“Every year I tell my students in my first lecture that at least half of what I am about to teach them will eventually be shown to be wrong. The trouble is I do not know which half. The future is a rough taskmaster. Nevertheless, a herd instinct often grips the imaginations of scientists. Like lemmings, we are prone to charge over cliffs when a large enough pack of us moves in that direction.”
— Michael Gershon, in The Second Brain, p. 34.
The first organizing principle underlying human movement and posture is our existence in a gravitational field. Imagine its absence in a spacecraft, where astronauts float unless they are strapped in place, and where outside the vessel little backpack rockets propel them from one work site to another. To get exercise, which is crucial for preventing loss of bone calcium on long voyages, they must work out on machines bolted to the floor. They can’t do the three things that most of us depend on: walking, running, and lifting. If they tried to partner up for workouts, all they could do is jerk one another back and forth. And even hatha yoga postures would be valueless; they would involve little more than relaxing and squirming around.
Back on earth, it is helpful to keep recalling how the force of gravity dominates our practice of hatha yoga. We tend to overlook it, forgetting that it keeps us grounded in the most literal possible sense. When we lift up into the cobra, the locust, or the bow postures, we lift parts of the body away from the ground against the force of gravity. In the shoulderstand the force of gravity holds the shoulders against the floor. In a standing posture we would collapse if we did not either keep antigravity muscles active or lock joints to remain erect. And even lying supine, without the need either to balance or to activate the antigravity muscles, we make use of gravity in other creative ways, as when we grasp our knees, pull them toward the chest, roll from side to side, and allow our body weight to massage the back muscles against the floor.
Keeping in mind that the earth’s gravitational field influences every movement we make, we’ll turn our attention in the rest of this chapter to the mechanisms that make movement and posture possible. First we’ll look at how the skeletal muscles move the body, then we’ll discuss the way the nervous system controls the operation of the skeletal muscles, and then we’ll examine how connective tissues restrict movement. If we understand how these three function together within the field of gravity, we can begin to understand some of the principles underlying hatha yoga. Finally, we’ll put it all together in a discussion of three postures. We’ll begin with the role of skeletal muscles.
THE NEURO-MUSCULOSKELETAL SYSTEM
To any informed observer, it is plain that the musculoskeletal system executes all our acts of will, expresses our conscious and unconscious habits, breathes air into the lungs, articulates our oral expression of words, and implements all generally recognized forms of nonverbal expression and communication. And in the practice of hatha yoga, it is plainly the musculoskeletal system that enables us to achieve external balance, to twist, bend, turn upside down, to be still or active, and to accomplish all cleansing and breathing exercises. Nevertheless, we are subtly deceived if we think that is the end of the story. Just as we see munchkins sing and dance in The Wizard of Oz and do not learn that they are not autonomous until the end of the story, we’ll find that muscles, like munchkins, do not operate in isolation. And just as Dorothy found that the wizard kept a tether on everything going on in his realm, so we’ll see that the nervous system keeps an absolute rein on the musculoskeletal system. The two systems combined form a neuro-musculoskeletal system that unifies all aspects of our actions and activities.
To illustrate how the nervous system manages posture, let’s say you are standing and decide to sit. First your nervous system commands the flexor muscles (muscles that fold the limbs and bend the spine forward) to pull the upper part of the trunk forward and to initiate bending at the hips, knees, and ankles. A bare moment after you initiate that movement, gravity takes center stage and starts to pull you toward the sitting position. And at the same time—accompanying the action of gravity—the nervous system commands the extensor muscles (those that resist folding the limbs) to counteract gravity and keep you from falling in a heap. Finally, as soon as you are settled in a secure seated position, the nervous system permits the extensor muscles and the body as a whole to relax.
The musculoskeletal system does more than move the body, it also serves as a movable container for the internal organs. Just as a robot houses and protects its hidden supporting elements (power plant, integrated circuits, programmable computers, self-repairing components, and enough fuel to function for a reasonable length of time), so does the musculoskeletal system house and protect the delicate internal organs. Hatha yoga postures teach us to control both the muscles that operate the extremities and the muscles that form the container.
SKELETAL MUSCLE
The term “muscle” technically includes both its central fleshy part, the belly of the muscle, and its tendons. The belly of a muscle is composed of individual muscle fibers (muscle cells) which are surrounded by connective tissue fibers that run into a tendon. The tendon in turn connects the belly of the muscle to a bone.
Under ordinary circumstances muscle cells contract, or shorten, only because nerve impulses signal them to do so. When many nerve impulses per second travel to most of the individual fibers in a muscle, it pulls strongly on the tendon; if only a few nerve impulses per second travel to a smaller population of fibers within the muscle, it pulls weakly on the tendon; and if nerve impulses are totally absent the muscle is totally relaxed.
[Technical note: One of the most persistent misconceptions doggedly surviving in the biomedical community is that all muscles, even those at rest, always keep receiving at least some nerve impulses. Fifty years of electromyography with fine-wire needle electrodes is at odds with this belief, documenting from the 1950s on that it’s not necessarily true, and that with biofeedback training we can learn to relax most of our skeletal muscles completely.]
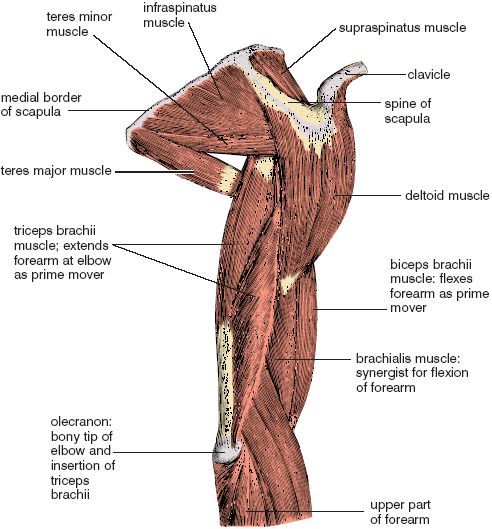
A muscle usually operates on a movable joint such as a hinge or a ball and socket, and when a muscle is stimulated to contract by the nervous system, the resulting tension is imparted to the bones on both sides of the fulcrum of the joint. In the case of a hinge such as the elbow that opens to about l80°, any muscle situated on the face of the hinge that can close will decrease the angle between the two bones, and any muscle situated on the back side of the hinge will open it up from a closed or partially closed position. For example, the biceps brachii muscle lies on the inside of the hinge, so it acts to flex the forearm (by definition, the segment of the upper extremity between the wrist and the elbow), pulling the hand toward the shoulder. The triceps brachii is situated on the back side of the arm (the segment of the upper extremity between the elbow and the shoulder) on the outside of the hinge, so it acts to extend the elbow, or unfold the hinge (fig. 1.1).
ORIGINS AND INSERTIONS
We use the words “origin” and “insertion” to indicate where muscles are attached to bones in relation to the most common movement at a joint. The origin of a muscle is on the bone that is relatively (or usually) stationary, and the insertion of a muscle is on the bone that is most generally moved. Flexion of the elbow is again a good example. Since ordinarily the arm is fixed and the forearm is moved, at least in relative terms, we say that the biceps brachii and triceps brachii take origin from the arm and shoulder, and that they insert on the forearm (fig. 1.1).
The origins and insertions of a muscle can be functionally reversed. When the latissimus dorsi muscle (figs. 8.9–10) pulls the arm down and back in a swimming stroke, its textbook origin is from the lower back and pelvis, and its insertion is on the humerus in the arm. But when we do a chin-up the arm is the relatively stable origin, and the lower back and pelvis become the insertion for lifting the body as a whole. In the coming chapters we’ll see many examples of how working origins and insertions are reversed.
AGONIST AND ANTAGONIST MUSCLES
The muscles surrounding a joint act cooperatively, but one of them—the agonist—ordinarily serves as the prime mover, assisted in its role by functionally related muscles called synergists. While the agonist and its synergists are acting on one side of the joint, muscles on the opposite side act as antagonists. As suggested by the name, antagonists monitor, smooth, and even retard the movement in question. For example, when the biceps brachii and the brachialis in the arm (the agonist and one of its synergists) shorten to flex the elbow, the triceps brachii (on the opposite side of the arm) resists flexion antagonistically while incidentally holding the joint surfaces in correct apposition (fig. 1.1).
Muscles also act in relation to the force of gravity. In the lower extremities extensor muscles act as antigravity muscles to keep you upright and resist crumpling to the floor. Examples: the quadriceps femoris muscle (figs. 1.2, 3.9, and 8.11) on the front of the thigh (the segment of the lower extremity between the hip joint and the knee joint) extends the knee joint as you step onto a platform, and the calf muscles extend the ankles as you lift your heels to reach an object on a high shelf. Flexor muscles are antagonists to the extensors. They can act in two ways. They often aid gravity, as when you settle into a standing forward bend and then pull yourself down more insistently with your hip flexors—the iliopsoas muscles (figs. 2.8, 3.7, 3.9, and 8.13). But they also act to oppose gravity: if you want to run in place the iliopsoas muscle complex flexes the hip joint, lifting the thigh and drawing the knee toward the chest; and if you want to kick yourself in the buttocks the hamstrings (fig. 3.8, 3.10, 8.10, and 8.12) flex the knee, pulling the leg (the segment of the lower extremity between the knee and the ankle) toward the thigh. Even so, the flexor muscles in the lower extremities are not classified as antigravity muscles, because under ordinary postural circumstances they are antagonists to the muscles that are supporting the body weight as a whole.
For the upper extremities the situation is different, because unless you are doing something unusual like taking a walk in a handstand with slightly bent elbows (which necessitates a strong commitment from the triceps brachii muscles), the extensor muscles do not support the weight of the body. In most practical circumstances, it is likely to be the flexors rather than extensors that act as antigravity muscles in the upper extremities, as when you flex an elbow to lift a package or complete a chin-up.
[Technical note: Throughout this book, in order to keep terminology simple and yet precise, I’ll stick with strict anatomical definitions of arm, forearm, thigh, and leg, which means never using ambiguous terms such as “upper arm,” “lower arm,” “upper leg,” and “lower leg.” The same goes for the careless use of the term “arm” to encompass an undetermined portion of the upper extremity and the careless use of the term “leg” to encompass an undetermined portion of the lower extremity.]
Figure 1.1. View of the right scapula, arm, and upper part of the forearm from behind and the side (from Sappey; see “Acknowledgements” for discussion of credits regarding drawings, illustrations, and other visual materials).
[And another technical note: Just to avoid confusion, I’ll not use the word flex except in regard to the opposite of extend. Everyone knows what a first grader means by saying “look at me flex my muscles,” but beyond this childhood expression, it can lead to ambiguity. For example, having someone “flex their biceps” results in flexion of the forearm, but “flexing” the gluteal muscles—the “gluts”—results not in flexion but in extension of the hips. For describing yoga postures it’s better just to avoid the usage altogether.]
[And one more: Until getting used to terminology for movements of body parts, it is often a puzzle whether it’s better to refer to moving a joint through some range of motion, or to moving the body part on the far side of the joint. For example, the choice might be between saying: extend the knee joint versus extend the leg, abduct the hip joint versus abduct the thigh, flex the ankle joint versus flex the foot, extend the elbow joint versus extend the forearm, or hyperextend the wrist joint versus hyperextend the hand. Even though the two usages are roughly equivalent, the context usually makes one or the other seem more sensible. For example, sometimes we refer specifically to the joint, as in “flex the wrist.” In that case, saying “flex the hand” would be ambiguous because it could mean any one of three things: making a fist, flexing the wrist, or both in combination. On the other hand, referring to the body part is often more self-explanatory, as in “flex the arm forward 90°.” Although the alternative—“flex the shoulder joint 90°”—isn’t nonsensical, it’s a little arcane for the non-professional.]
CONCENTRIC SHORTENING AND ECCENTRIC LENGTHENING
To understand how the musculoskeletal system operates in hatha yoga we must look at how individual muscles contribute to whole-body activity. The simplest situation, concentric contraction, or “concentric shortening,” is one in which muscle fibers are stimulated by nerve impulses and the entire muscle responds by shortening, as when the biceps brachii muscle in the arm shortens concentrically to lift a book.
When we want to put the book down the picture is more complicated. We do not ordinarily drop an object we have just lifted—we set it down carefully by slowly extending the elbow, and we accomplish that by allowing the muscle as a whole to become longer while keeping some of its muscle fibers in a state of contraction. Whenever this happens—whenever a muscle increases in length under tension while resisting gravity—the movement is called “eccentric lengthening.”
We see concentric shortening and eccentric lengthening in most natural activities. When you walk up a flight of stairs, the muscles that are lifting you up are shortening concentrically; when you walk back down the stairs, the same muscles are lengthening eccentrically to control your descent. And when you haul yourself up a climbing rope hand over hand, muscles of the upper extremities shorten concentrically every time you pull yourself up; as you come back down, the same muscles lengthen eccentrically.
In hatha yoga we see concentric shortening and eccentric lengthening in hundreds of situations. The simplest is when a single muscle or muscle group opposes gravity, as when the back muscles shorten concentrically to lift the torso up from a standing forward bend. Then as you slowly lower back down into the bend, the back muscles resist the force of gravity that is pulling you forward, lengthening eccentrically to smooth your descent.
ISOTONIC AND ISOMETRIC ACTIVITY
Most readers are already familiar with the terms “isotonic” and “isometric.” Strictly speaking, the term isotonic refers solely to shortening of a muscle under a constant load, but this never happens in reality except in the case of vanishingly small ranges of motion. Over time, however, the term isotonic has become corrupted to apply generally to exercise that involves movement, usually under conditions of moderate or minimal resistance. Isometric exercise, on the other hand, refers to something more precise—holding still, often under conditions of substantial or maximum resistance. Raising and lowering a book repetitively is an isotonic exercise for the biceps brachii and its synergists, and holding it still, neither allowing it to fall nor raising it, is an isometric exercise for the same muscles. Most athletic endeavors involve isotonic exercise because they involve movement. Japanese sumo wrestling between equally matched, tightly gripped, and momentarily immobile opponents is one obvious exception. And isometric exercise is also exemplified by any and every hatha yoga posture which you are holding steadily with muscular effort.
RELAXATION, STRETCH, AND MOBILITY
If few or no nerve impulses are impinging on muscle fibers, the muscle tissue will be relaxed, as when you are in the corpse posture (fig. 1.14). But if a relaxed muscle is stretched, the situation becomes more complex. Working with a partner can make this plain. If you lie down and lift your hands straight overhead, and then ask someone to stretch you gently by pulling on your wrists, you will notice that you can easily go with the stretch provided you have good flexibility. But if your partner pulls too suddenly or if there is any appreciable pain, the nervous system will resist relaxation and keep the muscles tense; or at the least, you will sense them tightening up to resist the stretch. Finally, if you allow yourself to remain near your limit of passive but comfortable stretch for a while longer, you may feel the muscles relax again, allowing your partner to pull more insistently.
Many of these same responses are apparent if you set up similar conditions of stretching on your own, as when you place your hands overhead against a wall and stretch the underside of the arms. This is more demanding of your concentration than relaxing into someone else’s work, however, because you are concentrating on two tasks at the same time: creating the necessary conditions for the stretch, and relaxing into that effort. But the same rules apply. If you go too far and too quickly, pain inhibits lengthening, prevents relaxation, and spoils the work.
MUSCULAR ACTIVITY IN A LUNGING POSTURE
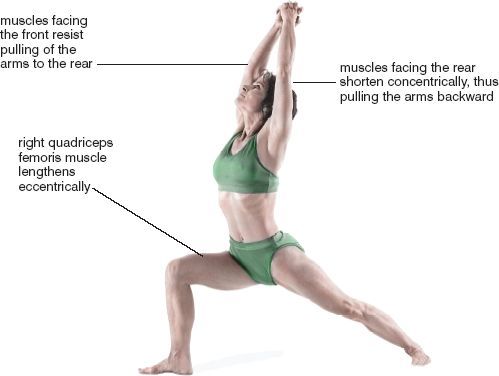
To discover for yourself how skeletal muscles operate in hatha yoga, try a warrior posture (warrior I) with the feet spread wide apart, the hands stretched overhead, and the palms together (figs. 1.2 and 7.20). Feel what happens as you slowly pull the arms to the rear and lower your weight. To pull the arms up and back, the muscles facing the rear have to shorten concentrically, while antagonist muscles facing the front passively resist the stretch and possibly completion of the posture. As you lower your weight the quadriceps femoris muscle on the front of the flexed thigh resists gravity and lengthens eccentrically. Finally, as you hold still in the posture, muscles throughout the body will be in a state of isometric contraction.
Several important principles of musculoskeletal activity cannot be addressed until we have considered the nervous system and the connective tissues in detail. For now, it is enough to realize that all muscular activity, whether it be contraction of individual cells, isotonic or isometric exercise, agonist or antagonist activity, concentric shortening, or eccentric lengthening, takes place strictly under the guidance of the nervous system.
We experience all—or at least everything pertaining to the material world—through the agency of specialized, irreplaceable cells called neurons, 100 billion of them in the brain alone, that channel information throughout the body and within the vast supporting cellular milieu of the central nervous system (the brain and spinal cord). This is all accomplished by only three kinds of neurons: sensory neurons, which carry the flow of sensation from the peripheral nervous system (by definition all parts of the nervous system excepting the brain and spinal cord) into the central nervous system and consciousness; motor neurons, which carry instructions from the brain and spinal cord into the peripheral nervous system, and from there to muscles and glands; and interneurons, or association neurons, which are interposed between the sensory neurons and the motor neurons, and which transmit our will and volition to the motor neurons. The sensory information is carried into the dorsal horn of the spinal cord by way of dorsal roots, and the motor information is carried out of the ventral horn of the spinal cord by way of ventral roots. The dorsal and ventral roots join to form mixed (motor and sensory) spinal nerves that in turn innervate structures throughout the body (figs. 1.3–9).
[Technical notes: Because this is a book correlating biomedical science with yoga, which many consider to be a science of mind, a few comments are required on a subject of perennial, although possibly overworked, philosophical interest—the nature of mind vis-a-vis the nervous system. Speaking for neuroscientists, I think I can say that most of us accept as axiomatic that neurons are collectively responsible for all of our thinking, cognition, emotions, and other activities of mind, and that the totality of mind is inherent in the nervous system. But I also have to say as a practicing yogi that according to that tradition, the principle of mind is separate from and more subtle than the nervous system, and is considered to be a life principle that extends even beyond the body.]
[How and whether these questions become resolved in the third millennium is anyone’s guess. They are topics that are not usually taken seriously by working scientists, who usually consider it a waste of time to ponder non-testable propositions, which are by definition propositions that cannot possibly be proven wrong. Such statements abound in new age commentaries, and are a source of mild embarrassment to those of us who are trying to examine older traditions using techniques of modern science. This says nothing about the accuracy of such proposals. It may be true, for example, that “life cannot continue in the absence of prana.” The problem is that short of developing a definition and assay for prana, such a statement can not be tested—it can only be accepted, denied, or argued ad infinitum.]
[This approach to experimentation and observation doesn’t require a lot of brilliance. It simply stipulates that you must always ask yourself if the nature and content of a statement make it potentially refutable with an experimental approach. If it’s not, you will be accurate 90% of the time if you conclude that the idea is spurious, even though it may sound inviting or may even appear self-evident, as did the chemical theory of phlogiston in the mid-l8th century. To give the benefit of the doubt to the purveyors of such statements, it’s rare that they are outright fabrications. On the other hand, one should always keep in mind that all of us (including scientists) have a huge capacity for deceiving ourselves when it comes to defending our ideas and innovations. The problem is that it’s often impossible to distinguish fantasy, wishful thinking, mild exaggeration, and imprecise language from out-and-out fraud. What to do? In the end it’s a waste of time to make a career of ferreting out errors—one can’t get rid of bad ideas by pointing them out. On the other hand, if we turn our attention to propositions that can be tested, the creative attention this requires sometimes brings inspiration and better ideas, which in turn disposes of bad ideas by displacing them. Lavoisier discredited the theory of phlogiston by pointing to brilliant experiments (many of them carried out by others), not by crafting cunning arguments.]
[One last concern: if your complaint is that you can’t understand a particular concept and do not feel competent to criticize it, don’t assume that the problem is your own lack of intelligence or scientific background. More than likely, the idea wasn’t presented in a straightforward manner, and it usually happens that this masks one or more fatal flaws in the putative reasoning. One dependable test of a concept is whether you can convincingly explain it, along with the mechanics of how it operates, to a third party. If you find yourself getting your explanation garbled, or if your listener does not comprehend your arguement or is unpersuaded, please examine and research the idea more critically, and if it still does not pass muster, either discard it or put it on the back burner. I invite the reader to hold me to these standards. To honor them, I’ll limit inquiries to what we can appraise and discuss in the realm of modern biomedical science, and to refine and improve my presentation, I ask for your written input and cordial criticism.]
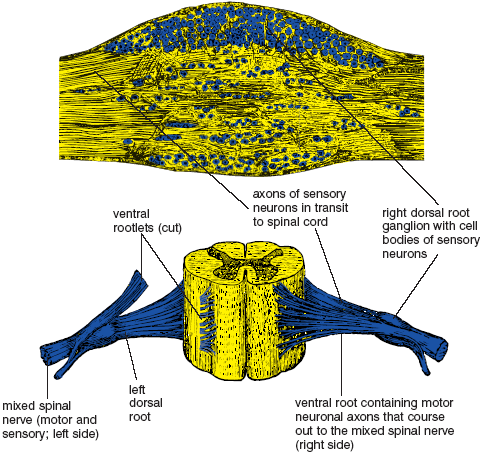
Figure 1.3. Microscopic section of dorsal root ganglion (above), and three-dimensional view of the first lumbar segment (L1) of the spinal cord, showing paired dorsal and ventral roots and mixed (motor and sensory) spinal nerves (from Quain).
Returning to our immediate concerns, it is plain that neurons channel our mindful intentions to the muscles, but we still need working definitions for will and volition. In this book I’m arbitrarily defining will as the decision-making process associated with mind, and I’m defining volition as the actual initiation of the on and off commands from the cerebral cortex and other regions of the central nervous system that are responsible for commanding our actions. So “will” is a black box, the contents of which are still largely unknown and at best marginally accessible to experimentation. The nature and content of volition, by contrast, can be explored with established methods of neuroscience.
NEURONS
The neuron is the basic structural and functional unit of the nervous system. Although there are other cell types in the nervous system, namely the neuroglia, or “nerve glue cells,” which outnumber neurons 10:1, these supporting cells do not appear, as do the neurons, to be in the business of transmitting information from place to place. So the neuron is our main interest. It has several components: a nucleated cell body that supports growth and development, and cellular extensions, or processes, some of them very long, that receive and transmit information. The cellular processes are of two types: dendrites and axons. Picture an octopus hooked on a fishing line. Its eight arms are the dendrites, and the fishing line is the axon. A typical motor neuron contains many dendrites that branch off the cell body. Its single axon—the fishing line—may extend anywhere from a fraction of an inch away from the cell body to four feet in the case of a motor neuron whose cell body is in the spinal cord and whose terminal ends in a muscle of the foot, or even fifteen feet long in the case of similar neurons in a giraffe. The axon may have branches that come off the main trunk of the axon near the cell body (axon collaterals), and all branches, including the main trunk, subdivide profusely as they near their targets.
Dendrites are specialized to receive information from the environment or from other neurons, and an axon transmits information in the form of nerve impulses to some other site in the body. Dendrites of sensory neurons are in the skin, joints, muscles, and internal organs; their cell bodies are in dorsal root ganglia, which are located alongside the spine, and their axons carry sensory information into the spinal cord (figs. 1.3–9). Dendrites of motor neurons are located in the central nervous system, and axons of motor neurons fan out from there (in peripheral nerves) to innervate muscle cells and glands throughout the body. Between the sensory and motor neurons are the association neurons, or interneurons, whose dendrites receive information from sensory neurons and whose axons contact other interneurons or motor neurons that innervate muscles (fig. 1.4). As a class, the interneurons comprise most of the neurons within the brain and spinal cord, including secondary and tertiary linking neurons that relay sensory signals to the cerebrum, projection neurons that relay motor signals from the cerebrum and cerebellum to intermediary neurons that eventually contact motor neurons of the spinal cord, and commissural neurons that connect the right and left cerebral hemispheres—that is, the “right brain” and the “left brain.”
Interneurons put it all together. You sense and ultimately do, and between sensing and doing are the integrating activities of the interneurons. It’s true, as the first-grade reader suggests, that you can think and do, but more often you sense, think, and do.
To operate the entire organism, neurons form networks and chains that contact and influence one another at sites called “synapses.” Synaptic terminals of axons at such sites release chemical transmitter substances that affect the dendrite of the next neuron in the chain (fig. 1.4). The first neuron is the pre-synaptic neuron, and the neuron affected is the post-synaptic neuron. The pre-synaptic axon terminal transmits to the post-synaptic dendrite—not the other way around; it’s a one-way street.
Two types of transmitter substances are released at the synapse: one facilitates (speeds up) the activity of the post-synaptic neuron; the other inhibits (slows down) the activity of the post-synaptic neuron. Thousands of axon terminals may synapse on the dendrites of one post-synaptic neuron, and the level of activity of the recipient neuron depends on its pre-synaptic input. More facilitation yields more activity in the post-synaptic neuron in the form of increasing numbers of nerve impulses that travel down its axon; more inhibition yields diminished activity. For example, the pre-synaptic input of association neurons to motor neurons either facilitates the activity of motor neurons, causing them to fire more nerve impulses per second to skeletal muscles, or it inhibits their activity, causing them to fire fewer nerve impulses per second. The peacock posture (fig. 3.23d) requires maximum facilitation and diminished inhibition of the motor neurons that innervate the abdominal muscles, deep back muscles, muscles that stabilize the scapulae, and flexors of the forearms. On the other hand, muscular relaxation in the corpse posture (fig. 1.14) requires reduced facilitation and possibly increased inhibition of motor neurons throughout the central nervous system (see fig. 10.1 for a summary of possible mechanisms of muscular relaxation).

Figure 1.4. Cross-section of the fifth lumbar segment (L5) of the spinal cord, with sensory input from a joint receptor, a generic interneuron, and motor output to a skeletal muscle cell. The small arrows indicate the direction of nerve impulses and pre- to post-synaptic interneuronal relationships. The long heavy arrows indicate the locations of the generic interneuron in the spinal cord dorsal horn and of the motor neuron in the spinal cord ventral horn.
VOLITION: THE PATHWAYS TO ACTIVE VOLUNTARY MOVEMENT
Exercising our volition to create active voluntary movement involves dozens of well-known circuits of association neurons whose dendrites and cell bodies are in the cerebrum, cerebellum, and other portions of the brain, and whose axons terminate on motor neurons. A small but important subset of projection neurons, the subset whose cell bodies are located in the cerebral cortex and whose axons terminate on motor neurons in the spinal cord, are known as “upper motor neurons” because they are important in controlling willed activity. These are differentiated from the main class of motor neurons, the “lower motor neurons,” whose cell bodies are located in the spinal cord. Collectively, the lower motor neurons are called the “final common pathway” because it is their axons that directly innervate skeletal muscles. In common parlance, if someone refers simply to “motor neurons,’ they are invariably thinking of lower motor neurons (fig. 1.5).
LOWER MOTOR NEURON PARALYSIS: FLACCID PARALYSIS
The best way to understand how the motor pathways of the nervous system operate is to examine the classic neurological syndromes that result from illnesses, or from injuries that have an impact on some aspect of motor function. We’ll start with one of the most famous: poliomyelitis, commonly known as polio, which destroys lower motor neurons. Anyone who grew up in the 1940s and early 1950s will remember the dread of this disease. And then a miracle—the Salk vaccine—came in 1954, putting an end to the fear.
Poliomyelitis can be devastating because it destroys the lower motor neurons and deprives the muscles of nerve impulses from the spinal cord, and this results in muscular paralysis. Our power of volition in the cerebral cortex has been disconnected from the pathway of action out of the spinal cord because the final common pathway has been destroyed. In its most extreme form the resulting paralysis causes muscles to become completely flaccid, and this accounts for its medical name: flaccid paralysis. The same thing happens in a less global fashion when a peripheral nerve is severed or crushed. Destruction of the lower motor neurons or their axons at any site in the spinal cord or peripheral nerves causes paralysis of all their muscular targets. Will, volition, and active voluntary movement are totally frustrated.
UPPER MOTOR NEURON PARALYSIS: SPASTIC PARALYSIS
When the upper motor neurons or their axons are destroyed as in an injury or stroke (the interruption of blood supply to the brain) that destroys the motor region of the cerebral cortex, we lose much of our voluntary control of the lower motor neurons, especially on the side opposite to the site of the injury. Our will can no longer be expressed actively and smoothly. The ultimate result of this, at least in severe cases in which a vascular mishap occurs at a site where the axons of other motor systems are interrupted along with those of the upper motor neurons, is not flaccid paralysis but spastic paralysis, in which the muscles are rigid and not easily controlled. A semblance of motor function remains because other parts of the nervous system, parts that have been spared injury, also send axon terminals to the lower motor neurons and affect motor function. The problem is that these supplemental sources of input cannot be controlled accurately, and some of them facilitate the lower motor neurons to such an extent that skeletal muscles are driven into strong and uncontrolled states of contraction. Although most of the time the condition does not result in total dysfunction, severe spastic paralysis is only mildly less devastating than flaccid paralysis; some active voluntary movements are possible, but they are poorly coordinated, especially those that make use of the distal muscles of the extremities (fig. 1.6).
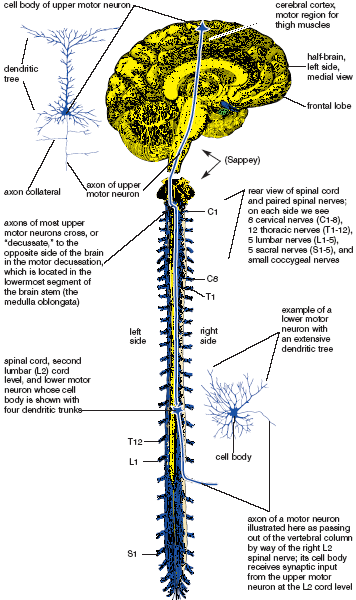
Figure 1.5. Upper and lower motor neurons. The cell body of an upper motor neuron is shown above in the left cerebral cortex, and its target—the cell body of a motor neuron whose axon is destined to innervate the right quadriceps femoris muscle—is on the right side of the spinal cord.
SPINAL CORD INJURIES
If the entire spinal cord is severed or severely damaged at some specific level, there are two main problems. First, sensory information that comes into the spinal cord from below the level of the injury cannot get to the cerebral cortex and thereby to conscious awareness. The patient is not aware of touch, pressure, pain, or temperature from the affected region of the body. Second, motor commands from the brain cannot get to the lower motor neurons that are located below the injury. Spinal cord injuries at different levels illustrate these conditions: a spinal cord transection in the thoracic region would result in paraplegia—paralysis and loss of sensation in the lower extremities; and a spinal cord transection in the lower part of the neck would cause quadriplegia—paralysis and loss of sensation from the neck down, including all four extremities (fig. 2.12). Injuries such as these are usually the result of either automobile or sports accidents.
So far our discussion has focused on neuronal connections from the top down—from our intention, to the cerebral cortex, to upper motor neurons, lower motor neurons, and skeletal muscles. But there is something else to consider, something much more primitive and elemental in the nervous system that bypasses our conscious choices: reflexes, or unconscious motor responses to sensory stimuli. In this context reflexes have nothing at all to do with the lightning-fast reactions (“fast reflexes”) that are needed for expertise in video games or quick-draw artistry. These reactions refer to unconscious responses carried out at the spinal level.
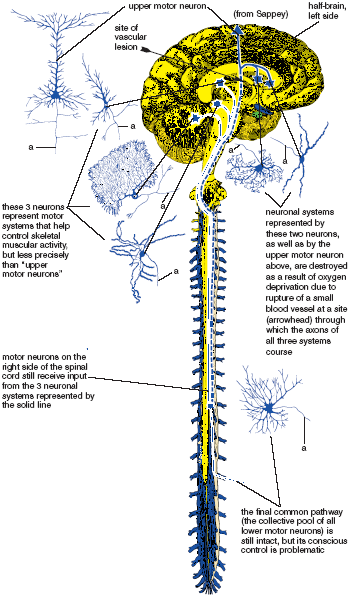
Figure 1.6. A hypothetical scheme illustrating how injury to a small region of the brain could interrupt pathways important for the precise control of skeletal muscular activity and cause spastic paralysis. The dotted line represents the systems that have been interrupted, and the solid line represents the remaining systems that cannot control muscular activity accurately by themselves. “a” = axon.
Reflexes are simple. That is why they are called reflexes. They always include four elements: a sensory neuron that receives a stimulus and that carries nerve impulses into the spinal cord, an integrating center within the spinal cord, a motor neuron that relays nerve impulses back out to a muscle, and the muscular response that completes the action. More explicitly, the sensory neurons carry nerve impulses from a muscle, tendon, ligament, joint, or the skin to an integrating center in the spinal cord. This integrating center might be as simple as one synapse between the sensory and motor neuron, or it might involve one or more interneurons. The motor neuron, in its turn, innervates muscle cells that complete the action. By definition, the reflex bypasses higher centers of consciousness. Awareness of the accompanying sensation gets to the cerebral cortex after the fact and only because it is carried there independently by other circuits. There are dozens of well-known reflexes. We’ll examine three, all of which are important in hatha yoga.
THE MYOTATIC STRETCH REFLEX
The myotatic stretch reflex, familiar to everyone as the “kneejerk,” is actually found throughout the body, but is especially active in antigravity muscles (fig. 1.7). You can test it in the thigh. Cross your knees so that one foot can bounce up and down freely, and then tap the patellar tendon just below the kneecap with the edge of your hand. Find just the right spot, and the big set of quadriceps femoris muscles on the front of the thigh will contract reflexly and cause the foot to fly up. You have to remain relaxed, however, because it is possible to override the reflex with a willed effort to hold the leg in place.
The receptors for the myotatic stretch reflex are located in the belly of the muscle, where the dendrites of sensory neurons are in contact with muscle spindles—specialized receptors barely large enough to be visible with the naked eye. Named for their shapes, each of these muscle spindles contains a spindle-shaped collection of specialized muscle fibers that are loaded with sensory receptors (fig. 1.7).
The reflex works this way: When you tap the patellar tendon to activate the reflex at the knee joint, the impact stretches muscle spindles in the quadriceps femoris muscle on the front of the thigh. This stretch is as fast as an eyeblink, but it nevertheless stimulates the specific sensory neurons whose dendrites end in the muscle spindles and whose axons terminate directly on motor neurons back in the spinal cord. Those axon terminals strongly facilitate the cell bodies of the motor neurons whose axon terminals stimulate the quadriceps femoris muscle, causing it to shorten and jerk the foot up. The myotatic stretch reflex is specific in that it feeds back only to the muscle in which the spindle is located.
As with all reflexes, this one takes place a fraction of a second before you are aware of it consciously. You feel it happen after the fact, after the reflex has already completed its circuit. And you notice the sensation consciously only because separate receptors for the modality of touch send messages to the cerebral cortex and thus into the conscious mind.
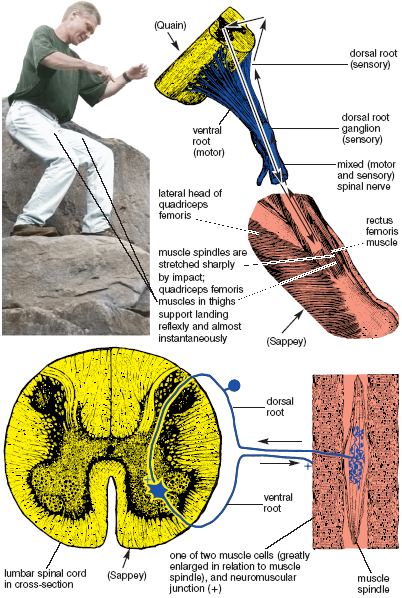
Figure 1.7. The myotatic stretch reflex. A 3-foot vertical jump momentarily stretches muscle spindles in all the extensor (anti-gravity) muscles of the lower extremities. The spindles then provide direct (monosynaptic) and almost immediate facilitatory input (+ in ventral horn of spinal cord) to extensor motor neurons, resulting in strong reflex contraction of the individual muscles.
You can feel the myotatic stretch reflex in operation in many sports in which your muscles absorb dynamic shocks. For example, when you are water skiing on rough water outside the wake of a boat, the muscle spindles in the knee extensors of the thighs are stretched by the impact of hitting each wave, and absorbing one bump after another would quickly collapse your posture were it not for the myotatic stretch reflex. Instead, what happens is that each impact activates the reflex for the quadriceps femoris muscles in a few milliseconds, thus stabilizing the body in an upright position. You can also feel the reflex when you attack moguls aggressively on a ski slope, run down the boulder field of a mountain (fig. 1.7), or simply jump off a chair onto the floor—any activity in which an impact shocks the muscle spindles. The reflex is therefore a major contributor to what we interpret as “strength” in our dynamic interactions with gravity. Athletes depend on it far more than most of them realize.
Stimulating myotatic stretch reflexes repetitively has another important effect: it shortens muscles and diminishes flexibility. We can see this most obviously in jogging, which only mildly engages the reflexes each time your front foot hits the ground, but engages them thousands of times in a halfhour. This can cause problems if taken to an extreme, and if you tend to be tight you should always do prolonged slow stretching after a run. On the other hand, if the muscles, tendons, and ligaments are overly loose from too much stretching and too few repetitive movements, joints can become destabilized, and in such cases an activity that tightens everything down is one of the best things you can do.
In hatha yoga we usually want to minimize the effects of the myotatic stretch reflex because even moderately dynamic movements will fire the receptors, stimulate the motor neurons, shorten the muscles, and thereby limit stretch. Any dynamic movement in hatha yoga activates the myotatic stretch reflex—bouncy sun salutations, jumping in and out of standing postures, and joints and glands exercises carried off with flair and toss. These are all fine, especially as warm-ups, but if you wish to lengthen muscles and increase flexibility it is better to move into postures slowly.
THE CLASP KNIFE REFLEX
The clasp knife reflex acts like the blade of a pocket knife when it resists closure up to a certain point and then suddenly snaps into its folded position. It is another stretch reflex, but this one causes the targeted muscle to relax rather than contract. The stimulus for the reflex is not dynamic stretch of a muscle spindle, but contractile tension on a sensory receptor in a tendon. This tension reflexly causes the muscle attached to that tendon to relax and the joint to buckle (fig. 1.8).
The sensory receptor for the clasp knife reflex is the Golgi tendon organ. Most of the receptors are actually located near musculotendinous junctions, where they link small slips of connective tissue with their associated muscle fibers. The Golgi tendon organ is therefore activated by the contraction of muscles cells that are in line (in series) with the receptor. Recent studies have clarified that the Golgi tendon organ is relatively insensitive to passive stretch, but that it begins to fire nerve impulses back to the spinal cord as soon as muscle fibers start tugging on it.
And then what happens? This is the main idea: unlike the myotatic stretch reflex, here the incoming sensory axons do not terminate directly on motor neurons (which would increase their activity and stimulate a muscular contraction), but on inhibitory interneurons that diminish the activity of motor neurons and thereby cause the muscle to relax. If you stimulate the receptor, the reflex relaxes the muscle (fig. 1.8). It is a precise feedback loop in which the contraction of muscle fibers shuts down their own activity. This feedback loop works something like a thermostat that shuts off the heat when the temperature rises. Anecdotal reports of superhuman strength in which a parent is able to lift an automobile off her child might be due to a massive central nervous system inhibition of this reflex, like a thermostat that stops working and overheats a house. In ordinary life we see the clasp knife reflex in action, at least in a gross form, when two unequally matched arm wrestlers hold their positions for a few seconds, and suddenly the weaker of the two gives way (fig. 1.8).
Whether intentional or not, we constantly make use of the clasp knife reflex while we are practicing hatha yoga. To see it most effectively and to begin to gain awareness of its utility, measure roughly how far you can come into a forward bend with your knees straight, preferably the first thing in the morning. Then bend the knees enough to flatten the torso against the thighs. Hold that position firmly, keeping the arms tightly wrapped around the thighs to stabilize the back in a comfortable position in relation to the pelvis. Then try to straighten the knees while keeping the chest tightly in place, and hold that position in an intense isometric pull for 30 seconds. This is the hamstrings-quadriceps thigh pull (fig. 1.16), and we’ll examine it in more detail later in this chapter. Release the pose and then check to see how much further you can come into a forward bend with the knees straight. The difference will be a measure of how much the Golgi tendon organs “stimulated” the hamstring muscles to relax by way of the clasp knife reflex.
The Golgi tendon organs are sensitive to manual stimulation as well as to muscular tension. If you manipulate any musculotendinous junction in the body vigorously, its Golgi tendon organs will reflexly cause their associated muscle fibers to relax. This is one of the reasons why deep massage is relaxing. This is also why body therapists wanting to reduce tension in a specific muscle will work directly on its musculotendinous junctions. It’s an old chiropractic trick—manual stimulation stimulates the clasp knife reflex almost as efficiently as contractile tension. Surprisingly, the results last for a day or two, during which time the recipient of the work has a chance to correct the offending musculoskeletal habit that gave rise to the excess tension in the first place.
Although you can test the effects of manual stimulation on tendons anywhere in the body, let’s experiment with the adductor muscles on the inside of the thighs because tight adductors, more than any other muscles, limit your ability to sit straight and comfortably in the classic yoga sitting postures. First test your ability to sit in either the auspicious or accomplished posture (figs. 10.11 and 10.14). Then release the pose and lie with the hips butted up firmly against a wall with the knees extended and the thighs spread out as much as possible for an adductor stretch. With the help of a partner to hold your thighs abducted, try to pull the thighs together isometrically, engaging the adductors as much as possible, and at the same time stimulate the Golgi tendon organs in the adductor muscles with vigorous rubbing. Some of the adductor tendons are the cordlike structures in the inner thighs near the genitals. Others are more flattened and are located further to the rear. All of them take origin from a pair of bones, the inferior pubic rami (fig. 1.12), that together form the rear-facing V which accommodates the genitals.
As you massage the adductors for a minute or so while keeping them under tension, you will feel them gradually release, as evidenced by being able to abduct the thighs more completely. Then sit up and check for improvement in your sitting posture. The combination of massaging the adductor tendons plus making an isometric effort with stretched adductors powerfully inhibits the motor neurons that innervate these muscles, and this allows them to release and permits you to sit straighter and more comfortably.
The hamstrings-quadriceps thigh pull and the adductor massage give us obvious examples of how the clasp knife reflex operates. It is also invoked in a milder form any time you are able to stay comfortable in an active posture for more than 10–15 seconds, which is what we often do in hatha yoga. In this case don’t bounce unless you want to induce the myotatic stretch reflex, and don’t take a posture into the discomfort zone unless you are prepared to trigger flexion reflexes, which we’ll discuss next.
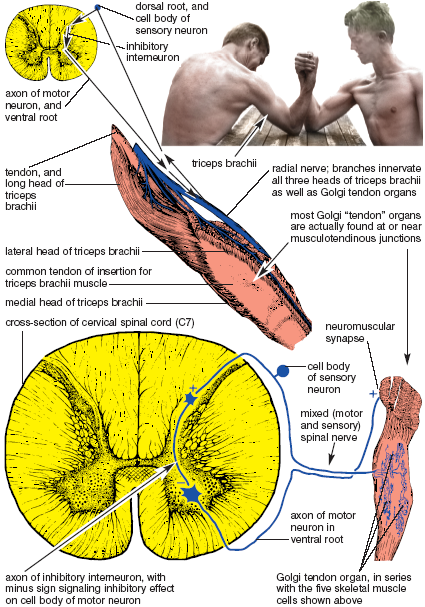
Figure 1.8. The clasp knife reflex. Muscular effort stimulates Golgi tendon organs, whose sensory input to the spinal cord activates inhibitory interneurons (+ in dorsal horn); the inhibitory interneurons then inhibit motor neurons (– in ventral horn), resulting in fewer nerve impulses per second to the skeletal muscle cells (+ effects at the neuromuscular synapse are minimized). Final result is relaxation of the muscle, or in this case, loss of the armwrestling match (Sappey).
FLEXION REFLEXES
The flexion reflexes (fig. 1.9) are pain reflexes. If you inadvertently touch a hot skillet you jerk your hand back reflexly. You don’t have to think about it, it just happens. As with the other reflexes, awareness comes a moment later. Flexion reflexes are more complex than stretch reflexes, but they are easier to comprehend because pain is such an obvious part of everyone’s conscious experience. Even if it is no more than a feeling of stretch that went too far while you were gardening, a pain in the knee or hip that developed after a strenuous hike, or a neck problem you didn’t notice until you started to turn too far in a certain direction, with rare exceptions your automatic response will be flexion. You may be only vaguely aware of the reflex itself, but you will certainly be aware of the fear and tension that accompanies it.
The sensory neurons (including their axons) that carry the modalities of pain and temperature conduct their nerve impulses more slowly than those that activate the myotatic stretch reflex. What is more, flexion reflexes are polysynaptic—that is, they involve one or more interneurons in addition to the sensory and motor neurons—and each synapse in the chain of neurons slows down the speed of the reaction. You can estimate the conduction time for temperature by licking your finger and touching a coffee pot that is hot enough to hurt but not hot enough to cause injury. It will take almost a second for the sensation to reach consciousness from a finger, well over a second from a big toe, and, for the adventuresome, about a tenth of a second from the tip of the nose. Such slow conduction times from the extremities would not serve the myotatic stretch reflex. If, for that reflex, it took a full second for nerve impulses to reach the spinal cord, you would be in serious trouble jumping off a platform onto the floor with bent knees—you would collapse and shatter your kneecaps before the extensor muscles could react enough to support your weight.
Like the two stretch reflexes we have just considered, the motor reflexes for flexion are spinal, not cerebral. So even if the spinal cord were cut off from the brain, the flexion reflex would still withdraw a foot from a toxic stimulus. That’s why neurologists have little reason to be encouraged when the foot of a patient with a spinal cord injury responds to a pinch.
RECIPROCAL INHIBITION
Flexion reflexes not only activate flexor muscles to pull the hand or the foot toward the torso, they also relax the extensors, which then allows flexion to take place freely. This is done through the agency of inhibitory interneurons. While facilitatory interneurons impinge on motor neurons that innervate flexors, thus causing them to contract, inhibitory interneurons impinge on motor neurons that innervate extensors, causing them to relax. The phenomenon is known as reciprocal inhibition, and it is an integral part of the flexion reflex (fig. 1.9).
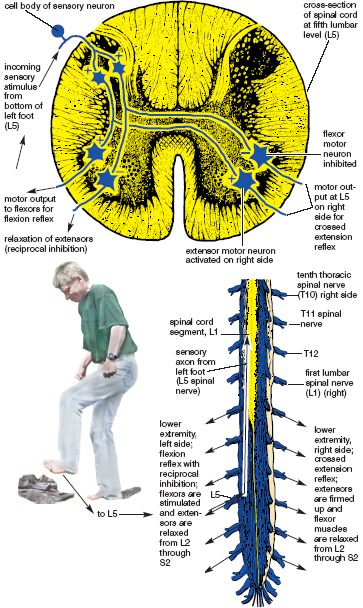
Figure 1.9. Flexion reflex on the left, and crossed extension reflex on the right. Incoming sensory input from bottom of left foot (L5) spreads to lower motor neurons from L2 through S2 for both lower extremities (Sappey).
Unlike stretch reflexes, flexion reflexes create effects well beyond the site of the stimulus. We can see this when a nurse pricks a child’s index finger with a needle to draw blood. The child’s entire upper extremity reacts, not just the flexors in the offended finger. A vehement jerk backward shows that the flexion reflex facilitates flexor motor neurons and inhibits extensor motor neurons for the entire upper extremity.
THE CROSSED-EXTENSION REFLEX
The crossed-extension reflex adds yet another ingredient to flexion reflexes—a supporting role for the opposite side of the body. Through the agency of this reflex, as the extremity on the injured side flexes, the extremity on the other side extends. This would happen if you stepped on a hot coal near a campfire. You don’t have to think about either reflex; you lift your injured foot in a hurry, contracting flexors and relaxing extensors on that side—everything at the same time—toe, ankle, knee, hip, and even the torso. And as the injured foot lifts, the crossed-extension reflex contracts the extensors and relaxes the flexors on the opposite side of the body, strengthening your stance and keeping you from toppling over into the fire (fig. 1.9).
The crossed-extension reflex is accomplished by interneurons whose axons cross to the opposite side of the spinal cord and innervate motor neurons in a reverse pattern from that seen on the side with the injury—motor neurons for extensor muscles are facilitated, and motor neurons for flexor muscles are inhibited.
The flexion reflexes also serve many other protective functions. For example, if you sprain your ankle badly, the pain endings become more sensitive. The next time you start to turn your ankle, the higher centers in the brain associated with volition and consciousness allow the spinal flexion reflexes to act unencumbered and collapse the limb before your weight drops fully on the vulnerable joint. This prevents re-injury. A “trick” knee demonstrates the same mechanisms—an old injury, a sudden physical threat, unresistant higher centers, and unconscious flexion reflexes. Your bad knee buckles quickly, and you are saved from more serious injury.
RECIPROCAL INHIBITION AND A STIFF BACK
Since they restrain movement, flexion reflexes usually influence hatha postures negatively, but there are certain situations in which we can use them to our advantage. If you tend to be stiff and not inclined to forward bending, try this experiment early in the morning. First, for comparison, slowly lower into a standing forward bend with your fingers extended. Notice that you hesitate to come forward smoothly. This may happen even if you are in excellent condition but not warmed up. The deep back muscles are extensors of the back; they lengthen eccentrically as you bend, resisting forward movement and only allowing you come into the posture with creaks and wariness. Come up. Next, holding the elbows partially flexed, flex your fingers tightly, making fists. Holding that gesture, come forward again. You will immediately notice that you do this more easily and smoothly than when your fingers and elbows were extended. Come up, and repeat the exercise to your capacity.
Making your hands into fists reciprocally inhibits the motor neurons that innervate the deep back muscles. If you are in good condition this merely helps you come forward more smoothly and confidently, but if your back is tense from excess muscle tone in the deep back muscles you will be amazed at how much the simple act of flexing your fingers into fists eases you into a relaxed bend.
Why might your back be stiff from excess muscle tone? It is usually because of pain that causes the back muscles to become taut and act as a splint to prevent movement. This is fine for a while as a protective measure, but at some stage it becomes counterproductive and leads to other problems. When stiffness and mild back pain emerge, you need enough muscle tone to prevent re-injury, it is true, but you do not need enough to lock you up for months on end. The reciprocal inhibition caused by making your hands into fists helps relax the extensor muscles in the back and allows you to ease further into a forward bend. If your back is chronically stiff, but not in acute pain, you can benefit by doing this exercise several times a day.
THE VESTIBULAR SYSTEM, SIGHT, AND TOUCH
So far we have seen how motor neurons drive the musculoskeletal system, how association neurons channel our will to the motor neurons, and how sensory input from muscles, tendons, and pain receptors participate with motor neurons in simple reflexes. But that’s only the beginning. Many other sources of sensory input also affect motor function. Some of the most important are the vestibular sense, sight, and touch.
THE VESTIBULAR SENSE
We have little conscious awareness of our vestibular sense even though it is critical for keeping us balanced in the field of gravity. Its receptors lie close to the organ of hearing—the inner ear—in little circular tubes called semicircular canals and in a little reservoir called the utricle, all of which are embedded in the bony region of the skull just underneath the external ear. The semicircular canals and the utricle are all involved with maintaining our equilibrium in space, but within that realm they are sensitive to different stimuli—the semicircular canals to rotary acceleration, and the utricle to linear acceleration and to our orientation in a gravitational field. They also participate in different reflexes: the semicircular canals coordinate eye movements, and the utricles coordinate whole-body postural adjustments.
Except for pilots, dancers, ice skaters, and others who require an acute awareness of equilibrium, most of us take the vestibular system for granted. We don’t notice it because it does almost all of its work reflexly, feeding sensory information into numerous motor circuits that control eye and body movements.
Because the semicircular canals are sensitive to rotary acceleration, they respond when we start or stop any spinning motion of the body. One of their several roles is to help us maintain our equilibrium by coordinating eye movements with movements of the head. You can experience these if you sit cross-legged on a chair or stool that can rotate, tip your head forward about 30°, and have some assistants turn you around and around quickly for 30–40 seconds. Make sure you keep balanced and upright. Don’t lean to the side or you will be pitched off onto the floor. Then have your assistants stop you suddenly. You eyes will exhibit little jerky movements known as nystagmus, and you will probably feel dizzy. Children play with this reflex when they spin themselves until they get dizzy and fall down. The sensation they describe as the world “turning” is due to nystagmus. The perception is disorienting at first but it slows down and stops after a while.
The receptors in the semicircular canals stop sending signals after about 30 seconds of spinning, which is why you have your assistants rotate you for that period of time. It is also why the reaction slows down and stops in 30 seconds after you are abruptly stopped. Third-party observers obviously cannot observe nystagmus during the initial period of acceleration while you are being spun around. To observe these eye movements in a practical setting, we must rely on what we call post-rotatory nystagmus, the eye movements that occur after you have been stopped suddenly.
The neurological circuitry for nystagmus is sensitive to excessive alcohol, and this is why highway patrol officers ask suspected drunks to get out of the car and walk a straight line. If the suspect is suffering from alcohol-induced nystagmus, the ensuing dizziness is likely to make walking straight impossible. Spontaneous (and continuing) forms of nystagmus that are not induced by drugs or alcohol may be symptomatic of neurological problems such as a brain tumor or stroke.
Occasionally students in hatha yoga classes are sensitive to dizziness when they do neck exercises. They may have had such problems from childhood or they may just not be accustomed to the fact that they are stimulating their semicircular canals when they rotate their head. And even otherwise healthy students who are just getting over a fever may be sensitive to dizziness. In any case, anyone who is sensitive should always do neck exercises slowly.
The second component of the vestibular organ, the utricle, detects two modalities: speeding up or slowing down while you are moving in a straight line, and the static orientation of the head in space. The rush of accelerating or decelerating a car is an example of the first case. As with the semicircular canals, stimulation ends after an equilibrium is established, whether sitting still or going 100 miles per hour at a constant rate on a straight road. The utricles also respond to the orientation of the head in the earth’s gravitational field—an upright posture stimulates them the least and the headstand stimulates them the most. The receptors in the utricle adapt to the stimulus of an altered posture after a short time, however, which is why it is so important for pilots of small planes to depend on instruments for keeping properly oriented in the sky when visual feedback is absent or confusing. For example, a friend of mine was piloting a small plane and flew unexpectedly into a thick bank of clouds. Instantly lost and disoriented, and untrained in flying on instruments, he calculated that he would just make a slow l80° turn. Unfortunately, after having made the turn and exiting the clouds, he was shocked to see that he was headed straight toward the ground. Fortunately, he had enough airspace to pull out of the dive.
In ordinary circumstances on the ground, the receptors in the utricle do more than sense the orientation of the head in space: they trigger many whole-body postural reflexes that maintain our balance. This is the source of the impulse to lean into curves while you are running or cycling around a track. We also depend on the utricle for underlying adjustments of hatha yoga postures that we trigger when we tilt the head forward, backward, or to one side. Every shift of the head in space initiates reflexes that aid and abet many of the whole-body postural adjustments in the torso that we take for granted in hatha yoga.
The well-known righting reflexes in cats can give us a hint of how the vestibular system influences posture in humans. If you want to see these reflexes operate, drop an amicable cat, with its legs pointed up, from as little as a few inches above the floor. It will turn with incredible speed and land on all four feet, even if it has been blindfolded. Careful study reveals a definite sequence of events. The utricle first detects being upside down, and then it detects the falling sensation of linear acceleration toward the floor. In response to this the cat automatically rotates its head, which stimulates neck muscles that in turn leads to an agile twisting around of the rest of the body and a nimble landing on all four feet. The cat does all this in a fraction of a second. Comparable reflexes also take place in human beings, although they are not as refined as in cats.
SIGHT
When we are moving we are heavily dependent on vision, as anyone can attest who has stepped off a curb unawares or thought erroneously that one more step remained in a staircase. This is true to a lesser extent when we are standing still. If you stand upright with your feet together and your eyes open, you can remain still and be aware that only minuscule shifts in the muscles of the lower extremities are necessary to maintain your balance. But the moment you close your eyes you will experience more pronounced muscular shifts. For an even more convincing test, come into a posture such as the tree or eagle with your eyes open, establish your balance fully, and then close your eyes. Few people will be able to do this for more than a few seconds before they wobble or fall.
Visual cues are especially important while coming into a hatha yoga posture, but once you are stable you can close your eyes in most poses without losing your balance provided your vestibular system and joint senses are healthy. On the other hand, if you want to study your body’s alignment objectively you can do it only by watching your reflection in a mirror. It is all too easy to deceive yourself if you depend purely on your muscle- and joint-sense to establish right-left balance.
THE SENSE OF TOUCH
The sense of touch brings us awareness of the pleasure and luxury of comfortable stretch, and because of this it is the surest authority we have for telling us how far to go into a hatha yoga posture. The vestibular reflexes and vision help with balance, and pain tells us how far not to go in a stretch. But the sense of touch is a beacon. It both rewards and guides.
The modality of touch includes discriminating touch, deep pressure, and kinesthesis. All three are brought into conscious awareness in the cerebral cortex, and along with stretch reflexes, vision, and the vestibular sense, they make it possible for us to maintain our balance and equilibrium. Discriminating touch is sensed by receptors in the skin, and deep pressure is sensed by receptors in fasciae and internal organs. Kinesthesis, which is the knowledge of where your limbs are located in space, as well as the awareness of whether your joints are folded, straightened, stressed, or comfortable, is sensed mostly by receptors in joints. If you lift up in a posture such as the prone boat and support your weight only on the abdomen, you can feel all three aspects of touch—contact of the skin with the floor, deep pressure in the abdomen, and awareness of extension in the spine and extremities.
Touch receptors adapt even more rapidly than receptors in the vestibular system, which means that they stop sending signals to the central nervous system after a few seconds of stillness. That’s why holding hands with someone gets boring in the absence of occasional squeezing and stroking. Without movement, the awareness of touch disappears. Rapid adaptation to touch is extremely important in hatha yoga postures, relaxation, and meditation. If your posture is stable, the receptors for touch stop sending signals back to the brain and you are able to focus your attention inward, but as soon as you move the signals return and disturb your state of silence.
TOUCH AND THE GATE THEORY OF PAIN
If you bump your shin against something hard, rubbing the injured region alleviates the pain, and if your knee hurts from sitting for a long time in a cross-legged posture, the natural response is to massage the region that is hurting. There is a neurological basis for this—the gate theory of pain, according to which the application of deep touch and pressure closes a “gate” to block the synaptic transmission of pain in the spinal cord. Although it has not been possible to substantiate this theory as it was initially proposed, we all know experientially that somehow it works. So even though the mechanism is still uncertain, the general idea is widely accepted as self-evident—somewhere between the spinal cord and the cerebral cortex, touch and pressure pathways intersect with the ascending pathways for pain and either block or minimize its perception.
We use this principle constantly in hatha yoga. To illustrate, interlock your hands behind your back and press the palms together. Pull them to the rear so they do not come in contact with the back, and come into a forward bend. If you are not warmed up you may notice that you feel mild discomfort from the stretch. Now come up, press the forearms firmly against the back on either side of the spine, and come forward again. The contrast will be startling. The sensation of deep touch and pressure against the back muscles stops the discomfort immediately.
Is this good or bad? That is a vital question, and one of the challenges of hatha yoga is to learn how far this principle can safely be taken. If you underestimate the importance of the signals of pain, and diminish that pain with input from touch and pressure, you may injure joints and tissues. But if you baby yourself, you’ll never progress. The answer, unfortunately, is that you may not know if you have gone too far until the next morning. If you are sore you know you misjudged.
Our bodies are made up of four primary tissues: epithelium, muscle, nervous tissue, and connective tissue. Epithelia form coverings, linings, and most of the internal organs. Muscle is responsible for movement, and nervous tissue is responsible for communication. That leaves connective tissue—the one that binds all the others together. If you were able to remove all the connective tissue from the body, what was left would flatten down on the floor like a hairy, lumpy pancake. You would have no bones, cartilage, joints, fat, or blood, and nothing would be left of your skin except the epidermis, hair, and sweat glands. Muscles and nerves, without connective tissue, would have the consistency of mush. Internal organs would fall apart.
To understand epithelia, muscle, and nervous tissue we have to understand their cells, because it is the cells that are responsible for what the tissue does. Connective tissues are a different matter. With the exception of fat, the one connective tissue that is made up almost entirely of cells, it is the extracellular (outside of cells) substance in each connective tissue that gives it its essential character. The extracellular materials impart hardness to bone, resilience to cartilage, strength to tendons and fasciae, and liquidity to blood. And yet the extracellular components of connective tissues are entirely passive. Trying to relax a ligament or release fasciae with our power of will would be like trying to relax leather.
So are the connective tissues alive? Yes and no. Yes, in that living cells in the various connective tissues manufacture its extracellular components and organize the tissue. Also yes, in that the extracellular space in connective tissue is teeming with electrical activity. But no, in that the extracellular materials are nonliving. And one more no, in that the only way we can access them is through the agency of living cells. Only through neurons and their commands to muscle cells can we release tension in a tendon, execute weight-bearing activities that add bone salts to bone, and stimulate the laying down of additional connective tissue fibers in tendons and fasciae. And only with cells derived from epithelial tissues can we accomplish the absorption, manufacturing, and eliminatory functions that are needed for supporting the tissues of the body in general. In the end, our aim of molding and shaping the extracellular components of our connective tissues can only be accomplished indirectly.
The fact that the various connective tissues are so unlike one another is a reflection of the fact that their extracellular materials are diametrically different. Bone contains bone salts; tendons, ligaments, and fasciae contain dense accumulations of ropy fibers; loose connective tissue contains loose accumulations of the same fibers; elastic connective tissue contains elastic fibers; and blood contains plasma. So we can’t work with connective tissues in general; we have to envision and work with each one individually.
Connective tissues not only give us shape, they also restrain activity. Bone butting against bone brings motion to a dead stop. Cartilage constrains motion, but more softly than bone. Ligaments constrain movements according to their architectural arrangements around joints. Sheets of fasciae, which are essentially layers of connective tissue, enclose and organize muscles and nerves, sometimes more restrictively than we would like. Finally, loose connective tissue helps bond the entire body together, constraining movement between fasciae and skin, adjacent muscle groups, and internal organs.
BONY CONSTRAINTS
Ligaments, muscles, and the joint capsule itself all aid in holding the elbow joint together, but underlying these supports, bony constraints ultimately limit both flexion and extension. Flexion is limited when the head of the radius and the coronoid process of the ulna are stopped in the radial and coronoid fossae in the lower end of the humerus, and extension is stopped when the hooked upper end of the ulna—the olecranon process—comes to a stop in a matching olecranon fossa in the humerus. Even though thin layers of cartilage soften the contact between the radius and ulna in relation to the humerus, the architectural plan limits flexion and extension as certainly as doorstops and provide us with clear examples of bony constraints to movement. It is not something we would want to alter (fig. 1.10).
In the spine we see another example of how one bone butting up against another limits movement. The lumbar spine can extend and flex freely, but matching surfaces of the movable intervertebral joints in this region are oriented vertically in a front-to-back plane that severely limits twisting (fig. 1.11). Because of this, almost all the twisting in a spinal twist takes place in the neck and chest, where the matching surfaces of comparable joints are oriented more propitiously (chapters 4 and 7). As with the elbow, we would not want to alter this design. If the lumbar region, isolated as it is between the pelvis and chest, could twist markedly in addition to bending forward and backward, it would be hopelessly unstable.
CARTILAGINOUS CONSTRAINTS
Cartilage has the consistency of rubber or soft plastic. It gives shape to the nose and external ears, and it forms a cushioning layer at the ends of long bones. Our main concern in this discussion, however, is not with these examples but with the joints called symphyses—the intervertebral disks between adjacent vertebral bodies (figs. 1.11, 4.10b, 4.11, and 4.13b), as well as the pubic symphysis between the two pubic bones (figs. 1.12 and 3.2). At all of these sites symphyses restrict movement, something like soft but thick rubber gaskets glued between blocks of wood that allow a little movement but no slippage. To that end the pubic symphysis is secure enough to bind the two halves of the pelvic bowl together in front and yet permit postural shifts and deviations; intervertebral disks bind adjacent vertebrae together tightly and yet permit the vertebral column as a whole to bend and twist.
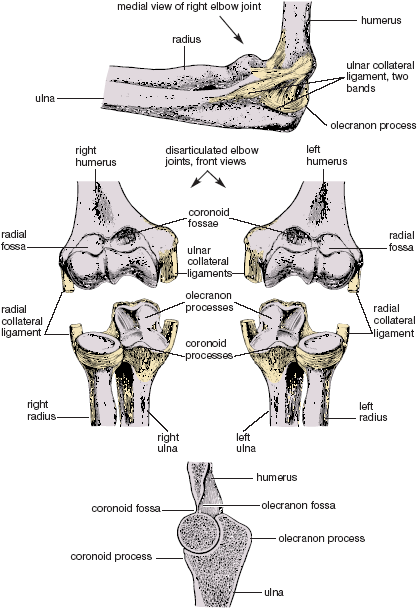
Figure 1.10. Bony stops for elbow flexion and extension, with the joint capsule pictured above, front views of the disarticulated right and left elbow joints shown in the middle, and a longitudinal cut through the joint and two of its three bones shown below. Extension is stopped where the olecranon process butts up against its fossa, and flexion is stopped where the head of the radius and coronoid process butt up against the radial and coronoid fossae (Sappey).
TENDONS AND LIGAMENTS
By definition, tendons connect muscles to bones, and ligaments connect bone to bone. They are both made up of tough, ropy, densely packed, inelastic connective tissue fibers, with only a few cells interspersed between large packets of fibers. Microscopically, tendons and ligaments are nearly identical, although the fibers are not packed as regularly in ligaments as in tendons. In a tendon the fibers extend from the belly of a muscle into the substance of a bone, lending continuity and strength to the whole complex. Ligaments hold adjoining bones together in joints throughout the body, often permitting small gliding motions, and usually becoming taut at the end of a joint’s range of motion.
Ligaments and tendons can accommodate no more than about a 4% increase in length during stretching, after which tearing begins. This can be a serious problem. Because the extracellular connective tissue fibers in tendons and ligaments depend only on a few scattered living cells for repair and replacement, and because the tissue is so poorly supplied with blood vessels, injuries are slow to heal. The most common of these is tendinitis, which is caused by tears in the fibers at the interface between tendon and bone. If someone keeps abusing this interface with repetitive stress, whether typing at a computer keyboard, swinging a tennis racket, or trying compulsively to do a stressful hatha yoga posture, the injury can take a year to heal, or even longer.
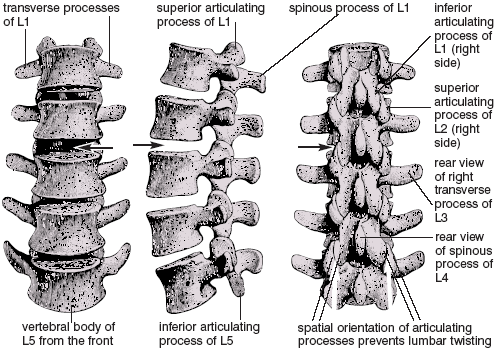
Figure 1.11. Lumbar vertebrae from the front, side, and behind. The vertical, front-to-back orientation of the articulating processes and their joint surfaces provides a bony stop that prevents lumbar twisting. Spaces that represent the location for the intervertebral disk between L2 and L3 are indicated by arrows (Sappey).
The main purpose of ligaments is to restrain movable joints, and this becomes a major concern in hatha yoga when we want to stretch to our maximum. We might at first think of loosening them up and stretching them out so they do not place so many restraints on hatha postures. But ligaments don’t spring back when stretched and lengthened (at least not beyond their 4% maximum), and if we persist in trying to stretch them beyond their limits we often do more harm than good. Once lengthened they become slack, and the joints they protect are prone to dislocation and injury. Ligaments have their purpose; let them be. To improve ranges of motion and flexibility, it is better to concentrate on lengthening muscles.
JOINT CAPSULES
Joint capsules are connective tissue encasements that surround the working surfaces of the class of joints known as synovial joints, including hinge joints, pivot joints, and ball-and-socket joints. Joint capsules for synovial joints have several roles: they provide a container for the slippery synovial fluid that lubricates the mating surfaces of the opposing bones; they house the synovial membrane that secretes the synovial fluid; they provide a tough covering of tissue into which ligaments and tendons can insert; and of special interest to us here, they and their associated ligaments provide about half the total resistance to movement.
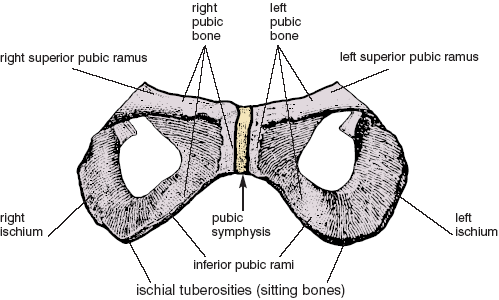
Figure 1.12. Pubic symphysis shown where it joins the two sides of the pelvis. This enlargement of the two pubic bones and ischia (front view) is taken from fig. 3.2, which shows the entire pelvis in perspective (Sappey).
The shoulder joint reveals an excellent example of a joint capsule. Like the hip joint, the shoulder joint is a ball and socket—the ball being the head of the humerus and the socket being the glenoid cavity of the scapula (fig. 1.13). The joint capsule surrounds the entire complex and accommodates tendons that pass through or blend into the joint capsule, as well as ligaments that reinforce it on the outside. To feel how it restricts movement, raise your arm overhead and pull it to the rear as far as possible: within the shoulder you can feel the joint capsule and its ligaments tightening up.
EXTENSILE LIGAMENTS
Extensile ligaments are not really ligaments; they are skeletal muscles held at relatively static lengths by motor neurons firing a continuous train of nerve impulses. They have greater elasticity than connective tissue ligaments because of their muscular nature, but other than that they function to maintain our posture like ordinary ligaments. What they don’t do, by definition, is move joints through their full range of motion, which is what we usually expect from skeletal muscles. According to the conventional definition, extensile ligaments are mostly postural muscles in the torso, but it is arguable that for maintaining a stable meditation posture, every muscle in the body (excepting the muscles of respiration) becomes an extensile ligament.
Unlike connective tissue ligaments, the length of extensile ligaments can be adjusted according to the number of nerve impulses impinging on the muscle. And since every muscle associated with the torso and vertebral column is represented on both sides of the body, the matching muscles in each pair should receive the same number of nerve impulses per second on each side, at least in any static, bilaterally symmetrical posture. If that number is unequal, the paired muscles will develop chronically unequal lengths that result in repercussions throughout the central axis of the body. In hatha yoga, this condition is especially noticeable because it is the primary source of right-left musculoskeletal imbalances.
Axial imbalances can be spotted throughout the torso and vertebral column, but they are especially noticeable in the neck, where the tiny suboccipital muscles function as extensile ligaments to maintain head position (fig. 8.20). If your head is chronically twisted or tipped slightly to one side, it may mean that you have held the matching muscles on the two sides at unequal lengths over a long period of time. Motor neurons have become habituated to long-established firing patterns, the bellies of the muscles themselves have become shorter on one side and longer on the other, and the connective tissue fibers within and surrounding the muscle have adjusted to the unequal lengths of the muscle fibers. Correcting such imbalances requires years of unrelenting effort; neither the bellies of the muscles or their connective tissue fibers can be lengthened or shortened quickly.
Figure 1.13. Disarticulated right shoulder joint as viewed from the front (top image); right shoulder joint with its capsule, also from the front (middle image); and right shoulder joint with its capsule as viewed from behind (bottom image). Envision the chest as being located in front of the scapula and to the observer’s right in the top two images (the surface of the scapula shown here faces the back of the chest). In the bottom image, envision the back of the scapula as being located to the observer’s left; except for being a much deeper and more confined dissection, this view is similar to the one shown in figure 1.1. Asterisks indicate stabilizing ligaments, and arrows indicate rotator cuff tendons (Sappey).
FASCIAE
Fasciae are sheets of connective tissue that give architectural support for tissues and organs throughout the body, holding everything together and providing for a stable infrastructure. They are crudely like leather gloves that form a boundary around your hands. Underneath the skin and subcutaneous connective tissue, fasciae organize and unify groups of muscles, individual muscles, and groups of muscle fibers within each muscle. They form a tough envelope around all the body cavities, and they surround the heart with a heavy connective tissue sack—the fibrous pericardium. We have superficial fascia just underneath the skin, and deep fasciae overlying muscle groups. The singular and plural terms are almost interchangeable—we can say deep fascia of the back, fascia of the body, or fasciae of the body.
Fascia is flexible if we keep moving, stretching, and breathing, but if we allow any part of the body to remain immobile, its fasciae become less flexible and eventually restrict our movements, like gloves that fit so tightly that you can’t bend your fingers.
LOOSE CONNECTIVE TISSUE AND GROUND SUBSTANCE
Loose connective tissue is composed of ground substance, scattered fibers, and cells. It fills in the spaces between the three primary tissues that are mostly cellular—muscle, epithelia, and nervous tissue—and between all the other connective tissues, including bone and cartilage, blood and lymph, tendons and ligaments, joints and joint capsules, fasciae, fat, and lymphatic tissue. But loose connective tissue is more than a filler. Its ground substance is crudely comparable to glycerine—it lubricates and smoothes movement throughout the body. The ground substance permits slippage of adjacent structures as well as slippage of individual connective tissue fibers relative to one another in tendons and ligaments. Were it not for the connective tissue fibers and their submicroscopic attachments to muscle fibers, nerve fibers, and epithelia, ground substance would let everything slip and slide apart. This would be as unwelcome—by itself—as an oil spill on an icy road.
Ground substance is normally fluid, but it congeals and loses moisture if the surrounding tissues are inactive. And as it loses moisture it loses its lubricating properties. The entire body tightens down. Tendons, ligaments, and joint capsules become brittle, muscles lose much of their elasticity and ability to function smoothly, and the tissues become susceptible to injury. These reversals are the main reasons for morning stiffness, and they are a compelling argument for beginning every day with a session of hatha yoga. To rehydrate the ground substance a short, lazy practice is not as effective as a long and vigorous one, and you get what you pay for. The benefit is well-being; the payment is work and stretch.
If you ask most people what it takes to lift a barbell overhead they will say muscles, bones, and joints. If you ask them what is needed for running a marathon they will say heart, lungs, and legs. And if you ask them what is required for dance or gymnastic performance they will say strength, grace, and agility. But if you ask them what is most important for increasing flexibility they will probably just give you a blank look. And yet those of us who practice hatha yoga know that improving flexibility is one of our greatest challenges. Even the simplest postures are difficult when one is stiff, and that is why instructors are always encouraging us to stretch. But what exactly do they mean?
Given what we have discussed so far, we know that we should be wary of increasing flexibility by trying to free up bony stops or loosening up cartilaginous restraints, joint capsules, tendons, and ligaments. What we can do is lengthen nerves and the bellies of muscles, the two kinds of extendable anatomical structures that run lengthwise through limbs and across joints.
THE BELLIES OF MUSCLES
Muscles have to be lengthened only a little to permit a respectable improvement in a joint’s range of motion. But when we are stretching them and looking for long-term results, are we dealing with their individual muscle fibers or with their associated connective tissue fibers? The answer is both. The individual muscle fibers within a muscle can grow in length by the addition of little contractile units called sarcomeres. We know this from studies of muscles that have been held in casts in stretched positions. And by the same token, if a muscle is held by a cast in a foreshortened state, sarcomeres are lost and the muscle fibers become shorter.
It is not enough to increase the length of muscle fibers alone. A matching expansion of the connective tissue within and around the muscle is also needed, including the overlying fascia, the connective tissue that surrounds packets of muscle fibers, and the wrappings of individual fibers. And this is what happens during programs of prolonged stretching. The connective tissue gradually follows the lead of the muscle fibers, the muscle as a whole gets longer, and flexibility is improved. Hatha yoga stretches are a safe and effective way to bring this about. And in the occasional cases when we want to tighten everything down, all we have to do is stop stretching so much and concentrate on repetitive movements and short ranges of motion. The muscle fibers will quickly get shorter, and the connective tissues will soon follow suit.
NERVES
The issue of peripheral nerves is another matter. Nerves are sensitive to stretch but are not robust enough to limit it; they can accommodate to stretch only because they don’t take a straight course through the tissues that surround them and because their individual nerve fibers meander back and forth within the connective tissue ensheathments of the nerve itself. During the course of stretching a limb, the gross path of a nerve through the surrounding tissues is first straightened, and as the stretch continues, the serpentine course of the individual fibers within the nerve is also straightened. And even after that, the enveloping connective tissue has enough elasticity to accommodate about 10–15% additional stretch without damaging the nerve fibers.
Without their connective tissue ensheathments nerves would be hopelessly vulnerable, not only to stretch but also to trauma and compression among tense muscles, bones, and ligaments. The protection is not fail-safe, however, because in extreme cases these ensheathments can accommodate to even more stretch than the 10–15% that is safe for their enclosed nerve fibers. The early warning signs are numbness, sensitivity, and tingling, and if these are ignored, sensory and motor deficits may develop. Your best protection is awareness and patience—awareness of why nerve stretch is a potential problem and the patience to work slowly when and if mild symptoms emerge. If nerve pain keeps turning up chronically, seek professional help.
IN THE LAST ANALYSIS
Research has shown beyond question that the length of muscle fibers can be increased as a result of prolonged stretching, or decreased as a result of chronic foreshortening. It is also clear that the connective tissue ensheathments of either muscles or nerves can be stretched too much. But there is another factor in the equation: the nervous system plays a pivotal role in causing muscles to either relax or tighten up, and this either permits stretch or limits it. So is it the active role of the nervous system or the passive role of the connective tissues that ultimately restrains movement? Since nerve impulses keep stimulating muscle cells during ordinary activities, there is only one way to find out for certain: to check someone’s range of motion when they are under deep anesthesia, when the nervous system is not stimulating any skeletal muscle cells except those needed for respiration.
This has been done, inadvertently but many times. Any operating room attendant can tell you that when patients are anesthetized, their muscles become so loose that care has to be taken not to dislocate the joints, and this will happen even if the patient is extremely stiff in waking life. So why can’t therapists take advantage of anesthesia-induced flexibility to increase the range of motion around joints? The answer is that without the protection of the nervous system the tissues tear—muscle fibers, connective tissue fibers, and nerves. And this proves that even though connective tissues provide the outermost limits to stretch, it is the nervous system that provides the practical limits in day-to-day life. When we have reached those limits the nervous system warns us through pain, trembling, or simply weakness that we are going too far, and most important, it warns us before the tissues are torn.
Three hatha yoga postures illustrate the principles of movement we have been discussing. They are all simple to analyze and study because they exhibit bilateral symmetry, in which the two sides of the body are identical in structure and perform identical movements. Each one presents different challenges. We’ll begin with the corpse posture.
THE CORPSE POSTURE
The corpse posture reveals several common problems that arise when people try to relax. Lie supine on a padded surface with the knees straight, the feet apart, the hands out from the thighs, and the palms up. Relax completely, allowing your body to rest on the floor under the influence of gravity (fig. 1.14). When you first lie down most of the motor neurons that innervate the skeletal muscles are still firing nerve impulses, but your breathing gradually becomes even and regular, and the number of nerve impulses per second to your muscles starts to drop. If you are an expert in relaxation, within a minute or two the number of nerve impulses to the muscles of your hands and toes goes to zero. Then, within five minutes the motor neuronal input to the muscles of your forearms, arms, legs, and thighs diminishes and also approaches zero. The rhythmical movement of the respiratory diaphragm lulls you into even deeper relaxation, finally minimizing the nerve impulses to the deep postural muscles of the torso. The connective tissues are not restraining you. Pain is not registered from any part of the body—the posture is entirely comfortable. This is an ideal relaxation.
In the early stages of practice any number of problems can interfere with the ideal. First, let’s say you hurt your right shoulder playing basketball earlier in the day. Tension in that region is still high and stands out painfully in contrast to the relaxation in the rest of the limb and on the other side. In addition, you have an old back injury and the muscles around the vertebral column are holding it in a state of tension. You would like to lift your knees to relieve the stress, but you do not wish to seem unsporting. So you override the impulses of the flexion reflexes and continue to suffer with your knees straight.
This is absurd. All problems in the body tattle on themselves in one way or another, and you cannot relax your body because it is rebelling. You would not be in so much pain if you were walking around the block because the movement would keep you from noticing it, but when you try to relax you are aware of nothing else. The posture becomes increasingly irritating, and your mind, far from being still, is oscillating between awareness of the discomfort and longing for escape. If your instructor holds you in this pose for more than a minute or two you are in the wrong class. You are not yet ready for this work. You need to heal, move, and stretch—not lie still.
Those who are uncomfortable can sometimes improve the situation by simply moving into partially flexed positions—bending the knees, placing the hands on the chest, and supporting the head with a thick pillow. For restful sleep, it is not surprising that most people lie on their sides and curl up in an attitude of flexion.
THE PRONE BOAT
The prone boat posture demonstrates the simplest kind of movement against gravity. To experience this lie face down on the floor. Stretch your arms toward the feet, straight out to the sides, or overhead, as you prefer. Raise the arms, thighs, and head away from the floor all at once, keeping the knees and elbows extended (fig. 1.15). You are lifting into the posture with the muscles on the posterior (back) side of the body. The neck, back, hamstring, and calf muscles are all shortening concentrically and drawing the body up in an arc.

Figure 1.14. The corpse posture, for whole-body relaxation.
Although by most standards the prone boat is an easy posture, especially with the hands alongside the thighs, it can be challenging if you are in poor physical condition. A set of muscles is being used which is rarely exercised as a group in daily life, and if you keep your elbows and knees extended you may not be able to lift your hands and feet more than an inch or so off the floor. The combination of inflexibility and unfamiliarity keeps the antagonist muscles on the anterior (front) side of the body active, and this in turn restrains the lift. Whole-body extension is the essence of the prone boat, but the pull of gravity, lack of strength posteriorly, muscular resistance anteriorly, an abundance of flexion reflexes, and various connective tissue restrictions in the spine may all limit you. For beginners the activity of the nervous system is the main impediment to the posture.
Fascia is the main obstacle for intermediate students. The nervous system is commanding the posterior muscles to contract strongly and the anterior muscles to relax, but connective tissues and the design of the joints prevent marked extension. With time and practice the anterior muscles will relax and permit a full stretch. Finally, advanced students confidently lift to their maximum and play with the edges of neuronal control, tugging on their connective tissues with an educated awareness while at the same time keeping the breath even and regular without straining or faltering.
THE HAMSTRINGS-QUADRICEPS THIGH PULL
This standing forward bend demonstrates the interactions among agonist muscles, their antagonists, gravity, and the clasp knife reflex. Stand with the feet about 12 inches apart. Flexing the knees as necessary, bend forward and press the torso tightly against the thighs, which keeps the back relatively straight and prevents strain. Now, holding the chest and abdomen firmly in place, try to straighten the knee joint. The quadriceps femoris muscles on the front of the thighs try to accomplish this, and the hamstring muscles on the back side of the thighs resist, but you can undercut the hamstring resistance by activating their clasp knife reflexes. Just massage the musculotendinous junctions of the hamstring muscles behind the knee joints while you are trying to press your hips up (fig. 1.16).

Figure 1.15. The prone boat. As you lift up into the posture, muscles on the back side of the body shorten concentrically; as you slowly lower yourself down, they lengthen eccentrically. Tension in muscles and connective tissues on the front side of the body increases as you lift up and decreases as you come down.
At this point the quadriceps femoris muscles are shortening concentrically, straightening the knees, and raising the body up against the force of gravity. At the same time the hamstring muscles, which are antagonists to the quadriceps, are actively even though unconsciously resisting. If you are in good condition your nervous system allows you to press upward to your personal maximum, but if you have recently hurt your knee or sprained your ankle, flexion reflexes responding to pain will limit you. As you press up you are making an isotonic movement. If you go to your maximum but then keep pressing, you are exercising isometrically.
If you were to bounce, which we do not want here, you would stretch the muscle spindles dynamically and stimulate the myotatic stretch reflexes. If you move slowly, you will be stimulating the Golgi tendon organs and eliciting the clasp knife reflex in both the quadriceps femoris and the hamstring muscles. Although this will tend to relax both sets of muscles, the focus of your will is to straighten the knee joint, with the result that the higher centers of the brain override the reflex in the quadriceps femori and allow it fuller rein in the hamstrings. Neurological circuits for reciprocal innervation also probably inhibit the motor neurons whose axons innervate the hamstring muscles.
The main resistance to lifting up comes from the hamstrings. If you are an advanced student and not feeling any trace of joint pain, you can try to relax the hamstrings and extend the knee joints more completely, contracting the quadriceps as much as your strength and health permit. This posture is different from the prone boat, in that your attention is more restricted. In the prone boat you are trying to relax the entire front side of the body; here you are trying to relax only the hamstrings.

Figure 1.16. Standing hamstrings-quadriceps thigh pull. The first priority is placing the torso solidly against the thighs in order to protect the lower back. Under those circumstances, trying to lift the hips forceably in combination with massaging the Golgi tendon organs in the hamstring tendons encourages deep relaxation and eventual lengthening of the hamstring muscles (simulation).
If you are relatively healthy, as you reach the limits of nervous system control, the fasciae begin to play an important role in limiting your efforts to straighten the knees and raise up. You reach a point at which the connective tissue fibers within and surrounding the hamstring muscles will not allow any more lifting. They are now like wires pulled taut, stretched to their limit. The only way to get more length in the system is to patiently lengthen the muscles and nerves with a long-term program of prolonged stretches.
We have covered a great deal of territory in this chapter, but in so doing we have laid the foundation for everything that is to come. To sum it up: sensory input to the brain and the power of will both ultimately influence the motor neurons, which in turn preside over the actions of the musculoskeletal system. The reflexes are in the background and out of our immediate awareness, but without them we would be in dire straits. Without the stretch reflexes our movements would be jerky and uncertain, like film portrayals of Frankenstein’s monster. And without pain receptors and flexion reflexes we would soon be a battleground of burns and injuries. Without the reflexes from our vestibular system we would teeter about, uncertain of our balance and orientation. Without sensation from touch and pressure pathways we would lose most of the sensory input that gives us pleasure—and along with its loss, its guidance. In the end, the nervous system drives the musculoskeletal system, and these two in combination maintain and sculpt connective tissues, which in turn passively restrict movement and posture. All of this takes place within the field of gravity under the auspices of will and creates the practice of hatha yoga.
“Fortunately, science is self-correcting. Whatever the herd is doing, truth eventually becomes known and stops the stampede.”
— Gershon, in The Second Brain, p.35.