TREES: A BRIEFING
Some people like a briefing before they plunge into new territory, others want to learn as they go. Readers can bone up here on tree basics, before exploring specific trees in our nontechnical profiles; or they can dive into the guide at any point, coming up for orientation as needed.
What follows is a crash course on tree life and terminology. In the tree profiles themselves, you’ll find the exceptions to the rules, details, and additional subjects that are better illustrated by example. For instance, the particular ways of pine trees are treated in the “Austrian and Scots Pine” profile. The Index locates these subjects. Our Glossary helps define specialized terms, including those in italics here.
WHAT IS A TREE?
We usually know a tree when we see one, but what about palms and certain shrubs and vines? Are they trees? It depends on whose definition you use, and there are several around, none fail-safe. They go something like this: a tree is a woody plant, a perennial that does not die back after its reproduction cycle; it grows to at least 20 feet tall (some say 13, some 15 or 17), with an erect main stem (trunk) at least three inches in diameter (at “breast height,” or 4½ feet). It bears leaves on a crown or mass of branches and tends toward a characteristic habit or shape.

These definitions, however, rule out such undersized, spindly, multistemmed plants as crapemyrtle, a beloved blossoming tree in the South. Several types of urban trees are squat or multitrunked or both; they might be a shrubby or downsized form of an ordinarily big tree. We count them as trees here, finding no lack of bona fide tree features in their parts.
A crown of branches and layer of bark usually indicate a tree, but the tree-sized species of palm have neither. Their crowns consist of leaves growing out of one large bud atop the stem. Only a rind covers the fibrous stem, and not the corky, regenerating material we call bark. Still, the bigger palms look like trees, serve as trees, and can be studied as a special family of trees, with woodlike stems (see “Palm” profile).
TREE WOOD
The very body of trees, true wood consists mainly of cellulose-walled cells, with a substance called lignin that encrusts those walls. Pectin (as in your favorite jelly) binds them together. Along with fibers made of interlocking cells, lignin gives wood its characteristic hardness and strength.
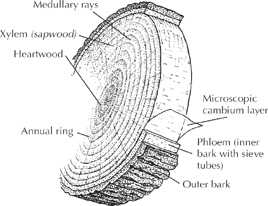
Wood is produced by an uncanny membrane called the cambium layer, a microscopically thin film of cells underlying the entire bark surface of the tree, including the roots. The soul of woody plants, if anything might be, this protoplasm-rich cambium would look like nothing more than a slimy transparent film if one were to surgically uncover it. But for at least part of the year, it is a heavy-duty factory churning out two critical products: wood cells (xylem), released toward the interior of the tree, and “sieve-tube” cells (phloem), released toward the outside.
The xylem cells build up into sapwood, the material that helps support the tree’s structure but also conducts water and nutrients upward. Its pipelines run from roots to leaves. As new sapwood is formed and the tree gets thicker, the old sapwood gums up and becomes inactive tissue called heartwood, accumulating into a dense, usually darker core surrounded by the sapwood. Eventually, the heartwood dominates an aging tree’s interior.
Each of the rings seen on a cross section of trunk or limb reflects a year’s production of sapwood cells. Cells produced in spring tend to be bigger than summer cells and appear lighter-colored en masse. The contrast between the denser summer cells and the lighter cells of the next spring creates the borders of the rings. Not only can the tree’s approximate age (or age of a branch) be read from these rings, but also whether a year was lean (narrow ring) or fertile, or marked by such traumas as fire or infestations. Thin conductive channels called medullary rays cut across these rings like spokes, from the core to the exterior layers.
On its outward surface, the cambium forms the phloem layers, a spongy network of tubes that transport and store the sugary food manufactured by the leaves. The phloem, sometimes called the inner bark, distributes this sappy food downward to the roots and other growing parts, while the interior xylem transports watery nutrients upward. (To remember which is which, think, “A river phloes downstream.”)
New phloem cells do not accumulate indefinitely like wood cells; old ones deteriorate under pressure and are replaced. But a cork cambium at the outer edge of the phloem manufactures oily-walled, waterproof cells that protect the tree and, when dead, form the outer bark. These bark layers do build up somewhat, even as they stretch, split, flake, and slough off.
Wood quality and character vary tremendously, depending on species. Some of the more extraordinary woods are encountered in our profiles.
TREE TYPES
Any tree watcher can tell that some trees keep their foliage all year, while others go bare-branched at least part of winter. The trees that drop their leaves are deciduous, those that keep them (or, actually, replace them before they all drop) are called evergreen. Within the evergreen group are most of the conifers, those trees with woody cones and needle-shaped leaves like pine and spruce, or leaves made up of tightly overlapping scales, like arborvitae.
You may recall from school biology that conifers belong to the plant group known as gymnosperms, a term referring to the “naked seed,” a seed not born of a flower’s ovary. Peer into a cone whose scales are just yawning open, and you’ll find such seeds. They may have a wing, but no covering formed of a matured ovary and other flower parts. In contrast, notice how the seeds of an apple are enveloped by the core, which develops from the ovary of the apple blossom. The more evolved “covered seed” trees, such as our familiar fruit, nut, and shade trees, are called angiosperms. Most of the nonneedle trees (broadleafs) belong to this group.

Pine cone
The conifers are by no means the only evergreens. Many broadleaf trees that grow in warm climates—such as live oak, magnolia, and eucalyptus—maintain their foliage year-round.
Sometimes, the deciduous trees are called “hardwoods” and the conifers “softwoods,” but this is a loose division more meaningful to the lumber industry than to the urban tree lover.
To bring precision to the study of tree types, botanists, after countless thousands of close observations, have organized trees according to certain shared features, just as other plant and animal types have been organized. The organization scheme starts with families (e.g., Willow) of generally related trees, subdivided into genera (e.g., the genus poplar) of closely related trees, within which are grouped the unique tree types or species (e.g., trembling aspen). Beyond species are the variations, if any, on a unique type. These are called varieties if they are found in nature, and cultivated varieties or named varieties if cultivated by horticulturists (e.g., the weeping ‘Pendula’ variety of American aspen).
Species commonly crossbreed with others in their genus when they are similar enough in makeup. The offspring are called hybrids. Often a hybrid shows increased growth and survival energy, a phenomenon called “hybrid vigor.” Both nature and human cultivators take advantage of hybridization in creating the fittest or the most pleasing trees. The London planetree, a cross between two sycamore species, is one of the world’s most successful street trees.
Plants do not always slip easily into a category, and botanists frequently disagree on their place; but the scheme basically works. An old mnemonic, “King Philip Came Over For Gene’s Special Variety,” helps nonscientists recite the scientific hierarchy of all life forms: kingdom, phylum, class, order, family, genus, species, variety.* Only the last four concern us in this guide, not for recitation but for seeing an urban tree’s place in the world of trees. We can also pick up on family traits and eccentricities—we know, for example, that Mr. Cottonwood of the thirsty Willows cannot be trusted around the sewer pipes.
TREE NAMES
In cities we have our stinktree, stinking maple, ghost tree, honeyshucks, pussy willow, Mormon tree, and cigar tree, to mention a few by popular or “common” name. For all their personality, such names create identification problems. They vary by region and nation; they overlap, they change. Some trees have more than 30 common names within one language.
These common names usually evolve from how or where the trees were used by people over time, who first studied them, or what struck observers as distinctive. Native Americans supplied the name “catalpa” from words probably describing the big flowers of this tree. But the northern catalpa is also known as the “cigar tree,” for its long seed pods, and the southern catalpa as the “fish-bait tree,” after the caterpillar larvae that cover the tree in deep-South fishing country.
Again for greater precision, scientists have adopted a naming system based largely on Latin and designed to fix every species with a standard, unique name, to be used and understood throughout the world.
This binomial or two-part naming system uses the genus name combined with a descriptive name or “epithet”—something like a surname and given name. Thus, the species that some people call trembling aspen, or quaking aspen, or Canadian aspen, or quiver-leaf, is nailed down scientifically as Populus tremuloides. (Populus is the ancient Latin word for poplars, and tremuloides here means “like that which trembles.”) The same in every nation, the two-part scientific name distinguishes this Populus from every other, such as Populus alba or Populus grandidentata.
The name of a variety takes this form: Malus baccata var. himalaica (Himalayan crabapple, variety of Siberian crabapple). A hybrid (crossbreed) gets an “x” in its name, as in Platanus x acerifolia, the London planetree.
A few descriptive terms such as “contorta” or “pubescens” may charm those who can guess their meanings, but many of these scientific names can be offputting to the layperson. Anyone who can pronounce “Malus sieversii ‘Niedzwetzkyana’” deserves a garland of this crabapple’s magenta blossoms. However, because the names are essential for indicating an exact tree species or looking it up in other sources, we provide them in each tree profile along with one or more common names.
TREE PARTS
Like all complex organisms, a tree is an engineering marvel, a system that sustains the individual and produces offspring under environmental shifts and strains. Woodiness aside, a tree is comparable to most other plants in the functions of its parts and specialized cells. But in scale and relation to our own lives, tree parts seem in a class by themselves.
Roots
Roots work several jobs simultaneously in their underground sweatshop. They hold the tree in position; gather all the water needed by the leaves for food making; absorb key minerals for the tree’s life processes; and filter out unwanted substances.
The anchoring job usually starts with a taproot that drives straight down before branching out. Some trees, like certain pines and oaks, eventually grow lateral roots but keep the taproot going as a support or water probe; other trees quickly shift to a wide-branching network of surface roots, within or close to the nutrient-rich topsoil.

The shape of a root mass is as variable as the crown shape. Sometimes, it mirrors the crown, stretching to the drip-line or margin of outermost leaves where the water drips off. Or the roots might form a tight clump where water collects under a narrow tree. Whatever their shape, roots must keep up with the water-greedy food factories within the leaves. They do so not simply by their mass, but with an advancing army of tiny, helmeted “root tips”—often millions of them—equipped with thousands of “root hairs” that absorb water and grip soil grains as they loot their minerals.
Roots absorb and elongate only at the probing root tips, which deftly twist their protective, lubricated caps through paths of least resistance. The rest of the root thickens with wood, presses against the soil in its anchoring role, and conducts water and nutrients from the root tips upward.
Tree roots often enter into mutually beneficial relations with the roots of neighboring trees or with fungi and other organisms, usually swapping sugary food for nitrogen and hard-to-break-down minerals. The acid-alkaline ratio of the soil (expressed as a pH value) may affect these friendships, as it does the chemical functioning of the host roots. Neutral soil rates a pH of 7; higher is alkaline, lower is acidic. Each species has its general pH preference, with some species more flexible than others.
A few trees, such as the city-loving ailanthus, are reputedly unsociable at the root level. They put out toxic chemicals to discourage competitors.
Trunk, Bark, and Branches
The trunk or stem of a tree is both support tower and pipeline. It elevates the foliage to the light source, sometimes raising it more than 200 feet in competition for a place in the sun. Most trunks take one of two basic patterns: the undivided, tapering pole of the typical conifer, with many horizontal branches in an overall pyramid shape; and the maple-or elm-type trunk that divides partway up to form subdividing limbs and branches.
In the crowded forest, the branchless, leafless part of the trunk (sometimes called the bole) may rise for most of the tree’s height. Low foliage would die in the shadows. Trees in open spaces, including urban parks and wide streets, tend to have crowns that start lower and spread wider.
Within these general patterns, trees exhibit hundreds of characteristic shapes according to species and a million individual styles. Live oaks spread wide enough to cross boulevards. Western redcedar is buttressed at its base like a cathedral wall. A willow may not only weep, but prostrate itself along a riverbank. Lombardy poplar, narrow as a smokestack, is one of many “columnar” trees. As it ages, Austrian pine transforms itself from robust, orderly pyramid to eccentric, flat-topped scarecrow.
A tree’s diameter increases from one source only: the output of the cambium. All other aboveground growth takes place at the “leader” and terminal buds, those vigorous tips at the extreme ends of the main trunk (or trunks) and branch twigs. Each growing season, the tree packs these buds with an embryonic shoot and sometimes side shoots. The next growing season, these shoots will develop into extensions of twig, branchlet, or trunk. Bud scars usually mark the points at which the last few extensions began.
Over the years, as the cambium adds wood and bark to these extensions, twigs become branchlets, branchlets become branches, and along the trunk, branches thicken into heavy limbs. The leader bud extends only the top of the trunk; initials carved in the bole 30 years ago remain at their original height.

Branchlet
Yearly growth rate can range from millimeters, as with the bristlecone pines, the world’s oldest living trees, to the 20-foot leaps of some eucalypts. Frequently, the slower-growing trees are stronger and more enduring. Cool climates often slow growth.
The xylem, or sapwood, as described above, conducts mineral-rich water through the trunk to the food-making chambers of the leaves, where evaporation through tiny pores results in tremendous water loss. But evaporation also contributes to a water “deficit,” a sort of vacuum or tension. This deficit pulls at tree-length, cohesive columns of water in the xylem tubes, and the columns move up to keep water in the leaf. Incompletely understood, the cohesion is one of the forces that, along with evaporation and root pressure, can move tons of water from the soil to the top of a 300-foot redwood.

Some bark types
The xylem brings water not just to leaves, but to all other parts of the tree, keeping the billions of cells fully charged with water.
Meanwhile, a sap of starches and sugars produced by the leaves travels down the phloem (inner bark) of twig, branch, and trunk. Cells convert it to fats, oils, and proteins as needed by the tree parts. The cambium layer and roots depend on a steady flow of this fuel. When a tree’s phloem is cut off, as by a tight girdle of wire around the trunk, the parts above the girdle live for a while; but soon the roots stop growing, and finally the tree dies of malnutrition. Bark stripping, too, is a form of tree murder or dendrocide, taking with it the phloem and cambium.
Bark may host thousands of living organisms, but essentially it is dead tissue from the cork cambium. The way it stretches, splits, or peels as the tree widens creates patterns that excite landscapers and announce the tree’s identity. Our profiles describe the “camouflage” pattern of sycamores, the torn-paper texture of river birch, the thick multicolored slabs of ponderosa bark, and much more. Often, bark is ornamented by raised, horizontal rows of breathing pores called lenticels. Some cherry trees wear their lenticels like designer stripes.
Waterproof and often rich in chemicals, bark has served humankind since prehistory, as has fibrous inner bark. The multitude of applications includes canoes from birch bark, aspirin from willow bark, and cork, the harvested bark of cork oak.
Leaves
Leaves begin life in a specialized bud. There, enfolded by leaflike bud scales, they form over a summer into embryonic versions of their mature selves. They remain in the bud until the following spring (in zones with varying seasons), when rising sap energizes the bud, the bud swells, the scales open, and out comes an intricately folded leaf with adult features in miniature. It grows with astounding speed to its full size, the better to build the tree’s food supply rather than drain it.
The food-making process takes place in the blade of the leaf or leafblade, the main part, usually held out and positioned for favorable light by a leafstalk or petiole. In broadleaf trees the blade is flat, though sometimes very narrow. Conifers have needlelike blades, often stiff, or blades of small, tightly overlapping scales. But virtually all leafblades have the factory parts for the process of photosynthesis, the light-driven conversion of carbon dioxide and water into simple sugars and oxygen.
Humans and other animals might be considered satellite factories, sucking up the freed oxygen for their own life processes and gladly supplying exhaled carbon dioxide in return. But the central factory is the leafblade, whose parts include:
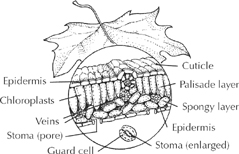
Leaf, cross-section
 Xylem and phloem pipelines (some visible as “veins”) to carry water in and sugar out, respectively.
Xylem and phloem pipelines (some visible as “veins”) to carry water in and sugar out, respectively.- A waxy skin (“cuticle”) that admits light and limits water loss.
- Tens of thousands of air intakes called stomata (“little mouths” in Greek) on the blade’s lower surface. Lined with sausage-shaped valves, they open in the light to admit air and its carbon dioxide, and close to conserve resources. On some conifer needles, masses of stomata can be seen as straight white stripes.
- Layers with two types of cells containing microscopic, chlorophyll-loaded chloroplasts. The chloroplasts interact with light, water, and carbon dioxide to produce the sugars—and also give green color to the leaf.
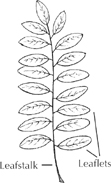
Compound pinnate leaf (alternate)
Minuscule hairs frequently cover the lower surface of leafblades to protect the stomata from water loss and infection.
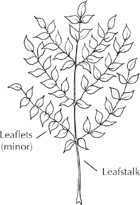
What botanists call a leaf begins not at the blade, but at the point where leafstalk attaches to wood. In numerous species, this leafstalk subdivides into several more stalks, each with its own blade. The whole structure is called a compound leaf, meaning made up of three or more leaflets rather than one simple leaf. Each leaflet looks and functions like a simple leaf, but its stalk lacks the base that snaps off cleanly from a twig. The stalk of a leaflet can only be torn away from where it joins the main leaf. When a main leaf is snapped off the twig, it reveals a leafscar on the twig surface. Within this scar is a pattern of vascular bundle scars representing the pipelines that had connected tree and leaf. Winter observers often use these patterns for tree identificaton.
A familiar compound leaf is that of the ash tree, with leaflets in opposite pairs along a main stalk. This featherlike arrangement is called pinnate. Sometimes the stalk subdivides twice, in a bipinnate pattern, such as that seen on some honeylocust leaves.
When leaflets extend from one point like fingers from a palm, the leaf is called palmate. The horse chestnut leaf is both compound and palmate, with its five or more long leaflets extending from the end of the main leafstalk.
Compound bipinnate leaf
Leaves arrange themselves in certain patterns on their branchlets. Opposite leaves, like those of ashes and maples, grow on each side of the twig in exactly opposite pairs. Most broadleafs are alternate, with the leafstalks at staggered intervals along the two sides, not matching up. And many trees, including conifers, arrange their leaves in whorled patterns, the leafstalks circling the branchlet or spiraling around it. Along with other features, these patterns can help identify a tree; but they can be subtle and tricky. Our profiles point them out when of special interest.
The shapes and textures of leafblades themselves are usually an observer’s best friend, not only for tree identification, but also for the aesthetic pleasure they give in their endless variations on a theme. From the coin-shaped aspen to the star-shaped sweetgum, the silky Russian olive to the waxy magnolia, you’ll discover a universe of leaf design among our urban trees.
Up to a point, the technical terms for leaf parts aid the casual observer. The pointed or rounded ears of a leaf are called lobes; there is no better word for them. Instead of “the dips or valleys or coves or spaces between the lobes,” one can say sinuses. The continuation of the leafstalk into the middle of the blade is the midrib. The “edges” of a leaf could mean just its sides, but margin clearly refers to the whole perimeter, including the “base” around the leafstalk and the apex or tip at the opposite end.
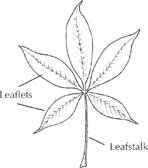
Compound palmate leaf
Among tree professionals, other terms save time and avoid confusion but can overwhelm the amateur. Oblanceolate is one of more than a dozen terms describing leaf shapes, while other technical glossaries describe the leaf margins, bases, tips, vein patterns, and surface textures. In the profiles, we use ordinary language such as “heart-shaped” or more ambitious metaphors when they do the job. Our accompanying drawings are often the most telling descriptions, showing just what is meant, for example, by the “double teeth” of the elm leaf.
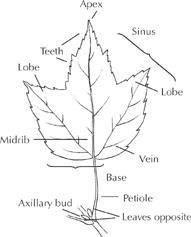
Leaf (simple)
Leaves of deciduous trees turn color when their green chlorophyll pigments break down, the work of photosynthesis completed for the season. Other lurking but less abundant pigments such as red and yellow now show their stuff. Even as they do so, a fragile layer of cells forms between the base of the leafstalk and the twig (except in oaks and beeches) and severs the leaf from the tree. It is “fall.”
Flowers
Tree observers welcome flowers as identification clues—not to mention the thrill of seeing blossoms light up their town. All tree species bear flowers of a sort, though the often colorful “cone flowers” of conifers lack ovaries and are technically called strobiles.
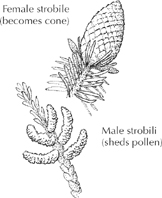
Conifer “Flowers”
Originating out of specialized buds, flowers are the first stage of the tree’s reproduction cycle. Basically, pollen grains from male flower parts called stamens fertilize the female eggs or ovules, which then develop into seeds within a fruit. But from the little perfumed blooms of the linden to the 60-million-blossom spray of the talipot palm, the varieties of flower design are staggering.
Trees arrange sexual roles in several ways. Many species bear flowers containing both male and female parts. Flowers with both parts plus petals and sepals (see below) are called complete flowers. Those with just the two reproductive parts are perfect flowers, perfect enough for nature’s purposes.
Species bearing male (staminate or pollen) flowers on one tree and females (pistillate) on another, as do ginkgoes, are called dioecious. The monoecious species, such as birches and most conifers, bear male and female flowers on the same tree. And if a tree bears both unisexual and bisexual flowers, it is called polygamous, which in the tree kingdom is legal.
In the course of evolution, a species usually fares better when each tree breeds with separate trees, rather than pollinating itself or reproducing by such self-contained asexual means as root sprouting. A mix of genes through cross-fertilization leads to mutations that are better adapted to environmental threats and changes—but it requires some not-always-present agents. Something must transport a tree’s pollen to the remote ovules of another tree. The agents are either wind or intermediaries such as insects and birds.
Big outputs of pollen and enlarged, sticky pollen receptors are typical of wind-pollinated trees such as oaks, poplars, ashes, and elms. Often their flowers are inconspicuous and appear before the leaves, which would block some of the pollen. Simple catkins or tassel-like clusters of minute flowers are often the simplest vehicles for pollen exchange.
To attract insects or birds, other species have developed dazzling, showy blossoms with abundant nectar and fragrance. Magnolias, catalpas, horse-chestnuts, and acacias provide some of the best flower shows in town. (See “Horse-Chestnut” profile for the love tryst between blossom and bug.)
The flower parts themselves wear many guises, depending on species, but have certain underlying features. The male flower organ is the stamen, typically a head (anther) of pollen on a stalk (filament). In nonconifers, the female pistil structure includes a sticky head (stigma) to receive pollen grains, and a stalk (style) connecting the stigma to the ovary. The ovules or eggs reside in the ovary, sometimes in chambers called carpels. When fertilized, they become seeds.

Flower (complete)
Among other flower parts—not always present and sometimes radically modified—are the corolla of petals; the calyx of sepals or green leaflike structures beneath the corolla; and the receptacle or flower base, which often becomes part of the fruit. When botanists cry peduncle, they are talking about the stalk that holds a flower or supports a bunch of flower stalks.
To avoid inbreeding, male and female flower parts of the same tree often mature at different times, or the female flowers grow high above the males (as on conifers), or the pollen is made incompatible with the ovules.
Fruit
To most people, fruit is what the supermarket sells at outlandish prices in its produce department. But, botanically speaking, the winged “whirlybird” of a maple tree is a fruit. So is a woody pine cone, or the dime-sized bib around an elm seed. The city teems with fruit on every leafy street.
A fruit is simply a plant’s seed-bearing part, developed from the fertilized flower. Botanists may distinguish the 20 or so different types—berry, capsule, nutlet, legume pods, etc.—by how the flower parts metamorphose into the fruit parts. (See “Crabapple” for one such metamorphosis.) The standard pattern is for the ovary to become the covering of the fertilized ovule or seed. Within the starchy flesh of edible fruits, one can often trace the former flower parts. But how did an 18-inch seed pod develop from the ovary of an orchidlike catalpa flower? What parts became the beaks of the spiny sweetgum burr?

Some fruit types
Matching fruit features to original flower parts can be edifying, but the first lesson for observers is to know a fruit when they see one. Apples (pomes), cherries (drupes), and acorns are familiar enough; more surprising are the bizarre cones arising from magnolias, the buttonlike gum nuts of a eucalyptus, or the small paddles dropping from ash trees. Many a new fruit will be met in the tree profiles, and many a familiar fruit revisited.
Not just flower parts, but flower formations (“inflorescences”), influence the look of a species’ fruits. Clustered blossoms on a hawthorn make for clusters of dangling berries (haws). Minute willow flowers on a stalk become a charm bracelet of jade seedfruit capsules. From a densely flowered catkin come the minifruits (“drupelets”) assembled into a single mulberry.
Seeds
Inside their ripening ovary, fertilized ovules take forms ranging from the gnatlike seeds of the birch tree to the edible coconut sphere. Like other tree parts, seeds have no lack of complexity to intrigue scientists. But for beginning tree observers, it may be enough to examine seeds, within their seed coats, as objects of amazement. A seed’s one sure trademark, if you can find it, is the belly-button-like scar (“hilum”) from where it was attached to the ovary wall. The dull circle on a shiny horse-chestnut seed is a large version of the hilum.
Having nurtured and protected its seeds, the ripened fruit now carries or releases them into the world, ideally at some distance from the parent to avoid competition and to expand the species’ territory. Timing is key. Some seeds, like those of willows, have only a few days of viability before they must germinate or die; others have years. Some require a period of dormancy before germination. Site or location, as they say in real estate, is everything. Seeds need amenable soil conditions, water and oxygen in the right temperature range, and the light-shade balance preferred by the species. With the odds long for such ideal conditions, trees tend to produce and disperse seeds in profusion.
How does the fruit help in dispersal? It may carry the seeds to their destinations, as when the gossamer-winged fruits (samaras) of the maple, elm, and ash drift off on the winds of spring and summer. Or the fruit may entice an intermediary to do the job by offering a sugary or starchy reward. Anyone who lives near a mulberry tree can see how birds have dispersed the undigested seeds in their purple droppings. Squirrels have been credited with “planting” some two-thirds of acorn seeds dropped by white oaks.
Seeds may be equipped with their own flight devices. Conifer seeds have insectlike wings. Willow, poplar, and sycamore seeds cruise long distances on feathery parachutes. Many cities know the summertime blizzards of cottonwood seeds that cover the streets and clog the window screens.
Protected by at least one coat, the seed settles into its new site. With the nutrients of its starchy “endosperm,” it supports the specialized cells that work themselves into embryonic plant parts. Soon the seed coat ruptures. The seed puts down a root. It bares its cotyledons, or embryonic “seed leaves.” It raises a stem. True leaves supplant the cotyledons. And a tree is born.