14. La parte sommersa dell’iceberg
Il costo energetico del vivente
In questo libro, abbiamo voluto esplorare due meccanismi fondamentali della costituzione del soggetto: la plasticità neuronale e i processi omeostatici. La plasticità è coinvolta nell’iscrizione dell’esperienza e, attraverso la riassociazione di tracce, produce, come abbiamo visto e in modo paradossale, una discontinuità. I processi omeostatici riguardano in primo luogo gli stati somatici S, identificati dai sistemi interocettivi che sono associati alle rappresentazioni R prodotte dalle assemblee dei neuroni. Come abbiamo visto, dalla tensione tra R e S risulta la pulsione che si scarica per mantenere l’omeostasi. Plasticità e processi omeostatici sono due meccanismi coinvolti in permanenza nel divenire specifico di ciascuno. Sono insiti nella materialità del vivente e da quel momento hanno ripercussioni concrete sul piano energetico. È l’occasione per rivisitare in modo nuovo la concezione freudiana che, al centro della questione del piacere pone dei parametri energetici, in cui l’aumento dell’eccitazione corrisponde a un dispiacere e la sua scarica a un piacere.
L’energetica è una questione di primo piano in biologia; è ugualmente centrale nella teoria psicoanalitica che si è cimentata nel distinguere il punto di vista topico (cioè quello delle istanze – inconscio, preconscio, conscio), il punto di vista dinamico (la dimensione conflittuale nella formazione dei sintomi) e, giustamente, il punto di vista energetico che, in Freud, è legato alla questione del piacere, per il tramite della scarica dell’eccitazione.
I processi mentali, così come appaiono per esempio nell’imaging cerebrale, non possono essere riportati unicamente all’attivazione da parte di stimoli provenienti dal mondo esterno, poiché non solo esiste un’intensa attività basale del cervello, che è il risultato, tra l’altro, del trattamento off line degli stimoli che riceve, ma anche di attivazioni provenienti dal corpo, dal vivente che il nostro cervello recepisce.1 Quest’ultimo punto è essenziale per il nostro discorso, poiché riguarda la questione del dispiacere – più ancora, dell’al di là del principio di piacere – associato all’eccesso del vivente.
Un fatto sorprendente della fisiologia del cervello è il suo grandissimo consumo di energia. Benché rappresenti solo il 2 per cento della massa corporea, il cervello consuma tra un quinto e un quarto dell’energia utilizzata dall’intero organismo.2-3 I neuroni sono forse più golosi dei muscoli? Quindi che facciamo, abbandoniamo le palestre e i centri di fitness? Basta con la puzza di sudore? Basta col jogging? Per restare magri, basterà pensare di più? Alleniamo le sinapsi ed ecco che i rotoli di grasso se ne vanno? Eh, purtroppo non è così, sarebbe troppo bello. Il nostro cervello consuma infatti 90 per cento delle proprie energie in una condizione detta «basale», cioè quando abbiamo gli occhi chiusi, le orecchie tappate e siamo stesi su un letto senza pensare a nulla. Fare un calcolo complicato, muovere una mano, declamare una poesia sono azioni che aumenteranno al massimo del 10 per cento – e solo localmente, per un tempo limitato alla durata dell’azione – il consumo di energia delle regioni cerebrali implicate in queste funzioni.4-5 Dimentichiamoci di dimagrire grazie all’esercizio mentale.
Di fatto, il nostro cervello è come un motore che gira in permanenza a 6000 giri al minuto e che, occasionalmente e per un tempo limitato, passa a 7000 giri. Per restare cosciente ha bisogno di un apporto energetico continuo: una piccola diminuzione di una piccolissima percentuale dell’afflusso sanguigno che gli porta il carburante, ovvero il glucosio e l’ossigeno, e si perde conoscenza. Ma a cosa serve questo consumo spropositato di energia? Molti studi hanno dimostrato che più dell’80 per cento è impiegato per il trasferimento di informazioni tra neuroni.6 Ricordiamo qui che i 100 miliardi di neuroni contenuti nel nostro cervello comunicano tra loro a livello delle sinapsi e che poiché ogni neurone è connesso ad altri attraverso circa 10000 sinapsi, esistono in tutto 1015 sinapsi a livello delle quali i nostri neuroni scambiano segnali.7-8
L’energetica del sistema nervoso, che viene chiamata neuroenergetica, ci illumina sui meccanismi cellulari che sono identificati dalle tecniche di imaging cerebrale.9 Queste nuove tecniche sono particolarmente utili per studiare i correlati neurobiologici della plasticità, dei processi omeostatici e i loro legami col binomio piacere-dispiacere. Conviene dunque vederli più da vicino, nei loro principi generali.
Tali segnali sono veicolati, a livello delle sinapsi, da molecole che sono chiamate neurotrasmettitori, il principale dei quali è il glutammato. Il glutammato liberato da un neurone esercita il suo effetto su altri neuroni (uno o più), interagendo con un recettore, un po’ come una chiave che apre una serratura (fig. 14.1).
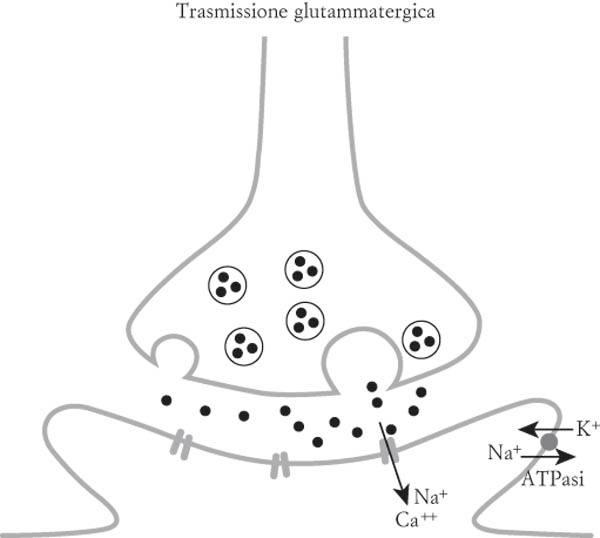
Figura 14.1
Rappresentazione schematica della neurotrasmissione mediata dal glutammato. Da notare il fatto che il sodio (Na) che entra nel neurone postsinaptico è rimandato all’esterno grazie all’attività di una pompa (ATPasi) che consuma energia. Altri meccanismi, che implicano anch’essi consumi energetici, mantengono l’omeostasi del calcio (Ca) che è anch’esso disturbato dalla trasmissione glutammatergica.
Apre così un canale, una «porta», attraverso la quale si infilano le cariche elettriche. La comunicazione tra i neuroni è dunque di natura chimica (neurotrasmettitori liberati a livello della sinapsi) ed elettrica all’interno del neurone (cariche elettriche). Diversamente da quanto accade con la corrente elettrica che si usa nelle nostre case, le cariche che portano la corrente non sono elettroni ma ioni. Per esempio il cloruro di sodio, costituito da uno ione cloro che porta una carica negativa e da uno ione sodio che porta una carica positiva. Quando il glutammato si lega al suo recettore, il sodio entra nel neurone bersaglio (in alcuni casi vi sarà anche un altro ione, il calcio, che porta anch’esso cariche positive) e genera il segnale elettrico. Poiché la concentrazione di sodio e di calcio è più elevata all’esterno che all’interno del neurone, questi ioni seguono una «pendenza» che li fa andare verso il compartimento dove la concentrazione è minore, ovvero l’interno del neurone. Si pone tuttavia un problema: poiché ogni neurone è bombardato in ogni momento da migliaia di molecole di glutammato che scatenano un’entrata di cariche positive, possiamo facilmente immaginare che questo gradiente verrà rapidamente a mancare e che questa «pendenza» che favorisce l’entrata del sodio e del calcio all’interno del neurone sparirà. Per fortuna esistono meccanismi che ristabiliscono questi gradienti rimandando regolarmente verso l’esterno del neurone gli ioni che sono entrati per effetto del glutammato. Mantenere questi gradienti, vuol dire mantenere la vita, mantenere i potenziali del vivente. Ed è là, a livello di questo processo che avviene il legame tra l’attività dei neuroni e il consumo di energia. Infatti questi meccanismi che ristabiliscono i gradienti consumano energia, molta energia, e sono sempre attivi. È come se una piccola nave imbarcasse acqua che il marinaio deve svuotare senza sosta a grandi secchiate per non affondare. Ecco cosa fanno continuamente le «pompe» presenti nei neuroni, che buttano fuori non acqua ma ioni.
Come tutti i processi cellulari, queste pompe utilizzano ATP (adenosintrifosfato) come carburante. L’ATP è fabbricato dalle cellule a partire dal glucosio e dall’ossigeno portati dal flusso sanguigno. Secondo le stime fatte a partire da dati sperimentali, il costo energetico di una sinapsi sarebbe pari a 3 × 109 molecole di ATP per neurone per secondo, il che implica un consumo energetico considerevole. L’attività elettrica e chimica di ogni neurone, alla base del processo di elaborazione del segnale e di trattamento dell’informazione da parte del cervello, è dunque un processo costoso in termini energetici, che è utilizzato in un certo qual modo per «ricaricare» continuamente le «batterie» dei neuroni.
Ma perché tutti questi dettagli? Perché il glucosio e l’ossigeno, a partire dai quali viene fabbricato l’ATP necessario al mantenimento dell’attività dei neuroni, sono per l’appunto i due parametri che vengono misurati dalle tecniche di imaging cerebrale e che permettono di visualizzare il vivente all’opera. Lo scopo di questo capitolo è infatti quello di far dialogare il piacere con i meccanismi che lo sottendono. Non si tratta, invece, di ridurre il piacere a questi meccanismi, non più di quanto si ridurrebbe l’amore ai meccanismi somatici che gli vengono associati. Non si tratta di fare una neofrenologia del piacere con una cartografia delle aree implicate. Si tratta piuttosto di cogliere i meccanismi cellulari in gioco. Possiamo accumulare immagini di attività cerebrali durante il piacere o il dispiacere: non ci daranno informazioni sui processi cellulari che sono alla base. Non è facendo una mappatura che capiremo l’implicazione del vivente nel piacere.
Il principio di costanza e il principio d’inerzia propri della teoria freudiana potrebbero essere rivisitati alla luce di questi dati. In effetti, in questi principi si ritrova la nozione di costo energetico del vivente e di processi omeostatici che mirano a ristabilire un livello basale costante. Vi si apprezza anche la natura dinamica dei processi propri del vivente.10 Nella teoria freudiana, il piacere corrisponde alla scarica dell’eccitazione. Considerando ciò che abbiamo visto in precedenza, possiamo dire che il costo energetico del piacere risiede piuttosto nel ristabilire le condizioni che permettono la scarica. Infatti ciò che costa, in termini cellulari, non è l’attività del neurone, ma piuttosto il mantenimento della potenzialità di essere attivo, dunque il ristabilirsi di condizioni che permettono l’eccitabilità del neurone. È la ricarica, piuttosto che la scarica, che costa energia. In termini freudiani, sono i principi di costanza (mantenimento dell’eccitabilità) e di inerzia (ristabilimento dell’eccitabilità secondo la neuroenergetica) che costano energia.11 Siamo consapevoli della diversità dei piani logici che stiamo mettendo in relazione, con da una parte il costo energetico che permette la scarica dell’attività neuronale e, dall’altra, l’esigenza di una scarica dell’eccitazione tipica della realizzazione del piacere nella concezione freudiana. Ciò che conta, per noi, è indicare la natura dinamica dei processi energetici che sono in gioco in tutti e due i piani logici. Ci sembra di poter essere autorizzati ad arrischiarci nell’ipotizzare un tale legame grazie a questa concezione dinamica dell’energetica e del piacere. Non si tratta di fornire delle spiegazioni definitive, ma piuttosto di iniziare a ragionare attorno all’intuizione freudiana dei principi di costanza e di inerzia mediante le procedure proposte dalla biologia contemporanea e che riguardano i meccanismi omeostatici. È la ricarica che costa. In ogni caso la neuroenergetica contemporanea combacia con le intuizioni e i ragionamenti freudiani in modo inatteso, per andare incontro agli enigmi del piacere.
Un altro aspetto stupefacente dell’attività cerebrale è il fatto che i neuroni sono attivi anche quando il cervello è in uno stato detto «basale», cioè quando il soggetto non riceve alcuno stimolo sensoriale e non è coinvolto in alcuna attività motoria o di pensiero. Sono le tecniche di imaging cerebrale funzionale che ci danno informazioni su questo aspetto. Tutti hanno oggi familiarità con le immagini prodotte da queste tecniche, che sia la tomografia a emissione di positroni (PET) o l’imaging mediante risonanza magnetica funzionale (fMRI), che permette di visualizzare le regioni cerebrali attive in un dato compito sensitivo, motorio o astratto. Se pronunciate delle parole, la PET mostrerà, per esempio, l’attivazione di una regione del lobo temporale, l’area di Broca; questa area si «accenderà» segnalando così il suo coinvolgimento nel compito. Ciò che forse è meno noto ai non specialisti è che queste tecniche non individuano l’attività elettrica o chimica dei neuroni ma il consumo di energia legato al loro «lavoro», cioè al lavoro delle «pompe» che ricaricano le batterie neuronali.12 L’essenziale è innanzitutto il mantenimento di una costanza, che è ciò che fa la vita. La scarica ha certamente luogo, ma il costo energetico è legato al ritorno dell’omeostasi cellulare. È anche una delle versioni del mantenimento della vita il fatto che la materia propria del vivente sia «animata».13 Come il lavoro muscolare è accoppiato all’aumento del flusso sanguigno e della disponibilità di substrati energetici sotto forma di glucosio e di ossigeno, così l’attivazione dei neuroni induce il consumo di una maggiore quantità di energia. Sono segnali legati al consumo energetico – dunque di glucosio e di ossigeno rilasciati dal flusso sanguigno – che vengono individuati dalle tecniche di imaging.14
Come abbiamo visto prima, il cervello non è attivo solo quando svolgiamo un compito motorio, cognitivo o quando proviamo una sensazione, un’emozione che può essere visualizzata dalle tecniche di imaging funzionale nel corso di paradigmi detti di attivazione. Questo approccio, che tanto ha contribuito al progresso delle conoscenze – in particolare nell’ambito delle neuroscienze cognitive, mediante l’identificazione di regioni particolarmente implicate in questo o quel processo mentale – utilizza giustamente dei protocolli di sottrazione. Così si sottraggono, mediante appropriati algoritmi, le immagini ottenute in condizioni «basali» da quelle ottenute durante l’attivazione, mettendo in evidenza le regioni implicate nel paradigma esaminato.
Gli aumenti locali di flusso sanguigno, di consumo di glucosio o di ossigeno associate all’attivazione che è identificata dalle tecniche di imaging funzionale corrispondono a cambiamenti nell’ordine massimo del 10 per cento. Resta dunque l’enigma del consumo di energia di base da parte del cervello, che corrisponde al 90 per cento del consumo totale. È forse questa una traccia per cogliere l’enigma del piacere?
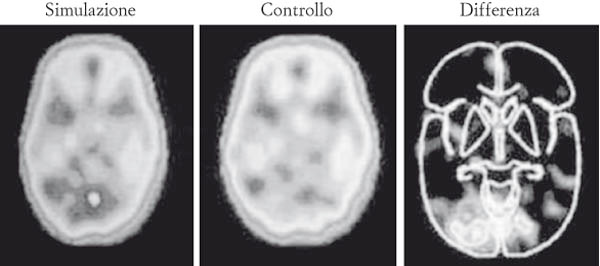
Figura 14.2
Le immagini prodotte dalle tecniche di imaging cerebrale funzionale sono il risultato di processi di sottrazione di un’attività basale molto intensa, la «cima dell’iceberg».
Quali sono dunque i meccanismi che operano in condizioni basali e qual è la loro funzione? Si possono evocare diverse possibilità per rendere conto di questa importante attività basale. Torniamo per un attimo alla comunicazione tra neuroni. Abbiamo parlato del ruolo del glutammato come molecola di segnale a livello della sinapsi. Facendo entrare le cariche positive (sodio e calcio), il glutammato attiva i neuroni – in termini neurofarmacologici possiamo dire che il glutammato è liberato a livello di sinapsi eccitatorie che costituiscono circa l’80 per cento delle sinapsi del sistema nervoso. Come nella maggior parte dei sistemi biologici, un «acceleratore» (l’effetto eccitatorio del glutammato) è controbilanciato da un «freno». Qual è dunque il freno dei neuroni? Si tratta di un altro neurotrasmettitore liberato dal 15 per cento delle sinapsi del cervello, l’acido gamma-amminobutirrico o GABA. Come il glutammato, anche quest’ultimo agisce su recettori specifici (le chiavi) che aprono dei canali (le porte) permeabili a uno ione, il cloro, che porta con sé cariche negative che possono dunque controbilanciare le cariche positive portate dal sodio e dal calcio. Ogni neurone del sistema nervoso riceve continuamente segnali eccitatori e inibitori nelle sinapsi che utilizzano, rispettivamente, il glutammato e il GABA. Questi segnali sono integrati a livello di ciascun neurone: se predomina l’eccitazione, il primo neurone ne attiverà altri che fanno parte dello stesso circuito di neuroni; se, al contrario, predomina l’inibizione, l’attività globale di un dato circuito neuronale diminuirà. L’attività globale di una rete di neuroni risulta da un sottile equilibrio tra eccitazione e inibizione.15 Prendiamo il caso frequente in cui l’attività inibitrice predomina: il circuito interessato non sarà attivato dal punto di vista elettrico ma, paradossalmente, lo sarà dal punto di vista del consumo di energia. In effetti, i gradienti ionici del sodio, del calcio e del cloro saranno stati dissipati dal glutammato e dal GABA rilasciati simultaneamente presso le sinapsi eccitatrici e inibitrici che convergono sui neuroni del circuito. Le «pompe» a ioni si attiveranno e consumeranno energia anche in assenza di un segnale elettrico. Potremmo qui riprendere l’analogia del motore di un’automobile. Immaginiamo che voi siate al volante e che spingiate a fondo e simultaneamente sui pedali del freno e dell’acceleratore: la vostra macchina non andrà avanti ma consumerà benzina. Dove sta l’interesse, direte voi? Sta nell’ottimizzare la velocità di esecuzione. Nelle corse automobilistiche questa modalità è utilizzata regolarmente nelle gare in cui rutilanti veicoli a motore stanno immobili sulla linea di partenza, con i freni bloccati e l’acceleratore a fondo; lo scopo è di realizzare l’accelerazione più rapida possibile su un centinaio di metri, dopo aver «mollato» il freno. Allo stesso modo, per i circuiti neuronali l’equilibrio tra eccitazione e inibizione, tra «freno» e «acceleratore» permetterebbe di mobilizzare istantaneamente un circuito neuronale togliendo l’inibizione. Il prezzo da pagare per questa disponibilità sarebbe il costo energetico elevato in condizione basale.16
Ma questo equilibrio dinamico permanente tra eccitazione e inibizione fornisce solo una spiegazione parziale per il consumo energetico in condizioni basali e altri meccanismi possono essere evocati, in particolare quelli in relazione col sistema di funzionamento del cervello che riusciamo a vedere grazie all’imaging. Innanzitutto, il modo dell’attivazione: il cervello è in quel caso on line sia per uno stimolo proveniente dal mondo esterno sia per un’attività motoria sia ancora per un’operazione mentale compiuta dal soggetto, e l’imaging rivela le regioni cerebrali coinvolte in questi compiti sotto forma di un segnale di attivazione. Come l’esperienza, che sia sensitiva, motoria o puramente mentale, lascia una traccia nel circuito neuronale grazie ai meccanismi di plasticità, così potremmo definire il processo di plasticità come corrispondente a una modalità di funzionamento cerebrale off line che implica dei processi avviati dalla percezione ma scaglionati nel tempo, e che si sviluppano in seguito a uno stimolo anche quando questo è cessato: l’informazione continua a essere elaborata off line.
I meccanismi di plasticità neuronale mettono in gioco una serie di processi cellulari e molecolari legati, tra l’altro, all’attività dei canali ionici, all’attivazione di enzimi, alla regolazione di diversi geni e anche a modificazioni microstrutturali delle sinapsi.17 E c’è ragione di credere che questi processi legati alla plasticità sinaptica abbiano un costo energetico proprio e che è indipendente dai costi legati all’attività on line del cervello. È importante segnalare qui che certe regioni del cervello sono più attive quando non sono coinvolte in alcuna stimolazione o attività che coinvolga una modalità particolare. Queste regioni sembrano costituire una rete particolarmente attiva quando il cervello è off line (nessuno stimolo sensitivo, nessuna attività motoria o cognitiva). Questo circuito è stato definito come default mode network, una sorta di attività per «difetto», cioè basale, del cervello.18 Il significato di questa modalità di attività resta ancora un enigma, ma è forse legato ai meccanismi di plasticità sinaptica?
Così il nostro cervello è modificato continuamente dall’esperienza, e mette in gioco meccanismi di plasticità che partecipano al consumo energetico in condizioni basali, nella misura in cui questi meccanismi operano in permanenza e al di fuori dell’attivazione on line del cervello. L’immagine che è spesso utilizzata per illustrare questa attività basale off line è quella dell’iceberg. In questa immagine, l’attivazione rappresenta solo la punta dell’iceberg del consumo energetico, mentre la parte sommersa, molto più importante, rappresenta il consumo basale. C’è di più, poiché il nostro cervello non riceve stimoli solo dal mondo esterno, mediante quello che la neurofisiologia definisce come sistema esterocettivo (per esempio, i sistemi visivo, uditivo, olfattivo, somatosensoriale); vi è anche il sistema interocettivo e il nostro cervello è «bombardato» in permanenza da stimoli che provengono dall’interno del corpo, a partire dai visceri (per esempio, il sistema digestivo, il sistema cardiovascolare), dei muscoli e delle articolazioni. Senza contare il livello di ormoni e di altre molecole che, presenti in circolo, sono identificate in ogni istante.19
Così il nostro cervello riceve in permanenza informazioni sullo stato del corpo – è ciò che noi abbiamo chiamato la lettura da parte del cervello degli «stati somatici». Il corpo, il vivente, ci «parla» dunque continuamente, anche se non lo sentiamo, poiché le percezioni veicolate dal sistema interocettivo rimangono per lo più inaccessibili alla coscienza. Chi ha coscienza dello stato della propria milza o del proprio colon ascendente? Per quel che riguarda la vescica, ne abbiamo coscienza solo quando è piena e niente più. Nonostante ciò qualsiasi percezione, cosciente o non, attiva il sistema nervoso e fa aumentare il consumo di energia di conseguenza. Ci pare dunque ragionevole postulare che questa attività permanente del sistema interocettivo comporti un costo energetico che interviene nel consumo basale di energia da parte del cervello. Per riprendere la nostra immagine, in termini energetici la parte sommersa dell’iceberg sarebbe prodotta dai processi neuronali off line legati alla plasticità neuronale, all’interocezione e ai processi omeostatici sottesi.
Questi processi entrano in gioco nella costituzione dell’inconscio freudiano – Unbewusst. Noi proponiamo che lo studio della dinamica dell’attività basale cerebrale, così come ci viene mostrata dall’imaging, possa essere il modo per esplorare l’inconscio all’opera nel corso di una terapia analitica. Si tratta proprio, in effetti, di una dinamica. L’inconscio può essere visto non solo come un sistema di tracce che si impone in modo determinate, ma come il movimento di una discontinuità dinamica che apre su ciò che non è ancora realizzato, e di cui lo studio dei costi energetici associati all’attività basale potrebbe essere il riflesso.