
Having surveyed species numbers around the world, let's continue by looking for unusual patterns of diversity. But before we do that, we should re-examine the nature of the information available in making our assessments. How do we actually quantify biodiversity? Three measures have been given special attention regarding local biodiversity. We've already mentioned the first; the actual number of co-occurring species in the area. This information is simple and is usually available for the better-known lineages. However, there are problems with even a simple species listing.
A primary concern is counting only those species which properly belong to the fauna or flora. This requires being able to distinguish those species that, while perhaps appearing indigenous today, were actually schlepped in by human activities. Nowadays, escalation of travel and commerce only adds to the difficulty of separating indigenous from non-indigenous species.1 Consequently, we will focus our attention on the native species of a region—not recent arrivals.
Another measure of biodiversity is a bit more subtle; it is not the number of species in an area but the number of species found nowhere else: the endemics. And here we can run into taxonomic problems. Is a local endemic species really that, or is it just a peculiar local variant of a more widespread species? So-called “good species” are the result of careful study, broad comparisons, and informed taxonomic decisions. Unfortunately, such a foundation is not available for many plants and animals. Nevertheless, when a region claims a high percentage of endemics there is the implication that we are picking up an important historical signal.
Finally, there is diversity at higher taxonomic ranks or more distantly related lineages. Imagine finding a small island with three species of mice on it and no other vertebrates. How would such an island compare with a similar island that had one mouse, one lizard, and one frog species? Surely, Mouse Island doesn't have the same level of biological diversity as does the island home of three different vertebrate lineages. From a conservation standpoint, there is an important difference. Mouse Island does not deserve as much effort as the island with three different vertebrate lineages. Think of these islands in terms of future potential; on Mouse Island, we can only hope for more mice.2 But before we discuss diversity at higher ranks, let's return to simple species numbers and look at one of the most obvious patterns on the planet.
THE LATITUDINAL SPECIES-DIVERSITY GRADIENT
Starting at the equator, and moving towards either pole, overall species numbers decline. There may be areas where the trend is locally reversed——especially when traversing the great deserts——but the overall trend holds true. Species numbers are greatest in warmer regions, and progressively diminished toward the poles. This general pattern is found both on the land and in the sea.3 Tree species numbers decrease steadily as we move northward toward the Arctic. Along the western Pacific we can expect to find forests with around 500 tree species in Taiwan, 250 species in southern Japan, about 80 in northern Japan, 20 on Russia's Kamchatka Peninsula, and only about 5 species above the Arctic Circle. Shifting to the western edge of North America, we can see a similar pattern in birds and mammals, only here we'll move southward. There are 222 species of breeding birds and 40 mammal species in Alaska. To the south, in British Columbia, Canada, we find 267 breeding bird species and 70 mammal species. In California, these numbers are 286 and 100. Moving to tropical Mexico the numbers explode, with 772 breeding birds and 491 mammal species. While some of this increase in numbers is due to Mexico's large area and diverse topography, there is a clear tropical effect. Butterfly numbers are just as dramatic. All of North America north of central Mexico is home to about 750 butterfly species; to the south, the Neotropical realm numbers an estimated 7,500 species; a ten-fold difference!
Of course, the latitudinal species diversity gradient has exceptions; some groups of plants and animals are more diverse in the cooler temperate zone. The rose family, the pines and birches, oaks, aphids and salamanders, all have more species in temperate areas than in the tropics. But these are exceptions. The generalization holds true: as you leave the tropics and move toward the poles, species numbers decline. This is especially visible in some iconic tropical plants. Palms, with over 2,500 species, have a very limited frost tolerance; coastal North Carolina is as far north as they naturally grow in North America. Even more intolerant of cold are the gingers, the bananas, and their kin. Often with huge leaves and colorful inflorescences, these plants give moist tropical vegetation much of its exotic appearance. But these are all specific examples; what might be the general causal factors for increased tropical species richness?
Surely simple biological productivity has to be a key factor, whether measured as annual carbon fixation or standing biomass. Given sufficient moisture and agreeable temperatures, more direct sunshine means more photosynthesis (more carbon fixation) and that means more fuel to keep the ecosystem running. In addition, neither cold nor drought is conducive to life activities. Plants can't photosynthesize while they are freezing or when they are parched. A clear example of the tropical advantage is seen in leaves. There is nothing in cooler northern climes that compares with the leaf of a banana plant or coconut palm. Large leaves are expensive to produce, and you find them only in the evergreen tropics, where they can continue photosynthesizing over several years.
This brings us to the problem of evapotranspiration, a measure of moisture loss from plants. When things get too dry, pores in the leaves close and photosynthesis shuts down. In severe cold, the plant cannot pump water up from its roots and, again, the plant must go into dormancy. Thus, seasonality is a critical determinant of plant productivity. Even though they may feed on ocean fish, a study of island-living birds around the world concluded that climate——temperature and precipitation——is the most important determinant of species numbers.4 Staying with birds, frigid Greenland is home to 57 species of breeding birds, while the state of Georgia in the eastern United States numbers 160 species. Further south, breeding birds find their zenith in Colombia, with around 1,600 species. Here's as dramatic an example of the latitudinal species-richness gradient as you are likely to find.
The geographical range of species is another factor contributing to the latitudinal diversity gradient. For example, if you are a species that can survive a typical Midwestern year, from subfreezing winter through torrid summer, you are likely to be able to range over a wide area. But if you are adapted to the cool misty interior of a tropical montane cloud forest, you are unlikely to be successful in the hot lowlands, or in any other stressful climate. In addition, it is unlikely that your seeds can reach a similar habitat on a distant mountainside. For this reason, montane cloud forest species often have very limited geographic ranges. Ecotourists visiting Costa Rica's Monteverde Cloud Forest Reserve can experience this unusual climate for themselves. Tourist hotels near the reserve offer comfortable first-class accommodations, and yet they do this without heating or cooling systems! At 1,500 m (5,000 ft.) above sea level, Monteverde provides a truly temperate experience. It's warm every afternoon and cool every evening, throughout the year!
Beyond the cloud forest and more generally, tropical environments support more “specialist” species than do strongly seasonal environments. In a recent comparison of butterflies, tropical forest caterpillars were shown to have more specialized diets than their temperate forest counterparts.5 This means more species can coexist without getting in each other's way. A more dramatic example is found among Costa Rica's mammals; a large number of species are fruit-eating and nectar-slurping bats! Such lifestyles are only possible in an environment where flowers and fruit are available throughout the year. Having worked with Costa Rica's flora over more than three decades, I was impressed by how much that flora differs from the floras of nearby Nicaragua or Panama. That's not what you see when you leave Indiana to visit Illinois to the west, or Ohio to the east. We had thought that Illinois had a single endemic flowering plant: the Kankakee mallow (Illiamna remota). But then this same species was found in West Virginia, and Illinois lost its only living endemic flowering plant. Midwestern floras are nearly identical, and why shouldn't they be? Large areas of the Midwest were sitting under an ice-sheet 20,000 years ago. Nearly all these plants are recent re-colonizers. Nicaragua, Costa Rica, and Panama have suffered no such indignity; their species have been free to sit where they are and diversify over millions of years. All three countries have many endemic species. Simply put, a greater percentage of tropical species have more limited geographic ranges, compared with temperate or polar species. Called Rapoport's rule, this generalization applies to both plants and animals.6
A recent paper in Science looked at the latitudinal diversity gradient from a genetic perspective. These authors compared closely related species pairs of birds and mammals in both tropical and temperate environments, and estimated their times of divergence by using DNA sequences.7 The result was surprising and significant: tropical species pairs are generally further apart in time than are bird and mammal pairs from higher latitudes! Speciation is more rapid in cold climates than in tropical latitudes! At first, this seems counterintuitive. However, if species are going extinct more rapidly in cold climates, more vacant niches will be available for new species to invade. A higher extinction rate, followed by speciation, means that local turnover rates are higher at high latitudes. Certainly, extinction is a significant factor underlying the latitudinal species-diversity gradient. Just imagine what conditions were like in the American Midwest or central Europe during the height of the last glaciation.8
And then there's something else we don't like to talk about: parasites and pathogens. Our own species is a fine example. If you would like to get really sick, let me suggest visiting moist lowland tropical Africa. There are two reasons why Africa has so many human maladies. The first is the climate: many of our pathogens and parasites can't survive a northern winter. The second reason is historical: Africa is where our species originated and where our closest relatives reside. Here, our diseases had ample time to make us their hosts. Here is where the HIV virus originated, where malaria, yellow fever, liver flukes, and many other human pathogens abound. This same problem affects other species, both animal and plant: there are many more pathogens, parasites, and herbivores in the tropics. In summation, the latitudinal species-diversity gradient seems to be the simple product of more benign environmental factors and more competitive challenges in tropical latitudes.
AREA AND ALTITUDE EFFECTS ON BIODIVERSITY
The latitudinal species-diversity gradient is the most prominent area-diversity relationship on the planet. Also, and in general, as one moves from smaller to larger land surfaces one finds more and more species. This is the species-area relationship, and it usually forms a simple linear gradient from smaller to medium-sized areas. As one reaches ever-larger areas, however, fewer new species are encountered, and the graphed relationship tends to flatten out. As you might expect, the latitudinal species-diversity gradient interacts with the species-area diversity gradient. A recent study of North American vascular plants found that species-area relationships change with latitude. That is, as you go further north, species tend to have wider ranges; some are even circumpolar.9 An Illinois prairie can have as many as 300 species of higher plants in the prairie and its moist depressions. Then, as you enlarge your survey westward to include Iowa, you do pick up new and different species. But you add only a small number. As we mentioned, this is one way in which the tropics really are different. Beginning in Costa Rica starts you off with lots of species, but then, as you expand to either Nicaragua or to Panama, you will be adding many species you hadn't encountered in Costa Rica. Here, adding the local and wider areas really does escalate the numbers.
A striking example of the area-diversity relationship comes from a study of islands in the Caribbean, from small to large. With both species numbers and island area graphed logarithmically (1, 10, 100, etc.), the relationship results in a straight line, showing clearly how greater area supports greater diversity.
TABLE 5-1: SPECIES AREA RELATIONSHIPS AMONG PLANTS AND TREES
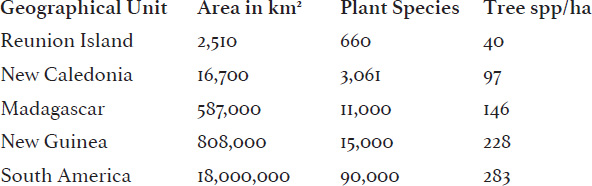
From a talk by E. G. Leigh Jr. at Field Museum, April 2007
An interesting aspect of the area-diversity relationship is that, on a small scale, the tundra can outnumber the rain forest. If you compare a square meter of tundra to a square meter on the floor of the rain forest, you'll find more species in your tundra quadrant! There will be mosses, liverworts, lichens, sedges, and surely a grass or two in your tundra plot. The dark floor of the rain forest may have a few seedlings, a sapling, and lots of rotting leaves, but not much more. In the rain forest, of course, much of the diversity sits high in the canopy. Even so, one reason the square meter of tundra is so rich is that the plants are so small. And the reason they are so small is because the environment is so severe. On a smaller scale (one to a hundred square meters) arctic and temperate vegetation can be impressive in their species numbers. However, it is at larger scales, where the moist tropics reign supreme—with bigger plants carrying lesser plants upon them. In fact it is the accumulation of little plants—growing as epiphytes on trunks and branches—that raises the numbers of plant species significantly in evergreen tropical wet forests. In an altitudinal study, ranging from 30 to 2,600 m elevation on the Caribbean side of Volcán Barva in Costa Rica, 264 species of ferns were found. Of these, 96 were terrestrial species, while 121 were recorded as low-trunk epiphytes, and 113 were found as high-canopy epiphytes. Though some species were recorded in more than one category, the epiphytic ferns greatly outnumbered the terrestrial ferns.10 Another recent survey estimated that around a quarter of all the species of native flowering plants in Costa Rica are epiphytes, and almost a half of Costa Rica's endemics are epiphytes!11 In fact, Al Gentry and Cal Dodson suggested that the extraordinary floristic diversity of the Neotropics was largely due to its epiphytes.12
A terrific way to experience the species-diversity gradient is to ascend from lower to higher elevations in the tropics. Again in Costa Rica, and studying ferns, Jürgen Kluge and Michael Kessler enumerated species along an elevational transect ranging over almost 3,400 m (11,150 ft.).13 They found about 20 species per plot at 100 m elevation, around 50 fern species in plots between 1,200 and 2,500 m, and around 10 species at 3,000 m. Both total fern species numbers and endemics were highest between 1,000 and 2,500 m elevation! Researchers in Borneo found a similar diversity pattern for moth species; the highest numbers were found between 500 and 1,000 m elevation.14 As in many other tropical studies, there is a clear “hump in species richness” at middle elevations. Because temperatures fall by about 6°C for every 1000 m (3°F per 1000 ft.) above sea level, air forced up the mountain slopes expands and becomes cooler. This cooler air cannot hold as much moisture, resulting in frequent rain and misting over much of the year. It is in these montane “cloud forests” where trees and their branches are festooned with smaller epiphytes—perhaps the richest biome on the planet.
It should be no surprise that the highest elevations of tropical mountains have low species numbers. Smaller areas and colder, more windy conditions are the reason. English speakers developed a phrase for the high elevation habitats they experienced in East Africa: “Summer every day and winter every night!” Above about 3,000 m (10,000 ft.) elevation, a clear night sky can drop temperatures to freezing. In this way, high tropical mountains offer a greater variety of temperature and moisture regimes than do the more uniform conditions in the hot lowlands. Varied altitudinal zones, in turn, allow for greater speciation, as ecological selection promotes diversification. Birds of Paradise living in New Guinea's mountains maintain high species numbers by living at different elevations, in what is called altitudinal stratification. By affording a great variety of habitats, mountain systems are a major reason why our planet is so rich in biodiversity.
NUMBERS OF SPECIES AND NUMBERS OF INDIVIDUALS
In both tropical and subtropical habitats, superabundant and widespread species tend to be few, while the great majority of species—animal or vegetable—are rare. This is especially evident in some of the large tropical trees, where you have to hike some distance to find another tree of the same species. And it is not just trees; many shrubs, herbs, and animals seem “always to be rare.” The meter-tall shrublet Piper veraguensis is very distinctive: its foot-long leaves hang vertically from leaf-stalks attached near the center of the blade, where smaller veins form circles around the stalk attachment. Though known from Costa Rica to Colombia, I have seen this species only twice in many years of field work in Costa Rica. Exactly how a species can manage to persist at such low population densities over so wide a range is a mystery to me.
Eric Dinerstein describes the importance of rarely encountered species, both for biodiversity and conservation, in his book The Kingdom of Rarities. He points out that some species are “doubly rare”—that is, both geographically restricted and rarely encountered in those areas where they do live. Another pattern is to travel for some distance through forests and over mountains and not see a particular species, only to come upon a particular valley and find it in considerable numbers. We botanists describe such a species as being locally abundant, while zoologists tend to use the word patchy. In an extensive study of Amazonian trees, a recent analysis found that only 227 species were “hyperdominant,” accounting for half of all the trees, while 11,000 species were rare.15 Surely, rare species are an important way of squeezing more species into the same geographical region. While these patterns have to do with contemporary species-packing, there are also distribution patterns with deep historical roots.
PLATE TECTONICS AND PATTERNS IN BIODIVERSITY
Scientists, like other human beings, are social creatures. If everybody in your discipline proclaims that the world's continents couldn't possibly move, you are not likely to sing a different song. Geologists in the United States held fast to their belief in the stability of the continents. But Alfred Wegener was a German meteorologist, and he was impressed by the shape of the Atlantic Ocean, with almost symmetrical eastern and western shores. Also, he had read the work of paleobotanists, who spoke of an ancient floristic Gondwanaland. Rocks from Gondwana (a little town in southern India) contained fossil leaf impressions that included a variety of different trees. One species, Glossopteris, had leaves rather like those of a willow. But there were also fern-like leaves and several other distinctive species as well. This 220 million-year-old fossil assemblage became known as the Glossopteris flora, characterized by a consistent suite of distinctive species. The co-occurrence of these tree species implied that they lived in the same kind of forest. And that was the problem! Rocks of similar age with this same suite of plant fossils were also found in Australia, South Africa, and South America!
Surely it's possible that one or two species might have been growing around the world in those ancient times——but all these species, always together? That seemed highly unlikely. Paleobotanists concluded that the Glosopteris flora had been a single forest spread over a single continent, calling this ancient continent Gondwanaland or Gondwana. With these and many other data, Alfred Wegener fashioned his theory of “continental drift” in 1915.16 Impossible! was the response of geological leaders in North America. After all, how could continents rupture or move around? Good scientists do good science, and continents drifting around the surface of the Earth was, clearly, bad science!17
But then, careful surveys of the ocean depths in the 1950s and 60s produced unforeseen discoveries. Nowhere did the ocean floor appear to be more than 250 million years old. This was totally unexpected! Furthermore, the floor of the deep oceans was crossed by long volcanic fissures. The Atlantic Ocean has a volcanic ridge-system running exactly down its center, all the way from Iceland to the Falklands! At about this same time, it had become known that Earth's magnetic field can flip, with the north magnetic pole becoming south, and back again. We still do not understand how or why the magnetic field changes in this way, but the reversal of magnetic poles could now be used to help date volcanic rocks! As lava cools and hardens, its magnetic polarity becomes frozen in place, and we can tell its north/south polarity at that place and time. Using this data to analyze lava beds east and west of undersea fissures produced an astounding result. Volcanic rocks on opposite sides of undersea ridges proved to be perfectly symmetrical in their pattern of magnetic reversals! The reversals on one side were the mirror image of those on the other side! Suddenly, geologists realized hot magma was welling-up along the centers of these undersea ridges, and then spreading laterally on both sides—producing symmetrical magnetic bands of lava rocks on either side as they moved apart. The mid-Atlantic ridge, by producing new magma at its center, has been expanding the ocean floor laterally, east and west!
Coupling the near-symmetrical continental shelves of the Atlantic with the mid-Atlantic ridge's continuing east-west expansion made something very clear: over millions of years, the eastern and western margins of the Atlantic Ocean were being continuously pushed further apart. Not all that long ago in geological time, there had been no Atlantic Ocean. Earlier, there had been a huge southern continent—Pangaea—on which the Glossopteris flora flourished. That land mass and its vegetation had been torn apart, just as Wegener imagined. Our planet's surface was made up of shifting plates! Surely one of the grandest revolutions in the history of modern science, the theory of Plate Tectonics now gives us deep insight into how the continents have changed over time. Little Gondwana and the Indian subcontinent on which it sits had been ripped loose from a larger southern land mass, then “rafted across” the Indian Ocean, only to smash into the Asian continent, driving the Himalayan mountains high into the sky.18 The theory of plate tectonics is Wegener's continental drift confirmed!
The notion of continents being torn apart and carrying their biotas with them became a new paradigm among certain biogeographers. Ridiculing the notion that plants and animals might disperse across vast oceans, these researchers promoted “vicariance biogeography.” Why do the flightless ratite birds all live on southern continents? Because their original home was in Gondwana, claimed these biogeographers. Unfortunately, recent DNA comparisons indicate the ancestors of these birds flew to their present homes.19 Lung fishes tell a more compelling story, with survivors living only in Australia, South America, and Africa. These do seem to be sitting on the broken parts of their original landmass. Vicariance biogeographers disparaged suggestions of cross-oceanic dispersal as untestable ad hoc hypotheses. Things have calmed down since those acrimonious debates in the 1980s, and people are now comfortable with the fact that some lineages have managed to travel across the oceans on their own, while others have held tight to their continental moorings.20 In fact, the idea of continental movements “having carried their living passengers with them” explains the world's most unusual animal disjunction.
WALLACE'S LINE
Supporting himself by collecting animal specimens, Alfred Russel Wallace traveled extensively between Malaya, Indonesia, and New Guinea. While collecting, he noticed something very unusual. Over a distance of as little as 48 km (30 miles)—between the Indonesian islands of Bali to the west and Lombok to the east—he had collected very different animals. There was a similar distinction in animal distributions between the islands of Borneo on the west and the Celebes to the east. Monkeys, cats, deer, rhinoceros, and squirrels could be found on the western side of this divide. To the east, Wallace found a world of strange arboreal marsupials, many lizards, birds of paradise and their relations—but no monkeys, no cats, no deer. Though a number of animal lineages cross Wallace's Line, the larger picture holds true. Here is a major division in animal distribution—a disjunction—over a very short distance!
How could two faunas be so close, and yet so distinct? Enter plate tectonics! As in the case of India, the Australian plate, carrying New Guinea and Australia with it, has moved northward from more southern climes. While not smashing into its northern neighbors with the force and effect of the Indian subcontinent, plate movement did bring a unique southern biota into close proximity with a northern fauna. This scenario explains why birds of paradise and bower birds are abundant in New Guinea: there are no cats to eat them. Thus, an explanation for two very different faunas in such close proximity is that two long-separated tropical faunas were brought into close proximity by tectonic movement.
Unlike the animal disjunctions, however, Wallace's Line is difficult to discern when examining the vegetation. Plants appear to have sent their disseminules back and forth across these marine barriers with great ease. Only a few nonflying terrestrial mammal families are shared between Southeast Asia and Australia. In comparison, sixty-nine flowering plant families are shared over these same areas. Plant species don't tell the same story as the animals.
Here is an instance where higher taxonomic levels, with a longer history, do retain a historical signal. Tropical American ferns number about 1,270 species, not very different from the 1,235 species listed for Southeast Asia, Australia, and the nearby islands (Australasia). However, the Australasian flora has 93 families of ferns and fern allies, while the Neotropical flora has only 38 families. That's a huge discrepancy! Australasia has more than twice as many families as all of tropical America. Fern families, a much higher rank than species, are telling us something. Twice as many fern families in Australasia supports the idea that two long-separated floras have come together and joined in this part of the world. Widely dispersed by minute airborne spores, species of the original Southeast Asian and New Guinea/Australian fern floras are no longer discernable.
THE EASTERN ASIA–EASTERN NORTH AMERICA DISJUNCTION
Asa Gray, professor of Botany at Harvard University in the mid-1800s, was the first to notice something unusual in northern plant distributions. Shipments of plant specimens from Japan and China often included genera also growing in the eastern United States. As collecting continued, something became very evident: there was a clear similarity in the floras of eastern Asia and eastern North America. In fact, for many groups, there was greater similarity between eastern North America and eastern Asia than between eastern North America and western North America! The flora of Oregon lacks many of the iconic plant genera shared by eastern Asia and the eastern United States. Tall and graceful, only two species of tulip trees are found in the world today: Liriodendron tulipifera in the eastern United States and Liriodendron chinense in eastern China. Our sassafras trees, often with mitten-shaped leaves, are members of a genus with three species: two live in eastern Asia and one is in eastern North America. Skunk cabbage (Symplocarpus foetidus) of our eastern swamplands is found in only two areas of the world: eastern Asia and northeastern North America. The same pattern holds true for lopseed (Phyrma leptostachya), a forest-floor herb.21
Shifting to animals, the strange paddle fish of the Mississippi River is a member of an ancient lineage with only one living relative, swimming in China's Yangtze River. A foot-long salamander that looks like it's been run over by a truck, the hellbender, lives in the Ozarks and Ohio River drainage; its only close relative lives in eastern Asia. The world has only two species of alligator—one in the southeastern United States and the other in southern China. What might explain these concordant disjunctions?
Twenty-million-year-old fossils tell us that Europe did have a much richer biota in those earlier times. The dawn redwood (Metasequia) was known as a fossil in northern regions around the world but was found as living populations in China only later. A rich biota of tulip trees, alligators and odd amphibians were all part of a northern “mixed-mesophytic forest” before the ice ages began. Since then, glaciation severely reduced the flora of northern Europe and, as we already noted, left eastern Asia the least diminished. Surviving species in eastern Asia and eastern North America have given us the biogeographic disjunction first noticed by Gray.22 Here again, history has left a biogeographic signature.
THE CONCEPT OF “BIODIVERSITY HOTSPOTS”
From a larger perspective, there are regions in the world where biological diversity is unusually high. A few of these areas have been labelled “hotspots” for the purposes of biological conservation. Unfortunately, and rather like beauty, special areas of biodiversity may lie in the eye of whoever is making up the listings. Criteria for assessing special regions are varied. How significant are overall species numbers in such determinations, and which plant and animal groups will be measured? Should species unique to the region—the endemic species—play a significant role in designating a hotspot? Should threat of imminent extermination be an important criterion? Though arbitrary, the notion of hotspots has been grandly envisioned in a large book with many stunning photographs: Hotspots: Earth's Biologically Richest and Most Endangered Terrestrial Ecoregions, by Russell A. Mittermeier, Norman Myers, and Cristina Goethsch Mittermeier.23 Using environmental threat and endemic species, these authors listed their hotspots. Inevitably, some choices seem arbitrary; New Caledonia and New Zealand are included, as are the islands of the Caribbean, but not New Guinea, where species numbers are high but threats are not as imminent. Also the areas covered by each hotspot vary greatly. Much of the central and western Pacific is assigned to the Polynesia/Micronesia hotspot—a huge area—but little New Caledonia is a hotspot all its own. Such choices are inevitable in any survey designating only twenty-five hotspots (Table 5-2).
TABLE 5-2: BIODIVERSITY HOTSPOTS (MITTERMEIER ET AL., 1999)
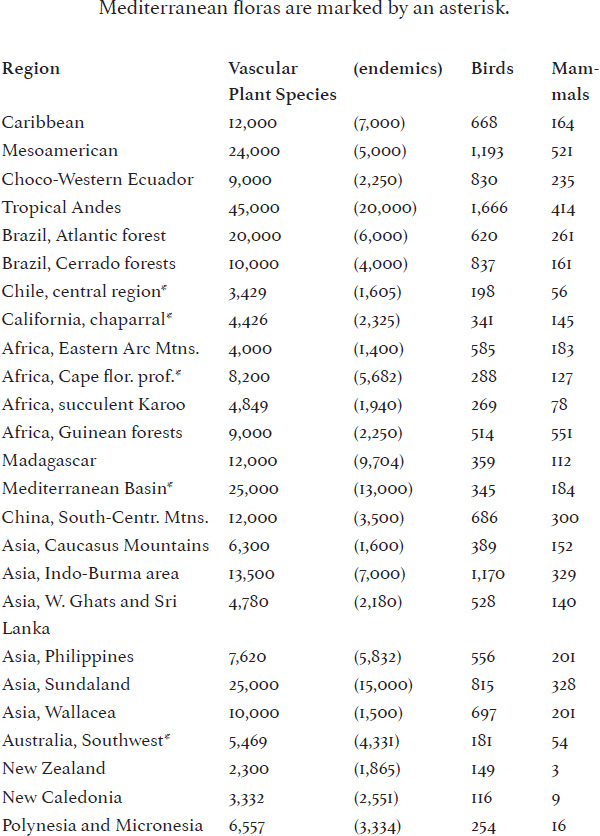
Numbers of vascular plants, mammals, reptiles, birds, and amphibians, are the basis on which these hotspots were chosen; these organisms are the ones we know the best, and the ones in which endemism is easy to assess. We've referred to these “hotspots” earlier and, as noted, one of the most significant criteria followed by Mittermeier, Myers, and Mittermeier for designating hotspots is advancing agriculture and continuing deforestation. In these circumstances, narrow-ranging endemic species are often the most susceptible to extirpation. The Caucasus Mountains, Africa's Eastern Arc Mountains, Madagascar, and the Philippines were all designated hotspots because of the threats they face. With the purpose of drawing world attention to the grave dangers being faced by specific areas, hotspots continue to play a role in conservation priorities. However, Peter Kareiva and Michelle Marvier have argued that our world also harbors important “biodiversity coldspots.” Yellowstone National Park may not have a large number of species, but it is the only place in the coterminous United States harboring the last assemblage of once widespread large mammals, such as grizzly bears, elk, mule deer, wolves, puma, and bison.24 Clearly, we need to preserve as many species as we can, and in every corner of the world that we can. That said, let's try and understand why some areas manage to support unusually high species numbers. We'll begin with little Costa Rica, which has played an important role in both tropical biological research and conservation.
COSTA RICA: A LITTLE COUNTRY WITH LOTS OF SPECIES
Costa Rica is a splendid example of tropical biodiversity. The people of Costa Rica have supported a vigorous intellectual tradition throughout their history. After World War II, they abandoned their army to spend more money on education. After many decades, and thanks to both resident biologists and visiting researchers, we now know the flora and fauna of Costa Rica quite well. Located in Central America's narrowed southern isthmus, between Nicaragua and Panama, this country is completely tropical. Bounded by two large oceans, and averaging only around 160 km (100 miles) in width, Costa Rica receives ample rainfall over its entire area. As mentioned earlier, Costa Rica is home to about 10,600 species of vascular plants. If you take that number and divide it by the number of square kilometers ascribed to Costa Rica, you get a larger figure than you do for any other country on the planet! Colombia, Brazil, and China have many more species, but they support them over much larger territories. No other country in the world packs so many species into so small an area as does Costa Rica.25
Though half the area of Ohio—on a flat map—Costa Rica boasts twice as many species of mammals as all of North America north of Mexico. Likewise, this area of about 51,100 km2 (19,730 sq. mi.) has twice as many species of ferns as does all of North America north of Mexico. Costa Rica's bird count stands at 878 species, compared to about 850 in all of northern North America. We can amplify these species numbers with 175 amphibians and 218 reptiles, to go along with 228 mammal species. With such numbers and a very cordial populace, it's easy to see why this little democracy has become so popular a destination for ecotourists.26
What is it about little Costa Rica that allows so many species of plants and animals to make it their home? Simply stated: a tropical setting with generous rainfall and high mountains. The driest part of the country, the northwestern province of Guanacaste, averages about one meter (39 inches) of rainfall each year. Falling from May to November, these rains support a deciduous woodland at lower elevations. Here, few trees retain their leaves over the hot, five-month-long dry season. At higher elevations (above about 800 m / 2,625 ft.) temperatures are cooler, evapotranspiration is diminished, and partly evergreen forests are the original land cover. With higher rainfall and shorter dry seasons, evergreen broad-leaved forests originally covered all the rest of Costa Rica, except at the highest elevations. On the Caribbean side of the country, rain falls virtually all year long, supporting swamp forests as well as lowland rain forest. However, these many lowland forests are only one factor accounting for Costa Rica's high species numbers.
Costa Rica is a mountainous country. Its volcanoes and elevated highlands are part of “the ring of fire” bordering much of the Pacific Ocean. With the Caribbean plate overriding the Pacific margin, mountains are still rising and earthquakes are frequent. Beginning in the west, near Nicaragua, with a few smaller volcanoes, Costa Rica has a central chain of mountains traversing its entire 480 km (300 mile) length. The Cordilleras of Guanacaste and Tilarán dominate the northwestern third of the country, with both active and older volcanoes. A central volcanic group forms the nation's midsection, dominated again by volcanoes old and new. Finally, in the eastern half of the country, we have a massive uplift—the Cordillera de Talamanca—merging with the Chiriqui highlands of western Panama. Costa Rica's highest elevation, Cerro Chirripo, reaches 3,420 m (11,220 ft.) above sea level. All these cordilleras support tropical rain forest along their lower foothills, cooler cloud forests on their mid-elevation slopes, montane forests above about 2,200 m (7,280 ft.) elevation, elfin woodlands along exposed windy ridges, and grassy alpine paramo formations above about 3,200 m (10,500 ft.). These varied highlands, and the many valleys descending their slopes, provide the scaffolding for little Costa Rica's spectacular biological diversity. As we've noted, cool, moist montane cloud forests are especially rich in epiphytes. These are mostly smaller plants growing on the trunks and branches of larger trees; little animals also live in these tree-top communities.
Mountains make a difference. Of Costa Rica's 878 bird species, 160 are found only at higher elevations. As we noted in the fern studies, many plants and animals are restricted to the cooler highlands, and some of these are found nowhere else in the world. Of Costa Rica's birds, forty-seven are unique to the country. Many other species, of course, are wider ranging, including migratory birds that spend our cold winters in tropical climes. Over time, Central America's narrow isthmus has served as a bridge between two grand biotas, having been enriched from both the north and the south. On Costa Rica's highest mountains, we find plants characteristic of northern climates and also the high Andes; these are plants that cannot compete in warmer, lower elevation vegetation. Oaks dominate the cooler high-montane forests; they are a northern element in this southern flora. (Oaks are diverse in the United States and Mexico, diminishing in numbers in southern Central America, with only one species reaching northern South America.)
To summarize, high mountains with varied and dissected topography, together with a tropical setting receiving ample rainfall with differing local seasonality, help Costa Rica support so many species. The separation of related species—both plant and animal—by “altitudinal stratification” is an especially important factor supporting diversity in this little republic. These same factors escalate as we move south into the Andes.
THE ANDES MOUNTAINS OF SOUTH AMERICA
South America is also home to many “hotspots” of species richness. The eastern slopes of the Andes, bordering the Amazon and Orinoco drainage basins, are a huge area, with conditions very similar to those of Costa Rica. The nations of Colombia, Ecuador, Peru, and Bolivia are home to very high numbers of plants and animals. Here, a warm climate is coupled with frequent rain along the eastern slopes of the world's largest tropical mountain system. Colombia has the highest recorded bird count in the world, with around 1,800 species. Supporting Colombia's biodiversity is: (1) the exceptionally rainy Choco region of the northwest; (2) the high Andes, split into three parallel cordilleras; (3) open grasslands in the Orinoco basin; and (4) the Amazonian flora in the southeast.
As in Costa Rica, higher elevations along the eastern flank of the Andes support cooler evergreen forests often shrouded in mist and fog. Prevailing winds force warm moist air upward along the mountainside; becoming cooler as it rises, this air can no longer hold as much moisture, resulting in frequent rain, drizzle, and misting. With humidity high for most all the year, branches and tree trunks become festooned with epiphytic mosses, leafy liverworts, ferns, orchids, aroids, and bromeliads. “Tank bromeliads” are especially significant in enhancing biodiversity. Their broad clasping leaf-bases hold little pools of water high in the canopy. Here, far above the forest floor, animals can find a drink of water, and smaller aquatic creatures can live out their entire lives. Edward Wilson declared epiphytes “biodiversity multipliers.” Not only do these plants add their own numbers, but by providing special homes for little animals they further elevate species richness.27 That's the good news; the bad news is that the weight of epiphytes often causes branches to break, resulting in trees considerably shorter than those in lowland rain forests. Nevertheless, cloud forest may be the richest biome of all, supporting more species per hectare than even lowland tropical rainforest.
We mentioned the high species numbers in Colombia, Ecuador, and Peru, but the complex Andean highlands have been well collected only along major roads and near towns. Also, much of our knowledge of this region comes from isolated nature reserves and the research stations they support. One of the largest nature reserves along the eastern flank of the Andes is Manu National Park in southeastern Peru. Overall, the park covers about 1,532,806 km2 and ranges from 300 to 4,000 meters in elevation. Rainfall is sufficient to support evergreen forests throughout all but the highest elevations; the dry season lasts about three months. Overall, higher plants probably number around 6,000 species.28 These numbers include the tropical lowlands, cooler montane forests, and high paramo formations. The number of bird species recorded over this same altitudinal range in 2006 was 1,005, the highest number for any preserve of similar area on Earth. The number of mammal species recorded at Manu is 222, including 92 bat species, 58 rodents, 21 carnivores, and 14 primates. Significantly, this survey found not a single new species of birds, but it did include at least twelve new mammal species.29 This is a clear example of how much better we know the birds than we know the more secretive mammals. And that's not all: nineteen species of murid rodents were found to be carrying fifteen species of lice, three of which had never been seen before!30
Because of the difficulties in traversing Manu's full altitudinal range, most surveys have been limited to lower elevations along the river near the Pakitza research station. Here, 1,300 species of butterflies have been captured and identified (as of 1995).31 Similar studies found 498 species of spiders in 33 families and 136 species of dragonflies and damselflies. Amphibians and reptiles numbered 128 species, including one crocodilian and 67 frog species. All these numbers were based on short-term studies. Longer term studies at another site in Peru resulted in finding 113 species of amphibians and 118 species of reptiles, indicating that there are many more species to be found at Manu.32 All told, the moist eastern slopes of the tropical Andes probably support the richest repositories of biological diversity in the world.
OTHER MOIST-TROPICAL HOTSPOTS AROUND THE GLOBE
As we've noted, locations with high concentrations of species diversity can be characterized by only a few fundamental variables. These factors are “high energy input with constant water supply and extraordinarily high spatiotopographic complexity.”33 Little Costa Rica and the eastern slopes of the Andes have this combination of tropical warmth, ample rainfall, and complex mountainous terrain. Other parts of the world are similarly blessed, especially in Southeast Asia.
The island of Borneo (Sabah) is famous for being home to unusually high numbers of plants and animals. Kinabalu Park supports between 5,000 and 6,000 species of vascular plants. This same small area of 737 km2 is home to 326 species of birds and more than 700 species of orchids. In these forests one can find many species of vining insect-digesting pitcher plants. Other nearby islands, such as Indonesia, the Celebes, and New Guinea, are also extremely rich in species numbers and in endemic species. Of the twenty-five hotspots designated by Myers and Mittermeier, four are located in southeastern Asia. The large Indo-Burma hotspot ranges from the southern slopes of the Himalayas and easternmost India, through Myanmar to Vietnam.34 The Sundaland hotspot ranges from the southern half of the Malay peninsula to the islands of Java and Borneo.35 The Wallacea hotspot includes the eastern islands of Indonesia, the Celebes, and islands west of New Guinea.36 The Philippines hotspot is restricted to the islands of this archipelago.37
Important tropical wet-montane biotas are also found in Mt. Cameroon of western Africa, and on the higher mountains of East Africa's rift system, especially along the Uganda-Congo border where the mountain gorilla survives. In Asia, moist highland forests are found along the southern flanks of the Himalayas, from Nepal through Burma, Thailand, Laos, Vietnam, and southernmost China. Species-rich montane hotspots are also found in the Western Ghats of India and in Sri Lanka. While New Guinea supports one of world's most unusual faunas, it is less threatened by human expansion.
SPECIES RICHNESS IN SEASONALLY DRY OR COLD FORESTS
Deciduous broadleaf forests of both the temperate and tropical zones also contribute to our planet's enormous species count. Despite severe winters, temperate broadleaf forests are important regions of biodiversity. We've already noted that the Caucasus Mountains have been designated a hotspot, and China's southwest mountains support high species richness. Nevertheless, some of the world's most impressive temperate forests are not especially rich in species. The Giant Redwood forests of northwestern California, and the coniferous forests of the Pacific Northwest, dominated by Douglas fir and western hemlock, are awesome in their height and impressive in the density of tree trunks. No other forests in the world have so many tall slender trunks packed so closely together. Yet these grand forests are poor in species. No matter where they are found, forests dominated by the cone-bearing Gymnosperms do not support high species numbers. Perhaps this is a consequence of the chemicals with which they protect themselves. Conifer leaves are less subject to insect attack than are the leaves of flowering plants; fallen conifer logs rot much more slowly than do their flowering plant counterparts. Similarly, the great needle-leaf forests of the taiga are poor in species numbers, but here a long and cruel winter plays an important role. Moreover, just as the seasonally frigid temperate forests contribute to the world's overall biodiversity, seasonally dry tropical regions also enrich the world's species numbers.
Seasonally dry tropical woodlands and thorn bush are often rich in endemic species, and many have been designated hotspots on that account. While species numbers may not be as high as in the wet tropics, many local endemics live in these seasonally parched landscapes. The floras of the Sonoran Desert in the southwestern United States and northwestern Mexico, as well as the unusual flora of the nearby Baja California Peninsula are quite rich in species. In South America, the chaco vegetation of northeastern Brazil, and the cerrado and caatinga formations of Bolivia and southern Brazil are all rich in endemic species. The cerrado includes an estimated 10,000 vascular plant species, with many having become adapted to intense fires over the last five million years.38 In Africa, the Acacia-Commiphora thorn bush in eastern Ethiopia and Somalia, and the semi-desert floras of the Karoo and Namibian deserts are outstanding in harboring unusual plants and animals. The vegetation in these areas varies from open woodlands of short trees to scattered thorn scrub on rocky soils, with grassland covering flat, fire-prone sites.
We've discussed an unusually rich biome in subtropical, seasonally dry environments: the Mediterranean floras. Our gardens bear witness to the botanical wealth of these floras. The Mediterranean region itself, along with the nearby mountains of western Asia, were the original home to many of our favorite garden ornamentals. Daffodils, hyacinths, tulips, snapdragons, poppies, and oleander originated here. This same floristic area nourished western civilization with wheat, barley, lentils, peas, carrots, onions, lettuce, celery, parsley, artichoke, and many other vegetables and seasonings. Grapes, figs, olives, date-palms, and pomegranates also originated in this corner of the globe. Such a wealth of useful plants is a clear reflection of the generous diversity found within this flora. In fact, all five of the world's Mediterranean-type vegetation zones are included among Myers and Mittermeier's twenty-five biodiversity hotspots. Of these, one in particular has unusual plant diversity: it is the Cape region of southernmost Africa.39
SOUTH AFRICA'S CAPE FLORA
The flora at the southern end of Africa has puzzled botanists over many decades. Bordered by dry deserts and seasonally parched savanna woodlands, the Cape region supports a Mediterranean flora with winter rains and summer drought. The mountains are of old, eroded sandstone; soils are poor. Trees are few, and the landscape is mostly covered by scattered shrubs. Current estimates for the number of higher plants in the Cape floristic region stand at around 9,000 species, representing almost 1,000 genera and five endemic families.40 For a seasonally dry flora covering only 74,000 km2, these numbers are astounding. In a study to determine the global patterns and determinants of higher plant diversity, Holger Kreft and Walter Jetz subjected 1,032 regional floras to statistical analysis. Their worldwide analysis found that the Cape flora supports twice as many species as would be expected for a region with similar rainfall and temperature.41 Why might this corner of the world, with limited rainfall and modest elevational range, be so rich in species?
The Cape flora of South Africa is particularly rich in irises, lilies, aloes, and orchids, but poor in annual plants. Many small- and stiff-leaved shrubs dominate this landscape. Succulents like the stone flowers, thick-leaved aloes, and cactus-like euphorbias are also numerous. In contrast, animal species are few in number, with only about 288 bird and 127 mammal species. Interestingly, over a thousand plant species in this flora have seeds dispersed by ants! Carrying the seeds into their nests—and feeding on the fat-body attached to the seed—results in having seeds deposited in a moister environment, where rodents can't find them. Also, since ants can't travel very far, this may have promoted local speciation. Along the same line, many of the Cape's plant genera have closely related species living in nearby but slightly different environments, suggesting that these species are the products of local ecological speciation.42 Polyploidy appears to have been important in forming new species in a few lineages; the iris family with 295 species in the area has 27 polyploid species, nearly 10 percent.43
An interesting aspect of the Cape flora is that it has a number of genera with huge numbers of species. Erica of the blueberry family has 560 species in the Cape flora, while Mesembryanthemum numbered 700 species, before being split into segregate genera. Aspalanthus, a genus of leguminous shrubs, and Pelargonium, in the geranium family, each have more than 200 species in the region. Such species-rich genera are thought to have differentiated profusely over the last five million years. Using DNA analyses, Ben Warren and Julie Hawkins studied Muraltia of the polygala family, with over 100 species in the Cape region. They found that a majority of these species had formed over the last ten million years. That time period is in line with the origin of the Benguela Current, which brings cold waters from the southern ocean northward, and has produced drier conditions along Africa's southwestern coast. A diversification of Muraltia species over this time period may have been in response to that current's effects.44 Perhaps the drying effect took place gradually, allowing plant populations to adapt and differentiate. And since we are invoking the factor of time again, perhaps we need to look more carefully at its role in biodiversity.
BIODIVERSITY AND REGIONAL HISTORY
In the last chapter, we used history to explain tropical Africa's poor floristic richness, postulating that severe and widespread droughts have eroded Africa's plant diversity. Regarding the Cape flora of South Africa, we can use this same reasoning to opposite effect. This is not contradictory. If severe drought—or other calamity—can cause species loss, then surely lack-of-drought will keep speciation going and result in higher numbers. Clearly, the southernmost tip of Africa has not been subjected to unusual species extinction over recent geological time. Given time, plant species have continued to subdivide the land amongst themselves. Unlike the flora of Australia, South Africa did not shift its position from one climatic belt into another. Southernmost Africa does exhibit a longitudinal species-diversity gradient, with the highest species numbers in the western sector. This is correlated with more topographic diversity in the west, and winter rainfall may have been more reliable in this area.45
Likewise, it is clear that Costa Rica has not suffered severe species loss in recent geological time. Growing on the trunks and branches of larger trees, epiphytes require a climate of reliable moisture for survival. Comparing two other regional floras, we found that Costa Rica had about twice as many dicotyledonous families of flowering plants with epiphytes than either Indonesia or the Flora of Tropical West Africa, far larger areas. Because epiphytes have disseminules that must be able to germinate on tree branches, such plants disperse widely. Many of Costa Rica's epiphytes may have migrated northward from the Andes. No matter the original conditions, a flora hugely rich in epiphytes implies a long history of reliable rainfall.
ENDEMICS AND LINEAGES: WHAT CAN THEY TELL US?
Historical biogeography has been richly informed by endemic species and lineages. These are plants and animals found only in a specific geographical area. Some may be recent evolutionary innovations, little more than curiosities. But others are hangers-on, survivors from more ancient times. The many endemics of Hawaii don't have much to say about ancient biotas; the islands are young, and many new species have originated there. The numerous fruit fly species in Hawaii are a recent and explosive radiation. A different story is related by the platypus in Australia, the tuatara in New Zealand, and the lemurs of Madagascar; these tell of a long and isolated history for the land masses where they survive.46 Clearly, old islands can act as “shelters,” harboring ancient lineages, while young islands can serve as “cradles,” providing new opportunities for expansive speciation.
Mammals are especially informative in historical biogeography, and for two reasons. First of all, land mammals are not very good at getting across oceans. Compared to birds or the flowering plants, non-flying terrestrial mammals are tightly constrained in their travels. True, some mammals range widely over continents; the puma once roamed from Alaska to Patagonia, while leopards ranged from southern Africa to Southeast Asia. Nevertheless, land mammals cannot cross larger bodies of water. A second factor in their biogeographic importance is that mammals have a fine fossil record; we know a lot about the divergence times of their many families.47
Comparing the larger biogeographic regions, we find that 91 percent of Australia's mammal families are found nowhere else. South America ranks second, with 47 percent of its families endemic. Africa comes in at 36 percent, followed by the northern Holarctic Realm with 16 percent and the Oriental Realm with 13 percent. Looking more closely at mammalian geography, we must keep in mind that present geographic distributions can be misleading. Today, horses, zebras, and wild asses are only found in Africa and Eurasia. However, the fossil record makes clear that the ancestors of horses originated and diversified over many millions of years in North America. They spread into the Old World only later, where they are found today. Horses did survive into modern times in North America, only to go extinct after humans improved their hunting technology, around 12,000 years ago.
A fine example of an informative mammalian lineage is our own, the primates. Fossils of the earliest primates are from North America, about sixty million years ago. None of these ancient insectivores are alive today. A few other isolated and early branches of the primate tree do survive. The bush baby of Africa, the slow loris in Southeast Asia, and the aye-aye of Madagascar belong to these early primate branches. These are all small arboreal animals that are active only at night. The lemurs of Madagascar are another early branch; they represent a clearly defined larger element within the primates. Lemur-like animals are found as fossils in Europe from around fifty million years ago. Isolated from the rest of the world on their island home, lemurs retain many primitive primate features, including a more pointed, dog-like face. New World monkeys are considered more primitive than Old World monkeys because they've got more teeth in their mouths, and most lack the three-pigment color vision that characterizes Old World monkeys and ourselves. (More teeth are an ancient trait, better color vision is a modern trait.)
So there we have it. A variety of little primate oddballs in Africa and the Oriental realms. Lemurs confined to Madagascar, New World monkeys confined to tropical America, and the more modern Old World monkeys ranging from Africa and southern Europe all the way to eastern Asia and Wallace's Line. There's a pattern here. An important early phase of primate evolution survives only on the island of Madagascar, though similar animals once lived in Europe and Asia. New World monkeys survive only in the Americas, while the most modern primates are found over the largest contiguous area: Africa and southern Eurasia. Then there's Australia, with no primates at all, either today or in the fossil record. What do these broad geographic patterns suggest?
Lemurs represent an important stage in early primate evolution. They lived over a wide range in earlier times but are confined to Madagascar today. Similarly, New World monkeys are not the latest models in primate evolution, and they are no longer found in the Old World where their ancestors once thrived. The only reasonable conclusion is that—isolated on their island homes, whether Madagascar or South America—these animals were not confronted by the more advanced members of their own lineage, or predation by more modern carnivores. They have survived where they live by virtue of their isolation. And the even more ancient lineages represented today by lorises and bush babies? They survive in small numbers in deep forests, active only in the dark of night. The message of primate evolution is this: our world appears to have become increasingly competitive. Unless you are isolated from the action—or find a secure little niche—you may be vulnerable to extinction, as newer, higher-powered models take over the landscape. (We will return to the subject of “competitive escalation” in chapter 8.)

Having briefly reviewed the geography of species richness and patterns of diversity, we need to consider another fundamental ecological question. How is it that biological communities can sustain so many different species in the same place at the same time? Why haven't superior competitors taken over the landscapes for themselves? Why does so much of the world remain green, when there are literally millions of herbivores ready to devour all the greenery? How is it that complex ecosystems remain so complex?