower the pH and the more acidic the solution. Some of the transport proteins in the tonoplast membrane can pump hydrogen ions into the vacuole, thus increasing their concentration and lowering the pH. The concentration of hydrogen ions is carefully regulated to keep the pH in the cytosol around 7, which is neutral.
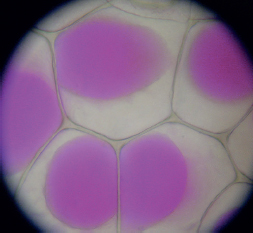
Epidermis cells after plasmolysis. The vacuoles (stained pink) have shrunk.
As for the wastes collected and deposited into the vacuole, in some instances part of the tonoplast merges with the plasmalemma to export wastes from the cell, or wastes are moved to the lysosomes and peroxisomes. Cell death is usually a result of the tonoplast no longer working. When it fails, wastes are no longer contained, turgor is affected, and the pH of the cytosol is altered.
The vacuole membrane can also merge with lysosomes and peroxisomes. Lysosomes are the cell’s recycling centers. They are formed when part of the Golgi apparatus membrane buds off. Lysosomes contain enzymes that digest large molecules (mostly proteins) into their components, which are then recycled in the cell. The number of these digesting vessels varies depending on the cell function, but can range up to 100 or so per cell.
The enzymes in lysosomes require an acidic pH around 5 to do their job. If the pH is any higher, they won’t work. The pH of the cytosol needs to be close to neutral so the other organelles and their products are not harmed. This means a lot of hydrogen ions must be pumped into lysosomes to keep the pH low enough for their enzymes to function. Should some of the enzymes leak out of the lysosome, the higher pH they encounter prevents them from functioning and digesting organelle membranes and materials needed for synthesis. At the end of a cell’s life, a signal is sent to lysosomes to release their enzymes to digest the dead cell. Apparently, without cellular function, the environment becomes acidic enough so the enzymes can work.
Lysosomes only have a single-layer membrane, which means they can more easily merge with the membranes surrounding vesicles and vacuoles, enabling them to receive waste that is then taken away and broken down. This waste stream includes bacteria, nutrient molecules, proteins, lipids, polysaccharides, and nucleic acids. It can even consist of organelles that are no longer functioning, such as a ribosome or spent mitochondrion. All of the material is recycled.
Peroxisomes also serve as digestion vessels in the cytosol. These are generated from the proteins and lipids made in the endoplasmic reticulum. Unlike lysosomes, the enzymes in peroxisomes digest lipids and fats. In seeds, for example, they supply the enzymes that start the conversion of stored fatty acids into sugars. Peroxisomes also help in the cellular assimilation of nitrogen and the metabolism of hormones.
The nucleus is the only organelle that is not considered part of the cytoplasm. It is here that a plant cell’s hereditary material, its genome, is stored along with instructions on how to synthesize proteins. The nucleus is the coordination center for cellular activity, and it accounts for about 10 percent of the space in a cell.
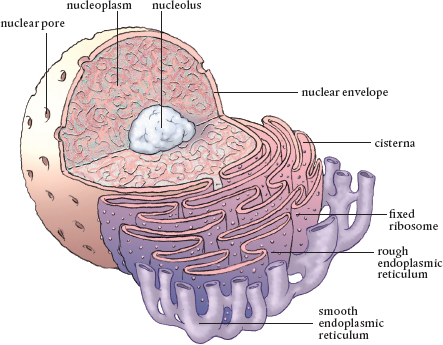
Arguably the most important organelle, the nucleus is where DNA is stored and protected. Note the connection to the endoplasmic reticulum and the nucleolus, where ribosomes and RNA are assembled.
The nucleus is surrounded by two phospholipid membranes that are separated by a space known as the nuclear envelope, which surrounds the entire nucleus. The outer nuclear membrane is continuous with the membrane of the rough endoplasmic reticulum and is also studded with ribosomes. These membranes keep the cellular cytoplasm separated from the stuff inside the nucleus, the nucleoplasm or nuclear sap that includes DNA and RNA.
There are holes in the nuclear envelope that cover up to 15 percent of its surface. These nuclear pores range in diameter from 10 to 100 nanometers. They allow the nucleus to communicate with the cytosol while screening and regulating the molecules trying to transit the nuclear membrane in either direction. Small building-block molecules have no problem moving through. However, large proteins and RNA have to be selected to move through these pores. Obviously, there is an important need for control of what gets in and what is let out of the nucleus. After all, it holds the DNA of the cell and can’t be contaminated.
The nuclear lamina, the inner membrane of the nuclear envelope, contains a mesh network of fibrous proteins that anchor the nucleus in the cell and give it some rigidity. Inside the nucleus are the building blocks, the cell’s precious DNA and RNA, and the nucleus’s own organelle, the nucleolus. This is where the units that make up ribosomes are produced, along with the RNA molecules used for duplication of the DNA and transport of amino acids during protein synthesis. Both are transported out of the nucleus to the endoplasmic reticulum and moved on to the Golgi apparatus. Most plant cell nuclei have more than one nucleoli.
Most plant cell organelles are connected to or in extremely close proximity to a network of transport tubules known as microtubules and actin filaments. These are 2000 times thinner than a human hair. Together actin filaments and microtubules provide structural support for the cell and serve as its transportation infrastructure and communication network. Molecules move along the microtubules and filaments as if they were railroad tracks. Signals can be sent through them and along them as well, so they serve as the intracellular Internet or telegraph system.
It may seem duplicative to have two kinds of organelles perform essentially the same functions. However, when it comes to being able to send signals and deliver the building blocks, finished products, and organelles necessary for survival, redundancy makes a lot of evolutionary sense. In addition, the microtubules and actin filaments work together, so there is synergism between the two systems.
It does take imagination to view plant cells in your mind. How do you get your mind around the fact that there are about 10,000 different kinds of proteins moving around in each microscopic cell, with as many as 10 billion individual ones in total? Then there are the precursors to proteins as well as almost countless numbers of other molecules made by the cell. The number of molecules inside a plant cell, frankly, is incomprehensible, and moving through them must be like swimming in a stew.
Most of us will never have access to the kind of equipment it takes to see the molecules in a plant cell in any greater detail than we did in biology class. The sizes are so small that the measures are not ones to which the gardener has a reference point. So, we’re left with imagining what things are like in a cell.
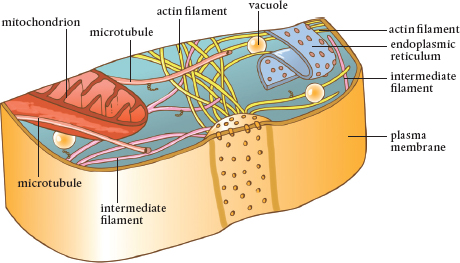
There are thousands of tiny filaments in a cell. Actin filaments are the smallest, followed by intermediate filaments, and larger filaments known as microtubules. These elements form a transportation system, a set of tracks, along which things can be moved around the cell.
Let’s try to put things in perspective by reducing ourselves down to the size where we can really relate to a cell. If a cell were 300 million times larger, the nucleus would look like a sponge ball about 50 feet across. Depending on where we are located, at this diminutive size an entire cell looks like a football stadium.
Unfortunately, due to all of the snowflake-sized molecules floating around, laced with hail-sized proteins, it is difficult to see anything clearly. Actin filaments are the diameter of a salt shaker, the intermediate filaments are halfway between that and the larger microtubules, which are the diameter of a beer mug. They hang about like cobwebs, and they are everywhere.
Speaking of cobwebs, tarantula-sized ribosomes appear all over the place, floating around in the jelly-like cytosol. And you can see them hanging on the rough endoplasmic reticulum, hundreds and hundreds of them. The endoplasmic reticulum is huge, extending from the nucleus to the plasmalemma, its sacs about as wide as an adult hand. While it extends the length of the cell, it only covers about a third of the cell’s width. It is almost like a slide, protected from the jelly-like substance that fills the cytoplasm. It is much easier to travel the endoplasmic reticulum than in the cytosol.
Mitochondria are now the size of tractor-trailers. You may see some being pulled along the microtubule network. Lysosomes, grinding away at their contents, are spheres that range from the size of softballs and soccer balls to huge medicine balls with diameters up to three times the size of a human. That Golgi apparatus you almost stepped on is twice the size of your offending foot and about as wide. You wonder where the other four or five are, so you won’t step on them, too.
You check out the apoplastic pathway by wandering around the cell wall system. It is surprisingly like thin cardboard or sliced luffa sponge. You can see the cellulose fibers, like wires; they are extremely strong.
You find the plasmalemma. This membrane is thicker than the cellulose fibers, but not by much; it has the thickness of this book. And it is constantly flowing, like oil in water, only it has all of these shoe-sized proteins sticking through it along with rope-like chains, which must be some sort of signaling molecule. Watch out. Don’t get sucked into an aquaporin or shocked by some of the peanut-sized charged hydrogen ions that are affixed to the surface of the cell wall.
Glowing ATP molecules are breaking apart all around, bursting into sparks like fireworks, and then the glowing stops as their energy is spent. Everywhere there are enzymes that are constantly changing shapes, attaching to and detaching from other molecules. The activity is overwhelming, and the noise is fantastic. Things are forming everywhere in incredible numbers and then being transported. Motion is constant.
Somehow, all of this activity moves forward. Energy is supplied, and if the nutrients are available to be converted into construction material, a tremendous amount of synthesis goes on inside the cell—building, repair, changing construction as a result of a signal sent through a carbohydrate molecule running through the cell membrane. Multiply this activity by 15 trillion and you get a birch tree. It’s amazing.
To be able to really see what goes on in a cell would be mind boggling. You probably wouldn’t be able to take too much of it and would want to get back quickly to read the next chapter in the story of how plants eat. We will continue this story by moving on to some chemistry and a bit of botany. All will be connected, and things will become clearer.