
|128|Abschnitt der Trias-Jura-Grenze auf den Five Islands, Nova Scotia, Kanada.
IN KER UNTERTRIAS WAR DIE BIOSPHÄRE von wenig vielfältigen kosmopolitischen Taxa besiedelt. Dies war auch bei jedem der zuvor besprochenen Intervalle, die einem Aussterben folgten, der Fall gewesen. Aber ebenso wie das Biomer-Sterben im späten Kambrium ging das Ereignis am Ende des Perms einem grundlegenden Wandel der Biota der Erde voraus. Vom frühen Ordovizium bis zum Ende des Perms war das Leben auf der Erde von der paläozoischen evolutionären Fauna dominiert gewesen, einer ökologisch losen marinen Gemeinschaft von Anthozoen (Korallen), artikulierten Brachiopoden (Armfüßer), stenolaematen Bryozoen (Moostierchen), cephalopoden Mollusken (Kopffüßer), stellaroiden und crinoiden Echinodermaten (Stachelhäuter) und kleinen, garnelenähnlichen Ostrakoden (Muschelkrebse). Die meisten fossilen Meeresfaunen dieses Zeitabschnitts bestehen rund zur Hälfte aus diesen Gruppen. Über der Perm-Trias-Grenze halbiert sich jedoch die Zahl der Gattungen, die diesen taxonomischen Gruppen zugeordnet werden, und im Mesozoikum stellen sie durchschnittlich weniger als ein Drittel der Meeresgemeinschaften. Heute bestehen nur noch knapp über zehn Prozent der modernen Meeresorganismen aus Spezies dieser Gruppen.
Beginnend mit der frühen Trias wurde der Platz der paläozoischen evolutionären Fauna von taxonomischen Gruppen eingenommen, mit denen wir heute besser vertraut sind, darunter kalkhaltiger Nanoplankton, Dinoflagellatenplankton und Foraminiferen (benthische und planktonische), Kieselalgen, Schwämme, deren Skelette aus Spongin, Siliziumdioxid oder beidem bestehen (Demospongien), Moostierchen mit einer fortgeschrittenen Muskelkonfiguration zum Hinausschieben des Fressorgans (Gymnolaematen), Schnecken (Gastropoden) und zweischalige |130|Weichtiere (Muscheln), die keksförmigen Seeigel (Stachelhäuter), marine krebsartige Arthropoden (Garnelen, Hummer, Krill), Haie (Knorpelfische/Chondrichthyes), Meeresreptilien und Knochenfische (Osteichthyes, siehe oben). Im terrestrischen Bereich lässt sich diese Liste durch Insekten, Frösche, Schildkröten, Eidechsen, Flugsaurier, Archosaurier (darunter Krokodile, nichtvogelartige Dinosaurier und Vögel) sowie Säugetiere ergänzen. In den Sedimenten, die unmittelbar über der Perm-Trias-Grenze liegen, wurden die Mitglieder der paläozoischen Fauna zahlenmäßig von Vertretern dieser Gruppen abgelöst, die daraufhin ihren unaufhaltsamen Siegeszug antraten und bis heute zu einer überwältigenden Mehrheit von 90 Prozent aller lebenden Arten anwuchsen. In der Welt der Pflanzen waren die Veränderungen an der Perm-Trias-Grenze, wie wir bereits bei früheren Aussterbeereignissen beobachten konnten, deutlich weniger ausgeprägt als in der Welt der Fauna. Die sogenannte mesophytische Flora bestand bis in die Untertrias fort, einschließlich Bärlappen, Nadelbäumen, Palmfarnen und Ginkgos. Allerdings erscheinen Farne erst in der Trias im fossilen Befund und sind seitdem ein bedeutender Bestandteil der Flora.

Rekonstruktion der mitteltriassischen Meeresumwelt in der Schweiz.
Vordergrund: der riesige Ichthyosaurier Shastasaurus (links), der Elasmosaurier Tanystropheus, der sich durch einen extrem verlängerten Hals auszeichnet (unten links), der Nothosaurier Cerisiosaurus (Mitte), der Placodontier Paraplacodus (marines Reptil, unten Mitte), der Strahlenfosser Saurichthys (unten Mitte) und Ammoniten (Cephalopoden, unten Mitte).
Während die biotische Welt der Trias von Organismen mit bekannten Körperbauplänen besiedelt wurde, war die physische Umwelt im Vergleich zur heutigen kaum wiederzuerkennen. Pangäa blieb zwar intakt, begann aber eine Bewegung in nördlicher Richtung, wodurch sich die miteinander verschmolzenen gondwanischen Kontinente von Südamerika, Afrika, Antarktika und Indien vom Südpol entfernten. Im Gegenzug schoben sich Sibirien und Kasachstan über den Nordpol. Als Pangäa mit einer Drehung im Uhrzeigersinn nach Norden driftete, schloss sich |131|langsam die Tethys, wobei an den nördlichen und östlichen Meeresrändern Tiefseegräben entstanden. Dies führte zur Bildung und fortschreitenden Verschmelzung von Inselbögen entlang der nördlichen und östlichen Tethysränder. Diese wachsenden Landmassen würden später einmal von der Türkei, Iran, Tibet, China, Ostasien und Indonesien aufgenommen werden, existierten zu dieser Zeit aber noch als kaum aufgetauchte Inseln oder vollständig untergetauchte ozeanische Plateaus. Wichtiger für ein Verständnis der triassischen Erde ist vielleicht, dass die Positionen dieser Landmassen – die infolge der tektonischen Prozesse entstanden – die ozeanischen Zirkulationsmuster sowohl innerhalb der Tethys als auch zwischen Tethys und Panthalassa beeinflusst haben dürften, wobei sich das Tethysbecken im Verlauf der Trias wahrscheinlich zunehmend isoliert hat.
Obertriassische terrestrische Umwelt in Kirgisistan.

Vordergrund: die Käfergattungen Notocupoides (unten links) und Hadeocoleus (Mitte links), das frühe Gleitreptil Sharovipteryx (Mitte), die Insektengattung Gigatitan (Titanoptera, oben rechts). Hintergrund: der Strahlenflosser Saurichthys (oben Mitte), das langschuppige (oder protogefiederte) Reptil Longisquama (Mitte) und das synapside Reptil Madygenia (oben Mitte).
Während der Trias stieg der Meeresspiegel an – möglicherweise im Zuge einer verminderten Vereisung in den hohen Breitengraden, aber höchstwahrscheinlich aufgrund der tektonisch bedingten Änderungen der Ozeanbeckenvolumen. Ein leichter glazialeustatischer Anstieg des Meeresspiegels wäre infolge der Wegbewegung Pangäas vom Südpol zu erwarten, aber für große Eisschilde in mittleren Breitengraden gibt es im späten Perm nur wenige direkte Beweise und in der frühen Trias überhaupt keine. Aktuelle paläogeografische Rekonstruktionen von Pangäa in der Trias legen nahe, dass in der Nähe des Nordpols breite Meereseinbuchtungen vorhanden waren. Diese könnten die Intensität |132|der Gletschertätigkeit in der nördlichen Hemisphäre abgemildert haben. Unabhängig davon gibt es genügend Hinweise auf die Ablagerung von marinen Sedimenten auf den triassischen kontinentalen Plattformen in Australien, Indien, Arabien und im östlichen Nordamerika, die zu dieser Zeit alle an den südlichen und westlichen Rändern der Tethys lagen. Die westlichen Ränder von Nord- und Südamerika und Antarktika waren in der Trias durch die Subduktion der ozeanischen Platten und die damit einhergehende Entstehung von Bergen geprägt, wodurch die Festlandsockel in diesen Regionen ziemlich schmal blieben.
Paläogeografe der frühen Trias
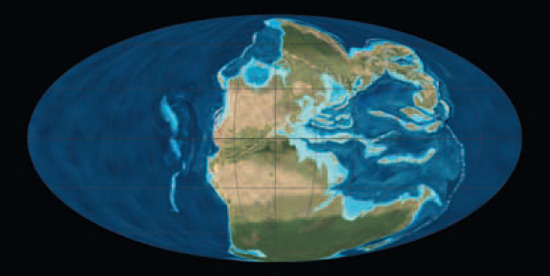
(vor ca. 250 Millionen Jahren) und der späten Trias
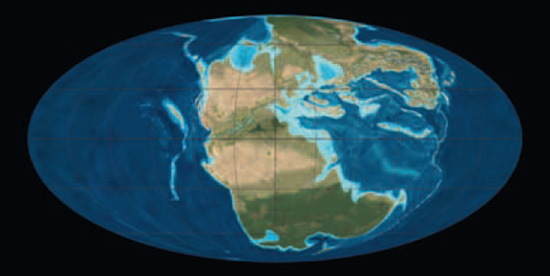
vor ca. 220 Millionen Jahren), mit den Positionen der kontinentalen Landmassen und Ozeanbecken.
In der späten Trias kehrte sich dieser Trend jedoch um. In ganz Europa und der nordamerikanischen Arktis gibt es unzählige Beweise für eine erhebliche und geografisch ausgedehnte Absenkung des Meeresspiegels. Die Beweise für diese Regression sind vielfältig und umfassen marine Hiatus mit Erosionsbereichen, die Exposition von Riffen und die ausgeprägte Verflachung der Umgebung von Karbonatplattformen. Noch bedeutender ist nach dieser regressiven Phase die Fortsetzung der marinen Ablagerungen, oft in Form von dysaerobem/anaerobem Schwarzschiefer, der die Ausbreitung von sauerstoffarmem Wasser aus den Tiefseeregionen über die Festlandsockel anzeigt. Wie wir bereits gesehen haben, ist diese regressiv-transgressive/dysaerobe Kombination oft mit einem umfassenden marinen Artensterben verbunden. Der oder die indirekten kausalen Mechanismen für diesen starken regressiv-transgressiven Impuls sind unklar, doch scheint dieser Zeitabschnitt mit den frühen Phasen der tektonischen Grabenbildung zwischen Nordamerika, Südamerika, Afrika und Europa verknüpft zu sein, die einmal den Atlantischen Ozean bilden werden.
Im Vergleich zu den extremen Klimabedingungen im späten Perm war das Klima in der Trias eher gleichmäßig. Im Verlauf dieses Zeitraums zog sich die Zone extremer Aridität – abgesteckt durch die Orte ausgedehnter Evaporitablagerungen – zum triassischen Äquator zurück, was von der Entwicklung breiter warm-gemäßigter und tropischer Regionen begleitet war, in denen sich Kohlesümpfe bildeten. Besonders in der nördlichen Hemisphäre gab es viele Zonen der Kohlebildung. Trotzdem |133|fanden sich, wie Calcret- und Evaporitablagerungen anzeigen, entlang der nordwestlichen Ränder der Tethys auch weiterhin aride Bedingungen, in Gegenden, die heute an den westlichen Rändern von Europa, Skandinavien und Ostgrönland liegen.
In dieser triassischen Welt brachte die Evolution mehrere interessante Innovationen in der Geschichte des Lebens hervor. In ökologischer Hinsicht waren die modernen Phytoplankton-Gruppen am wichtigsten: kalkhaltiges Nannoplankton und Dinoflagellaten. Zusammen mit den Kieselalgen, deren Radiation später stattfand, stellen diese Gruppen im gesamten Mesozoikum, und noch bis heute, die Grundlage für marine Nahrungsketten. Als solche spielen sie allerdings nicht nur in Bezug auf die Meeresökologie eine entscheidende Rolle, denn Phytoplankton sitzt auch – oder besser schwebt – am kritischen Nexus zwischen dem physikalischen und dem organischen Bereich. Da diese Gruppen schlagartig im Fossilienbestand erscheinen, könnten sie auf eine lange, bisher unsichtbare spätpaläozoische Evolutionsgeschichte zurückblicken.
Im seichten Meeresgewässer schlugen sich die Brachiopoden, zweischaligen Weichtiere und Ammoniten schnell einen evolutionären Vorteil aus den entvölkerten Lebensräumen. Jede dieser Gruppen durchlief in der frühen Trias recht bemerkenswerte adaptive Radiationen. Im Gegensatz zu den Phytoplankton-Radiationen, die eventuell eine lange vortriassische Geschichte hatten, ist bekannt, dass alle triassischen Brachiopoden und Weichtiere von sehr wenigen Überlebenden der Artensterben am Ende des Perms abstammen. Auch Riffe erschienen zur mittleren Trias wieder in der Tethys und wuchsen in vielen tropischen und warm-gemäßigten Regionen an der nördlichen und östlichen Küste der Tethys zu beeindruckenden Gebilden heran. Eine wichtige Ergänzung erfuhr dieses Biom durch das Auftreten der modernen Korallen, die heute die meisten modernen Riff-Ökosysteme in den Ozeanen dominieren.
Die bei Weitem erstaunlichsten evolutionären Entwicklungen in der Trias fanden jedoch unter den Wirbeltieren statt. Knochenfische, Haie und Rochen entwickelten sich alle in den Ozeanen weiter, mussten diesen ökologischen Raum aber zunehmend mit luftatmenden Eindringlingen teilen. Pflasterzahnechsen (Placodonten) und robbenähnliche Reptilien ernährten sich von den reichlich vorhandenen Muscheln und Nothosaurier mit nadelscharfen Zähnen jagten Fische in den flachen Meeren der Trias. Die Spitzenräuber im Meer der frühen Trias waren die fischähnlichen Ichthyosaurier. An Land stellten die säugetierähnlichen therapsiden Reptilien eine Erfolgsgeschichte der frühen Trias dar, sie wurden am Ende dieser Periode allerdings von den kleineren, grazileren und schnelleren thecodonten Reptilien ersetzt, darunter die beliebtesten |134|Nachfahren der Thecodontia: die Dinosaurier. Obwohl Dinosaurier im triassischen Fossilienbestand zuerst als Tetrapoden (Vierfüßer) von der Größe kleiner Hunde erschienen, hatte ihre Evolutionslinie zur Zeit der Trias-Jura-Grenze Giganten von über sechs Metern Länge hervorgebracht. Ebenfalls traten in der Trias erstmals echte Säugetiere auf, sie erschienen jedoch während des gesamten Intervalls als seltene, kleine, nagetierartige Spezies, die sich von Insekten oder Aas ernährten. Trotz der offensichtlichen Erfolge dieser neuen, modern aussehenden Arten waren die guten Zeiten der Trias von kurzer Dauer. Schon 25 Millionen Jahre nach dem verheerenden Massenaussterben am Ende des Perms wurde die Erde erneut in ein Umweltchaos gestürzt, aus dem nur ein kleiner Bruchteil der marinen und terrestrischen Arten wieder auftauchen würde.
Obwohl das Massensterben am Ende der Trias das zweitjüngste der „großen Fünf“ ist, wissen die Geowissenschaftler seltsamerweise wohl am wenigsten darüber. Zu einem gewissen Grad ist dies einem Mangel an gut dokumentierten und vollständigen geologischen Schnitten und Bohrkernen der Trias-Jura-Grenze geschuldet, was sich wiederum einem massiven globalen Rückgang des Meeresspiegels an dieser Grenze verdankt. Doch wie wir gesehen haben, ereigneten sich auch alle bisherigen Massensterben, abgesehen vom Oberdevon, zu Zeiten eines signifikanten Meeresspiegeltiefstands. Der andere Faktor, der Rückschlüsse erschwert, ist ein relativer Mangel akkurater Datierungsmöglichkeiten der Triassedimente, zumindest im Vergleich zu jenen Datierungsmethoden, die für das darüber liegende Jura, die Kreidezeit und das Känozoikum zur Verfügung stehen. Schätzungen zur Größenordnung des Massenaussterbens |135|am Ende der Trias stimmen darin überein, dass mehr als 20 Prozent der marinen und terrestrischen Tierfamilien und über 50 Prozent der damals vorhandenen Gattungen zu diesem Zeitpunkt den Fossilienbestand verlassen. Simulationen zeigen, dass dies mehr als 75 Prozent aller fossilisierbaren Arten entspricht, womit die Aussterbeintensität ungefähr auf demselben Niveau liegt wie am Ende des Devons.

Der Lystrosaurus war ein Vertreter der Dicynodonten, ein säugetierähnliches Reptil, das vor ca. 252– 247 Millionen Jahren in der unteren Trias lebte. Seine Fossilien wurden in Südafrika, Indien, Antarktika, China und Russland gefunden.
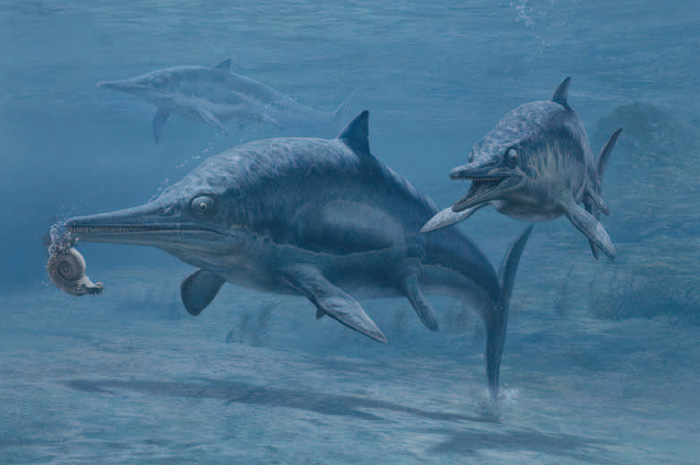
Ein Ichthyosaurus grendelius mit Jungtier isst einen Ammoniten. Ichthyosaurier lebten während des Mesozoikums und starben vor rund 90 Millionen Jahren in der Kreidezeit aus.
Abermals wurden in der späten Trias Riff-Ökosysteme dezimiert. Im Anschluss an das Aussterben am Ende des Perms entwickelte sich in der unteren Trias eine „Riff-Lücke“, die sich erst in der Mitteltrias mit der Entwicklung niedriger Hügel aus Stromatoporen, Kalkschwämmen und Krustenalgen wieder füllte. Steinkorallen tauchen in der Mitteltrias auf und waren die dominierenden Riffbildner im Norium, einer Zeitperiode, die von Riff-Paläobiologen als „Morgendämmerung der modernen Riffe“ bezeichnet wird. Allerdings muss die zunehmende Dominanz der Steinkorallen im Zusammenhang mit sinkenden Riffvorkommen und einer schwindenden Vielfalt in der gesamten mittleren und oberen Trias gesehen werden. Am Ende des Rhaetiums war das Riff-Biom fast völlig verschwunden, und mehr als 95 Prozent der rhätischen Korallen verließen den Fossilienbestand. Die letzten verbleibenden triassischen Riffe fanden sich in der Tethys. Mit ihrem Verschwinden hörten alle Riff-bedingten Karbonatablagerungen in dieser Meerespassage auf, bis sich |136|im frühen Jura erneut einige wenige kleine Riffe zu formen begannen.
Prominente Opfer des Massenaussterbens am Ende der Trias:

Ammoniten (Cephalopoden, Ceratites nodosus aus Deutschland).

Nothosaurier (Neusticosaurus pusillus aus Basano, Lombardei, Italien).
Das Verschwinden der Riffe steht möglicherweise im Zusammenhang mit einem Rückgang der marinen Primärproduzenten, eine Hypothese, für die unter anderem das Aussterben der Zooplanktongruppen (z.B. Radiolarien) spricht. Muscheln waren besonders stark betroffen und verloren mehr als 90 Prozent ihrer Arten. Brachiopoden und Ammoniten erlitten Verluste von 80 beziehungsweise 60 Prozent ihrer Arten. Die Ammonitenzahlen sind besonders bemerkenswert, da sie ein klar fortschreitendes Aussterbemuster zeigen: Von 150 Ammonitengattungen im Karnium sind im Norium noch 90 vorhanden und in der obersten Stufe der Trias, dem Rhaetium, nur noch sechs oder sieben. Der Ammonitenbefund zeigt ebenso Hinweise auf eine differenzierte Selektion zum Nachteil großer, kompliziert gewundener und (wahrscheinlich) ökologisch spezialisierter Formen. Innerhalb des Mikroplanktons erlitten die Radiolarien einen Verlust von etwa 69 Prozent der Gattungen, doch seltsamerweise starben nur wenige Familien aus. Bemerkenswerter ist der Verlust ganzer Organismengruppen, deren größte die Conodonten sind – ihre letzten Repräsentanten verlassen den fossilen Befund an diesem Punkt unwiderruflich, wie auch die meisten Labyrinthodonten (Amphibien), säugetierähnlichen Reptilien und viele thecodonte Reptilien, darunter eine große Zahl an Dinosauriergattungen und -arten.
|137|Prominente Opfer des Massenaussterbens am Ende der Trias:

Korallen (Astraemor sp.).
Conodonten (Hadrotontina sp.).

Das placodonte Reptil Placochelys placodonta aus Veszprém, Bakony, Ungarn.

Die Amphibie Benthosuchus sushkini vom Scharschenga-Fluss, Vachnevo, Russland.
Die Schätzungen zum Zeitablauf des obertriassischen Aussterbens gehen auseinander. Raups und Sepkoskis Daten – die im Wesentlichen von marinen Fossilien stammen – verorten den Großteil der Extinktionen im Norium, während Mike Bentons Datensammlungen für terrestrische Wirbeltiere für ein Zwei-Stufen-Ereignis sprechen, das auch die vorherige stratigrafischen Stufe, das Karnium, umfasst. Diese letztere Interpretation hat durch neuere Untersuchungen der Meeresdaten an Popularität und Unterstützung gewonnen.
Ein Mangel an obertriassischen Sedimenten, die für Paläontologen zugänglich sind, hat das geografische Verständnis dieses Ereignisses erschwert. Nimmt man Riffe als Modell, scheint es allerdings plausibel zu sein, dass tropische Regionen am stärksten betroffen waren. Diese Spekulation stimmt nicht nur mit den Verteilungsdaten der Riffe überein, sondern steht auch für eine Erwartung, die sich an das gut dokumentierte und fast steile Absinken des Meeresspiegels knüpft, das die späte Trias auszeichnet. Marginale küstennahe Lebensräume in der Tethys stellen die letzten Refugien der triassischen Riffgemeinschaften dar. Am Ende dieser Periode waren auch diese aus dem Fossilienbestand verschwunden.
Die Muster der Meeresspiegelregression in der Obertrias werden noch sehr kontrovers diskutiert. Einige Geologen glauben, dass es eine schleichende Regression war, die in der Mitteltrias begann. Andere argumentieren dafür, dass es ein scharf umrissenes Ereignis war, das sich auf die oberste Trias beschränkte und im frühen Jura fortsetzte. Wieder andere sehen für Westeuropa eindeutige Hinweise auf einen Meeresspiegelabfall, aber nicht für andere Regionen. Auch hier fehlen die Sedimente aus der letzten Phase der Trias, die die nötigen Daten liefern könnten, um die Kontroverse zu entscheiden.
Schätzungen zur Größenordnung der Meeresspiegelregression sind ebenfalls unterschiedlich und variieren zwischen 50 und 100 Metern. Diese Schätzwerte weisen der Obertrias den geringsten Meeresspiegelrückgang aller größeren geologischen Aussterbeereignisse zu (abgesehen vom devonischen Ereignis, das, wie wir uns erinnern, mit einem Anstieg des Meeresspiegels verbunden war). Was die Ursachen dieses absinkenden Meeresspiegels betrifft, ist die kontinentale Vereisung der Nordhalbkugel infolge der Wanderung Nordpangäas über die Nordpolarregion ein offensichtlicher Kandidat, obwohl eindeutige obertriassische Glazialsedimente – Tillit – und/oder Dropstone-Ablagerungen rar sind. Es könnte auch sein, dass die starke Ausbreitung tektonischer Subduktionszentren innerhalb dieses Zeitabschnitts zur Instabilität des |139|Meeresspiegels beigetragen hat, indem sie zur Entstehung neuer Ozeanbecken führte und die Wärmeflussmuster an den mittelozeanischen Rücken veränderte.
Obernorische Sedimente in Nova Scotia sind ausgiebig nach geochemischen und physikalischen Beweisen für einen Himmelskörperimpakt an der Trias-Jura-Grenze (TJ-Grenze) abgesucht worden. Bis heute sind die Ergebnisse dieser Suche negativ. In Zentralportugal ist ein großes (35 Kilometer im Durchmesser) rundes topografisches Merkmal gefunden worden, das zuerst auf Satellitenbildern entdeckt wurde: der Einschlagkrater im Distrikt Guarda. Die Entstehung dieser Struktur wurde auf vor etwa 200 Millionen Jahren datiert. Sollten zukünftige Analysen diese Schätzung als zutreffend erweisen, würde die Formation dieser Struktur zeitlich mit der TJ-Grenze zusammenfallen. Basierend auf ihrem Alter und der kreisförmigen Gestalt wird diese Struktur in manchen Zusammenfassungen als (möglicher) Impaktkrater aufgeführt. Allerdings wurden bis heute keine zweifelsfreien Beweise für diese Interpretation vorgelegt. Die meisten veröffentlichten Übersichten und/oder Berichte über das obertriassische Aussterbeereignis lehnen stattdessen die Vorstellung ab, dass das Massensterben in irgendeiner Weise mit einem außerirdischen Impakt zusammenhängen könnte.
Anders als die Verbindung mit Anzeichen eines Himmelskörpereinschlags konnte der Zusammenhang dieses Aussterbeereignisses mit der gewaltigen Eruption einer magmatischen Provinz gut nachgewiesen werden. Entlang einer Linie, die sich quer über die gesamte (moderne) östliche Meeresküste der USA zog – im Westen bis nach Texas und im Süden möglicherweise bis nach Venezuela –, ereigneten sich während der Obertrias außergewöhnlich große Eruptionen magmatischer Provinzen. Die Basaltlavagruppen, aus denen die Provinz besteht, besitzen Gegenstücke des exakt gleichen Alters in Nordeuropa und Westafrika. Diese Eruptionen stellen die ersten Schritte der tektonischen Grabenbildung in Pangäa dar, allgemein auch bekannt als Zentralatlantische Magmatische Provinz (Central Atlantic Magmatic Province, CAMP) (siehe S. 140 und Tanner 2004).
In ihrer Gesamtheit bedecken die CAMP-Vulkangesteine eine Region von mindestens zehn Millionen Quadratkilometern mit einem Volumen von zwei bis drei Millionen Kubikkilometern. Dieses Material wurde infolge einer Reihe separater Eruptionen eingebettet, die episodisch in der gesamten Grabenzone auftraten. Die Gesamtdauer der eruptiven Aktivität der CAMP wird auf etwa 500.000 Jahre geschätzt. Einzelne |140|Eruptionszentren waren bis zu 100.000 Jahre lang aktiv, wobei die meisten in einer deutlich kürzeren Zeitspanne ausbrachen.
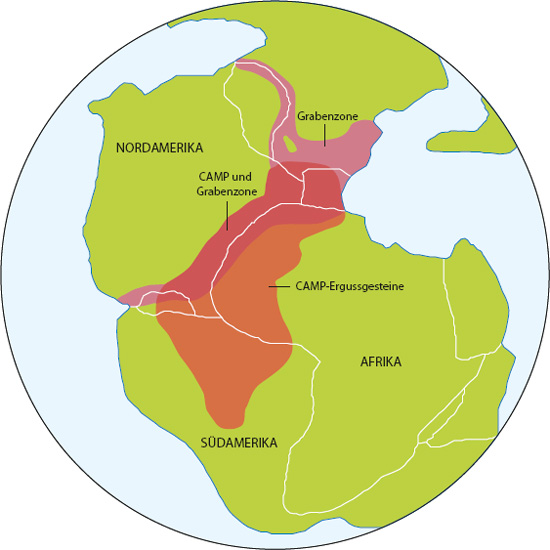
Position der Zentralatlantischen Magmatischen Provinz (Central Atlantic Magmatic Province, CAMP) in einer Rekonstruktion der obertriassischen Paläogeografe.
Was den Zeitpunkt angeht, fand dieses Ereignis mit einer möglichen Abweichung von ± zwei Millionen Jahren vor etwa 200 Millionen Jahren statt. [Anmerkung: Der genannte Fehlerschätzwert bezieht sich auf das radioisotopische Datum von 200 Millionen Jahren, nicht auf die Dauer des gesamten CAMP-Einbettungsereignisses.] Diese Datierung verortet den CAMP-Vulkanismus eher im Rhaetium als im Norium. Sepkoskis Familien- und Gattungskompendium schloss das Rhaetium nicht als triassische Stufe ein, sondern listete stattdessen das Norium als oberste triassische Stufe. Unter der vertretbaren Annahme, dass Sepkoski in seiner Analyse die Stufen des Noriums und Rhaetiums zusammenfasste, bleibt |141|die Verbindung der CAMP-Daten und der Datierung des Höhepunktes des norischen Aussterbens demnach haltbar.
Unter Berücksichtigung der Unklarheiten hinsichtlich der Korrelation von paläontologischen und geologischen Daten innerhalb dieses Zeitintervalls ist das Aussterbeszenario, das mit der Mehrheit der Daten übereinstimmt, ein längerfristiges Muster im Fossilienbestand, das wahrscheinlich das Ergebnis des Meeresspiegelrückgangs in der späten Trias ist. Wie wir bei früheren Massensterben gesehen haben, hätte der Rückgang des Meeresspiegels den Umfang der warmen Flachmeere reduziert, die einen Großteil der triassischen kontinentalen Plattformen bedeckten. Eine Verringerung des Lebensraums hätte Meeresorganismen, die an neue Orte abwandern konnten, in einen intensiven Wettbewerb miteinander gestellt. Sesshafte Arten – einschließlich der Riffe – wären natürlich einfach an Ort und Stelle gestorben, wenn die Umgebungsbedingungen die artspezifischen Toleranzen überschritten hätten. An Land wirkten die gleichen Prozesse in umgekehrter Richtung. Die sich zurückziehenden Küstenlinien hätten Habitate in Richtung der Ozeanbecken verlagert und mobile Arten dazu gezwungen, mit ihnen zu wandern. Aber im Gegensatz zu marinen Lebensgemeinschaften, die sich darauf verlassen konnten, dass das Ozeanwasser über weite Gebiete eine konstante Umwelt aufrechterhält, hätten die trockengelegten Kontinente an den Breitengraden erheblich steilere Temperaturunterschiede und aufgrund der Kontinentalität zunehmend extreme Umweltbedingungen erlebt.
Dieser Mechanismus passt zu den langfristigen Mustern, die wir in den Ozeanen beobachten können. Die Details der terrestrischen triassischen Fossilien sind weniger gut erschlossen, insbesondere in Bezug auf die Zeitpunkte der wichtigsten biotischen Veränderungen. Aber nichts scheint im Widerspruch zu dieser allgemeinen Erklärung für das Aussterben in der späten Trias zu stehen, vor allem wenn die Wettbewerbsmuster mit den neuen, durch Evolution entstandenen, erfolgreichen Adaptationen berücksichtigt werden (z.B. die Verdrängung von Lystrosauriern durch Thecodonten). Ganz am Ende der Trias setzte der CAMP-Vulkanismus die gesamte Biosphäre zusätzlich unter Stress, was infolge der erhöhten planetaren Albedo und der Veränderungen im Niveau und Muster der Erosion und Nährstoffzufuhr durch die in die Atmosphäre entlassenen Treibhausgase die globalen Abkühlungsmuster verstärkte.