8 Biology of the oceans
For many the attraction of marine science is the variety of beautiful, and at times simply bizarre organisms that live in the oceans, although thankfully not of the size and ferocity depicted in Figure 8.1. Of course, over and above this the oceans, and especially shelf seas, provide considerable amounts of food for humans to eat.
The whales, at the apex of a complex food web, enjoy a certain charisma among marine animals. A whale can easily weigh around 100 tonnes. This compares with the bacteria at the other side of the web, which can weigh around 0.1 picogram (0.1 x 10–12 g) – a difference of 21 orders of magnitude. To help put this into perspective, the total biomass of bacteria in the surface waters of the Southern Ocean is estimated at around 3.2 × 107 tonnes, whereas the biomass of whales in the same waters is only around 8 × 106 tonnes.

Figure 8.1 The creatures of the sea are many and varied, but thankfully not as fearsome as those depicted in this section of the Carta Marina by Olaus Magnus, which was first printed in 1539.
The current estimate of total number of species of eukaryotic organisms in the oceans (plants, animals, algae, protozoans) is around 230,000. It is thought that in the region of 140,000 of these are known to scientists. In contrast there are estimated to be up to 1,000,000,000 species of bacteria (prokaryotes) in the oceans, of which only a minute fraction have been described. So it is very clear that we still have very much to discover about the biota of the oceans.
8.1 Zones of life in the oceans
At the onset of a general description of life in the oceans, it is useful to place the organisms into groups. The most obvious separation of groups is between those that live attached to or within the sediments or benthos (benthic), and those that live in the water (pelagic). Looking at the former, some benthic habitats are always covered by the water, even at low tide (subtidal) and others are periodically uncovered as the tide goes out (intertidal). It is important to remember that many of the species living in soft sandy or muddy benthic habitats are not visible, since they burrow into the sediments. In this short introduction we will not really touch on benthic organisms.
The upper 200 m of the pelagic is called the epipelagic, which corresponds to the depth of the continental shelf. Underneath this, mesopelagic extends from 200 m down to 2000 m, the bathypelagic from 2000 to 4000 m, the abyssopelagic from 4000 to 6000 m. The deepest zone is the hadalpelagic, which extends from 6000 m to the bottom of the deepest trenches around 11,000 m (Figure 8.2).
There is life in every part of the oceans, and sophisticated deep sea missions have brought back video footage of creatures living in the deep ocean trenches, although most of the life at such great depths is microbial (Chapter 12). However, the majority of the marine biota is concentrated in the surface waters, and as a general rule the total mass of biota per unit volume of seawater decreases with depth. This is because most of the food to sustain the food web is produced in the photic zone. Just as on land, the fundament of the food web are those organisms that can photosynthesise, utilising sunlight, carbon dioxide and water to create new organic matter for growth (Chapter 9). On land this is done by the plants, whereas in the ocean this is accomplished by the algae and some bacteria. As described in Chapter 7, the photic zone only extends down to a maximum of between 250 and 300 m in the very clearest of waters, and mostly at a depth of 100 m or less.
8.2 Life in the pelagic zone
The organisms living in the pelagic are crudely divided into the plankton and nekton. The latter are those species that are strong enough swimmers to be able to move against a water current (e.g. large crustaceans, fish, squid and whales). The plankton are the group of organisms that, although they may be able to swim, cannot swim against an ocean current, and so effectively drift with the water mass (from the Greek, plantos that can be roughly translated as ‘wanderer’). The organisms that make up the plankton are generally classified by their size, as shown in Table 8.1. The other main classification of the plankton is the organisms that can photosynthesise (mostly unicellular algae, some flagellates and some bacteria) called the phytoplankton, and the non-photosynthesising animals and protozoans referred to as the zooplankton.

Figure 8.2 Depth zone classification in the oceans.
Table 8.1: Size classes of plankton. It is important to remember that there is considerable overlap between these rough classifications. Organisms less than 200 µm in size can only be seen with good microscopes.
| Name | Size range | Examples of typical organisms in each size class |
| Femtoplankton | 0.02 to 0.2 μm | Viruses & small bacteria |
| Picoplankton | 0.2 to 2 μm | Bacteria, flagellates |
| Nanoplankton | 2 to 20 μm | Algae, flagellates, protozoans |
| Microplankton | 20 to 200 μm | Algae, flagellates, protozoans, crustacean juveniles, larvae |
| Mesoplankton | 0.2 to 20 mm | A few algae, crustaceans, small jelly fish, larvae |
| Macroplankton | 2 to 20 cm | Large crustaceans, jelly fish, larvae |
| Megaplankton | > 20 cm | Jelly fish |
But even at these most crude of classifications there are exceptions to the rule, since many benthic organisms that live in their adult stage on or in the benthos actually have larval (singular = larva) stages that live in the pelagic. These include fish, molluscs (e.g. mussels), crustaceans (e.g. barnacles or crabs), sponges, worms, echinoderms (e.g. starfish or sea urchins). The release of the larval stages (or meroplankton), or gametes (e.g. by seaweeds and corals) into the open waters is a key way that adult populations can ensure that their offspring are dispersed as far as possible, since many of them cannot move very far, or, as is quite often the case, not at all. Organisms that spend all of their life in the plankton are called haloplankton.
8.3 Femto- and picoplankton
If you take 1 ml of seawater from most oceans and pass it through a filter with holes of a diameter 0.02 µm (0.02 x 10–6 m), there will be approximately 1 million bacteria cells retained on the filter surface. There will also be around 10 to 100 times more viruses on the filter as well. Viruses are not living organisms, since they cannot grow or replicate themselves without infecting a living cell. However, they are included in this biology of the ocean section, since viral infections can have profound consequences for the rest of the biology. The study of viruses in marine systems is fairly young (two to three decades), but they have been shown to infect most other organisms in the ocean, and during viral epidemics they can result in the very striking death of swarms of crustaceans, algal blooms, seaweeds, fish, whales, etc. Again, to show how important the viruses are – assuming the volume of the oceans is approximately 1.3 x 1021 litres, then there are roughly 4 x 1030 viruses in the oceans. These contain about 200 million tonnes of carbon, which would be the equivalent of the carbon contained in 75 million blue whales (which are about 10% carbon).
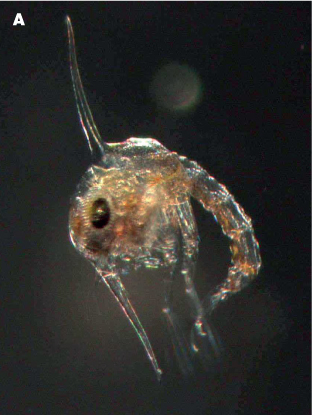
Figure 8.3 (A) Two larval stages of a crab Zoea.

Figure 8.3 (B) Two larval stages of a crab megalopa.

Figure 8.4 High power microscope image of bacteria stained with fluorescent dye. N.B: this sample has been concentrated, and in normal seawater there would only be one or two cells in this field of view. The big bright spot at the front is a ciliate.
Much more is known about the bacteria and archaea and their role in marine ecosystems. Both bacteria and archaea are prokaryotes, which means that they do not have a membrane-enclosed nucleus or other cell organelles (cf. eukaryotes, which have complex nuclear membranes and membrane-bound mitochondria and chloroplasts – including the plants, fungi, animals and algae). Many of the bacteria in aquatic systems grow by breaking down organic matter dissolved in the seawater (dissolved organic matter, or DOM, introduced in Chapter 7). The DOM pool in the ocean comes from land via rivers and estuaries (allochthonous DOM), but also from dead and decaying marine organisms, and as faecal matter and secretions of material (e.g. slime) produced by many marine organisms (autochthonous DOM). The significance of bacterial breakdown of DOM will be discussed in chapters 8 and 9. Although bacteria and archaea are ubiquitous in marine waters and sediments, the archaea are noteworthy in that they are frequently found in extreme conditions of high temperature (e.g. hydrothermal vents), hypersaline waters and anoxic sediments.
Some bacteria are able to photosynthesise and utilise sunlight in a similar way to algae and plants (chapters 8 & 9). These include the cyanobacteria (formerly called blue-green algae), and there are many species that fall into these small size classes. They are thought to be some of the most numerous photosynthetic organisms on the planet, and are present in most waters, although remarkably, they have only been studied in detail since the 1980s. There are also species of photosynthetic eukaryotes in the picoplankton, although these are even less studied than the cyanobacteria.
8.4 Nano- and microplankton
At the small end of this size range the species are dominated by both photosynthetic and non-photosynthetic flagellated organisms. The latter get their energy by feeding on picoplankton-sized organisms, and they generate feeding currents by moving their flagella to waft food items towards them. They are voracious grazers and have been estimated to be able to process 100,000 times their own body volume of water per hour. This is essential, since although there are millions of picoplankton per millilitre of water, living organisms in fact fill only a small fraction of this volume, and the encounter rate between a flagellate and food particle is very low.
There are a host of other grazers ranging from 20 to 200 µm, such as armoured dinoflagellates and ciliates. The former can even digest prey larger than themselves by exuding a sinister structure called a pseudopod that envelopes the prey item and encases it. Enzymes are then released to break down the prey, and once this is complete the pseudopod is retracted with the digested remains. Ciliates use another effective technique, by beating numerous hair-like structures (cilia) to filter particles out of the water. Foraminifera, also unicellular organisms, are another example of grazers that are able to exude pseudopodia (in this case called reticulopodia) into the surrounding water to catch their prey. These can extend up to 25 times more than the size of the individual producing them, thereby forming a huge net to catch prey. Foraminifera have shells (tests) made from calcium carbonate. In some regions of the world’s oceans there are dense accumulations of dead foraminiferal tests (foraminiferal oozes), well preserved in the sediments, that are used by palaeo-oceanographers to describe climate and ocean conditions over geological time.

Figure 8.5 High power microscope image of a ciliate. The feather-like cilia create a vortex to concentrate food particles (bacteria and phytoplankton), which are then ingested.
The most obvious members of the nanoand micro-phytoplankton are the unicellular photosynthetic algae, well represented by a diverse group of species called diatoms. The characteristic of all diatoms is that they produce cell walls, or frustules, made of a hard silicate, effectively glass, which are not only very beautiful to look at, but are also very strong. This wall has holes in it, often laid down in very beautiful patterns, which allows water, gases and nutrients to be exchanged. Many diatom species also have highly ornate frustules, with spines, spikes, hooks, and other protrusions. These adaptations are thought to slow down sinking rates, but also to deter grazers from attempting to eat them: the spines of diatoms have even been known to clog fish gills and pierce delicate membranes in gill tissues.

Figure 8.6 Electron microscope image of a dinoflagellate.
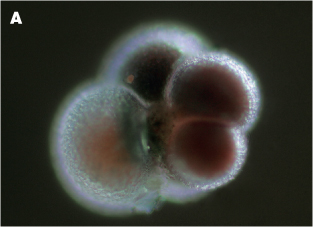
Figure 8.7 (A) A living pelagic foraminifer, Neogloboquadrina pachyderma

Figure 8.7 (B) a scanning electron microscope picture of the same species preserved in a marine sediemnt.
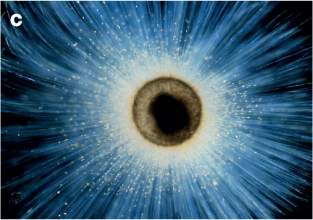
Figure 8.7 (C) Foraminifer, Orbulina universa with extended pseudopodia to catch food.

Figure 8.8 (A) Diatoms close up.
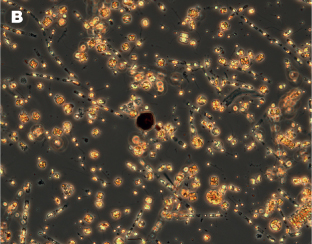
Figure 8.8 (B) during spring bloom.
Diatoms can form dense blooms in coastal waters, and are an important food source for protozoan, and especially zooplankton grazers. These blooms can be so dense and extensive that they are easily seen by colour sensors on satellites (see Chapter 14). However, once formed the frustules dissolve very slowly, and some ocean floor sediments are described as diatomaceous or siliceous oozes – massive accumulations of diatom frustules that have sunk from the surface waters. Like the foraminiferal remains described above, these diatom records are prime resources for palaeo-oceanographers for reconstructing past ocean and climate events. Diatomaceous earth can also be mined, and was a key component of the first dynamite invented by Alfred Nobel in 1867.
Diatom blooms are not the only micro-phytoplankton that can be seen from space. The coccolithophores are a group of photosynthetic algal species that are equally beautiful as the diatoms when looked at under the microscope. Instead of being encased in silicate, these single-celled organisms are covered in intricately patterned plates of calcium carbonate (coccoliths). They can also form extensive blooms where the ocean surface turns a milky white. One of the well-known species, Emiliana huxleyi, forms blooms in the north Atlantic covering an area of the ocean equivalent to the size of Great Britain. Due to the highly reflective nature of the white coccoliths, these blooms are easily visible from space, and so can be tracked using satellite imagery. The coccoliths sink and are incorporated into sediments, where they can accumulate, locking up huge amounts calcium carbonate. The impressive chalk ‘white cliffs of Dover’ are largely made up of calcium carbonate from coccolithophores.
8.5 Meso- and macroplankton
There are very few phytoplankton in the mesoplankton, and none in the macroplankton, and so these size classes are dominated by zooplankton species and both halo-and meroplankton larvae. One of the biggest groups of species in this size range is a group of crustaceans (i.e. having an exoskeleton), the copepods, of which there are thought to be over 2000 species. These are hugely important since they are a primary food source for many fish species and even some whales. Copepods largely feed on the algae and other organisms in the smaller size classes by filtering them out of the water (herbivores or omnivores). However, there are some species that are carnivores, hunting other copepods and having heavily armoured mouth-parts with which to tear apart their prey. Although they cannot swim against currents, they are still good swimmers, and when avoiding predators can swim up to several hundred body lengths per second! Copepods have male and female adults and complex life histories. In some species, after mating the female carries the fertilised eggs in specialised egg sacs, whereas in other species the fertilised eggs are released into the water. When the eggs hatch they release larvae called nauplii. These undergo five further growth phases before reaching a miniature adult-shaped stage (called a copepodite stage). As they grow larger the copepodites undergo five more developmental stages before reaching adult maturity.

Figure 8.9 Coccolithophore bloom in the Barents Strait that is so extensive that it is visible to satellites.

Figure 8.10 Adult copepod.
Towards the larger end of the macroplankton, the crustaceans are also represented by the Euphausiids or krill. These are a diverse group of species that range in size from 1 cm to around 10 cm, the most well-known species of krill being Euphausia superba, the Antarctic krill, which can form swarms akin to locust swarms. These swarms can be so dense that they have been recorded as turning the ocean a blood-red colour. Krill are present in such huge stocks in the Southern Ocean that they are estimated to exceed 1.5 billion tonnes (cf. the total mass of all the people on the Earth is approximately 0.5 billion tonnes). Large, dense krill swarms contain up to 30,000 animals per cubic metre, and since each individual can eat up to 25% per cent of its body weight per day, a krill swarm can very effectively clear a water column of any food particles. In turn, krill are the primary food for squid, penguins, many seal species and many baleen whale species.
Some of the most beautiful organisms in this group are the ctenophores (comb jellies) that have rows or fused plates of iridescent cilia which they use to swim with. They are gelatinous multicellular organisms and have specialised guts (therefore different from the ciliates described above, which are unicellular with no specialised guts). Many species have long tentacles that can be extended. These contain sticky substances with which to catch their prey, which includes copepods and other similar sized organisms. Salps are another frequently encountered gelatinous group in the plankton. They are found in the plankton as either individual transparent tubular bodies, or in long chains of individuals. They are very efficient at filtering particles from the water, especially phytoplankton cells.

Figure 8.11 The Antarctic krill, Euphausia superba. In 2009 a megaswarm of krill was discovered by scientists working in Wilhelmina Bay on the western Antarctic Peninsula. The swarm extended from the surface to 400 m deep and was estimated to contain 2 million tonnes of krill. In turn this attracted 500 humpback whales to feed on the swarm.
It would be normal to think of molluscs (snails and bivalves) as primarily benthic organisms. However, all benthic molluscs have larval stages that live as meroplankton. Moreover, there are several groups of molluscs that are haloplankton, and even some that have heavy shells. A good example of the latter are the pteropods (winged foot), whose feet (the part of a snail that is normally used to move along a solid object) are flattened into a set of paired ‘wings’ with which they swim. Some pteropods produce mucous floats to help them keep suspended, whereas other species have lost the heavy shells altogether, although they still use mucous nets to help keep them suspended and for catching prey.

Figure 8.12 Ctenophore or comb jelly.
Some of the most fearsome predators of the plankton world are the chaetognaths (arrow worms) which can be between 1 and 10 cm long. They are very commonly found in samples of plankton. They are transparent, streamlined in shape, and are effective swimmers that can dart forward with great speed to catch their prey (often copepods and larvae). Some species of arrow worm also use powerful neural toxins to kill their prey before ingesting them.

Figure 8.13 A pteropod or ‘flying snail’.

Figure 8.14 The mouth of a chaetognath, a voracious preadator of zooplankton and small larvae.
It is pertinent to point out that many species of zooplankton are good enough swimmers that they take part in a diurnal vertical migration (DVM); many populations of zooplankton are observed to move up from depths between 100 and 400 m at dusk and down again at dawn in a highly synchronised way. The most popular explanation for this is that by spending periods of strong light during the day at depth means that they avoid predators like fish, birds and squid that use vision to seek prey. Then by moving up to the surface at night they are able to feed. However, it is not necessarily so simple, and many ecologists have spent considerable effort in trying to unravel the secrets of this phenomenon. For instance, one theory is that the zooplankton migrate to deeper waters in the day to avoid damaging ultraviolet radiation. Clearly the primary trigger to induce the behaviour is light, but temperature is also thought to play a role, and seasonal differences are observed in some water bodies where DVM takes place in the summer but not in winter.
Some species of copepods are also known to migrate to deeper waters to overwinter when food is scarce in the surface waters. They enter a physiological state called diapause, where they become physically inactive, don’t feed, reduce their metabolism to a minimum and sink to deep waters where they are neutrally buoyant (where their density is the same as that of the surrounding seawater). Depending on the species, the copepods can overwinter in this state at depths between 500 and 3500 m.
8.7 Megaplankton
The most noticeable megaplankton are the jellyfish (belonging to the group Cnidarians, which also includes sea anemones, corals and the comb jellies mentioned above). Although jellyfish swim effectively with rhythmic contractions of their bell, they are unable to swim against ocean currents, and so drift where the water masses take them. This is surprising, since several species can reach huge sizes, for instance the Arctic ‘lion’s mane’ jellyfish (Cyanea arctica) which has tentacles up to 40 m long.
Jellyfish are mostly carnivores feeding on zooplankton, larvae and small fish. To capture their prey many species trail tentacles laden with specialist cells called nematocysts. When touched, these cells can release a hollow, harpoon-like structure that anchors the prey (zooplankton and larvae) and injects a powerful toxin to immobilise it.

Figure 8.15 A compass jellyfish caught in the Namibian Benguela, where the biomass of this species now exceeds that of commercially important pelagic finfish. NB the deck in the background is awash with them.
Many of the smaller jellyfish species, more typical of macroplankton, have a plankton medusa phase, which alternates with a polyp stage that grows attached to the benthos or structures in the sea. The polyps produce and release larval medusae that eventually develop into the more visible jellyfish.
Some jellyfish species, such as the siphonophores, are colonial without any polyp stage. They are remarkable in that the individuals within the colony adopt different functions for propulsion, feeding, or reproduction. In the Portuguese Man O’War (Physalia physalis) one part of the colony develops into a large gasfilled float (up to > 30 cm high) which keeps the colony suspended at the water surface. It is hard to see one of these structures and to realise that it is not a single organism. These also have tentacles laden with nematocysts, which in the largest of specimens can trail below the float for several tens of metres.
The potency of the toxins released by the nematocysts is well known to any swimmer or diver who has had the misfortune to touch a tentacle. The encounter results in what can be an intensely painful experience. However, the toxins produced by the sea wasp or box jellyfish (Chironex fleckeri) are so potent that they have caused several human deaths. For this reason, beaches are often closed if they are spotted in the water.
8.8 Nekton and others
The marine organisms of the nekton are better known and include fish, reptiles, squid and mammals, including the toothed and baleen whales, sea otters, manatees and dugongs. There is a whole group of other marine animals that are clearly able to swim against a horizontal current, and so are nekton, but which can also have periods out of the water. These include polar bears, seals, sea lions, saltwater crocodiles and turtles. Clearly, fish and squid are only nekton when adults, since their egg and/or larval stages take place among the plankton.
It is impossible to do justice to such a variety of animals in just a few short paragraphs. However, by being able to swim so effectively many of these organisms, such as the baleen whales and many species of fish, are able to roam large distances on tremendous migrations through many ocean realms. Naturally, by being able to swim effectively, some of the animals are able to live in deeper waters and migrate to surface waters just to feed.

Figure 8.16 (A) Many of the larger animals of our oceans spend time outside the water, but are all part of the nekton Polar bear.

Figure 8.16 (B) Many of the larger animals of our oceans spend time outside the water, but are all part of the nekton Leopard seal.
Finally in this section, it would be wrong not to introduce a key group of organisms, other than humans, that feed on the animals of the nekton. Although not part of the nekton, there are the myriad of bird species that exist by feeding on fish and zooplankton mostly in surface waters of the oceans (0 to 50 m deep). However, some, such as the Emperor Penguins (Aptenodytes forsteri) are such exceptional swimmers that they can dive to depths of 500 m in search of fish, squid and krill. The rich diversity of bird species and the huge biomass of birds that the oceans support mean that they are an integral part of the ocean ecosystem.
8.8 Living and sinking in a viscous medium
The experience of movement in water is very different from that in air, since water is more viscous than air. The viscosity of the medium (the resistance of a fluid to flow under the influence of an applied external force) is dependent on the density of the water, and hence temperature and salinity. A term called the kinematic viscosity is a measure of the drag on objects moving through the fluid, and for that object inertia tries to keep the object moving, whereas viscosity tries to stop it. Basically the greater the viscosity of a medium, the more slowly an object will move through it; for instance, olive oil has a much greater viscosity than water, and a stone will sink more slowly in the olive oil compared to the water.

Figure 8.17 (A) Birds are an important part of the open ocean ecosystem, e.g. Giant petrels that seem to appear out of nowhere when some debris is caught up on the water surface.

Figure 8.17 (B) Birds are an important part of the open ocean ecosystem, e.g. Emperor penguins.

Figure 8.17 (C) Birds are an important part of the open ocean ecosystem, e.g. Gannets.

Figure 8.17 (D) Birds are an important part of the open ocean ecosystem, e.g. Puffins.
For the movement of any particle in a specific medium we talk about the Reynolds Number (RN) which is derived from:
RN = (particle velocity × particle size)/kinematic viscosity
Very crudely, the smaller the organism the smaller the RN: a large whale swimming in seawater at 10 m s–1 has an RN of 300,000,000; a large fish swimming at the same speed an RN of 30,000,000; a copepod swimming at 0.2 m s–1 an RN of 300; and a bacteria swimming at 0.01 ms–1 an RN of 0.00001.
The consequence of this is that for small organisms the fluid is effectively highly viscous, and a moving object has little momentum: e.g. when a bacteria stops swimming its momentum will keep it coasting for less than 10 microseconds (around 1 nm), whereas a large fish or a whale will continue to glide a considerable distance through the water, even though it has stopped active swimming. This lack of momentum on the micro-scale means that when you look at micro-organisms through a microscope they appear to be moving through oil or thick syrup.

Figure 8.18 As shown here, the spines and shape of diatoms will greatly influence their rate of sinking through the water.
Naturally, these effects translate into how fast different-sized organisms sink in the water column, and this is of immense consequence, for instance, to photosynthetic organisms wanting to stay in the photic layer. Stokes’ law describing falling spheres can be used to calculate the terminal velocity of an organism at low Reynolds numbers (effectively for particles/organisms up to 500 µm in diameter). The rate of fall of a sphere (of radius r) is described by the equation:

where V = velocity of particle, g = acceleration due to gravity, ρ = density of particle, ρ = density of water, n = kinematic viscosity of water. Therefore a particle’s rate of sinking increases in proportion to the square of its radius. Observations of sinking rates of phytoplankton cells actually show that their sinking speed increases with cell radius raised to a power lying between 1 and 2. The reason is that in many cases the density (ρ) of the cell decreases with cell size, and this lowers the power to which the size is raised in this sinking speed equation.