Historical Biogeography
Michael J. Donoghue
OUTLINE
1. Early developments
2. Cladistic biogeography
3. Inferring ancestral areas
4. A fresh look at old patterns
5. Beyond the standoff
Historical biogeographers try to understand how life and the earth have evolved together, accounting for current geographic distribution patterns in terms of past events. They try to understand where lineages originated, and how, when, and why they have spread, adapted to new environments, and diversified. These spatially oriented questions are as central to evolutionary biology today as they were at the time of Darwin and Wallace. Although we are still analyzing patterns that were noted long ago, the landscape of ideas and methods has changed dramatically over the years. A key recent period, beginning in the 1970s, saw the rise of cladistic biogeography in its various forms. Although this period of conceptual and methodological turmoil served to clarify fundamental issues, in hindsight it appears that an overly narrow view of the permissible questions and admissible evidence resulted in relatively little progress in understanding empirical patterns. Today, the emphasis has shifted to more integrative approaches, especially in regard to inferring ancestral areas using methods capable of accommodating information on the ages of lineage-splitting events and on the relative likelihood of geographic movements at different times in the past. Consequently, we are now taking a fresh look at a number of long-recognized biogeographic patterns. The major challenge ahead is to fill the wide gap still separating those focused on general patterns and the relationships among areas of endemism from those developing and using methods to infer ancestral areas within particular lineages.
GLOSSARY
Ancestral Area/Center of Origin. Geographic area where a clade (or multiple clades) originated and began to diversify before spreading to other areas; this may or may not be a center of diversity.
Area of Endemism. A geographic area that harbors multiple endemics.
Clade. A monophyletic group. An entire branch of a phylogenetic tree, including an ancestor (e.g., an ancestral species) and all its descendants.
Cladogram. A branching diagram that depicts hypothesized relationships among the terminals. Includes area cladogram, with geographic areas inhabited by terminal taxa replacing the names of those taxa (often species; figure 1), and general area cladogram, a summary tree for a set of area cladograms for different groups of organisms occupying the same areas of endemism. The term phylogeny (or phylogenetic tree) is commonly used when the intention is to convey information about the inferred evolutionary history of a group of organisms.
Dispersal. The movement of organisms resulting in the expansion of the geographic range of a species; long-distance dispersal generally refers to movement well outside the current range across a natural dispersal barrier (a mountain range, ocean, etc.).
Endemic. A species or clade that is naturally restricted in its distribution to a particular geographic area.
Integrative Biogeography. The attempt to incorporate multiple sources of relevant evidence in biogeographic inferences; moving beyond area cladograms, this might include information on the timing of lineage-splitting events and on the likelihood of movements between areas at different times in the past.
Niche Conservatism. The tendency for related species (possibly even large clades) to retain ancestral ecological characteristics. In historical biogeography, this is manifested in the phenomenon of habitat tracking, where lineages spread and contract within retained environments; niche conservatism ultimately underlies disjunct distributions.
Pseudocongruence. When area cladograms for different groups of organisms are the same even though the groups diversified at different times and in response to different causal events (figure 1).
Vicariance/Cladistic Biogeography. An approach to the identification and analysis of biogeographic patterns in which area cladograms are the underlying source of evidence and general area cladograms provide the basis for identifying common causes (often vicariance events related to earth history).
Vicariance/Vicariance Event. The splitting of an ancestral geographic range (of a species or clade) or of an entire biota by the formation of a barrier, often geological in nature, such as drifting continents, mountain building, or climate change.
Historical biogeography, to paraphrase Leon Croizat (1964), is the study of how life and the earth have evolved together. The fundamental aim is to account for the current geographic distributions of species and clades in historical terms. From this standpoint it is important to infer where lineages originated, how, when, and why they spread to other areas, and how such movements influenced genetic variation, adaptive evolution, speciation, and extinction. This problem can just as well be viewed from the standpoint of particular biotas, asking how these were assembled through time and, therefore, how long the component lineages have been interacting. And from yet another angle, studies of patterns of endemism and disjunction can help us understand the history of the earth and its changing climates.
Historical biogeography forms a natural bridge between evolutionary biology and the earth sciences, including paleontology and climatology. It also connects directly to the study of ecological processes (perhaps especially to life history and reproductive biology), and to genetic studies of population histories within species (phylogeography; see chapter II.5). It is clear that historical biogeography is, and must always be, very broadly integrative, and this necessity has presented challenges for the development of a unified methodology (Morrone 2009; Lomolino et al. 2010). There has been a tendency to default to a narrative mode of explanation, as opposed to a hypothesis-testing mode, and it has proven difficult to move beyond individual case studies to draw very general conclusions (Crisp et al. 2011). Not surprisingly, the history of historical biogeography has been marked at intervals by heated debate over proper methodology and relevant evidence.
1. EARLY DEVELOPMENTS
Historical biogeography has deep pre-evolutionary roots (Lomolino et al. 2004). The recognition of many biogeographic patterns and the origin of several general conceptual approaches go back to the eighteenth century. For example, Georges-Louis Buffon (1707–1788) made the fundamental observation (now known as Buffon’s law) that widely separated but environmentally similar regions are typically inhabited by quite different assemblages of species. Likewise, the recognition that floristic belts and species diversity tend to shift in parallel along latitudinal and altitudinal gradients traces to Alexander von Humboldt (1769–1859).
From the beginnings of evolutionary thought in the early to mid-nineteenth century, geographic patterns figured prominently both as challenges to evolutionary explanation and by providing support for descent with modification. The ideas of Charles Darwin (1809–1882) and Alfred Russel Wallace (1923–1913) were to a significant extent inspired by their direct experiences with geographic patterns in nature (for Darwin, think Galápagos; for Wallace, the Malay Archipelago). Both used biogeographic patterns as key evidence of shared ancestry, but also recognized the need to explain oddball disjunctions that might otherwise be seen as favoring separate creation. Darwin devoted two chapters of The Origin to biogeography, and these were largely designed to defend the continuity of evolution: “so in space, it certainly is the general rule that the area inhabited by a single species, or by a group of species, is continuous, and the exceptions, which are not rare, may, as I have attempted to show, be accounted for by former migrations under different circumstances, or through occasional means of transport, or by the species having become extinct in the intermediate tracts.”
As but one example, Darwin and Wallace both devoted considerable energy to explaining (invoking two different mechanisms) bipolar distributions (lineages distributed at both high northern and high southern latitudes, but absent in between), which are now perfectly understandable in evolutionary terms. Under the circumstances at the time, it is hardly surprising that they focused special attention on the likelihood of movements (including chance long-distance dispersal over existing geographic barriers) over long stretches of geological time.
Wallace, more so than Darwin, devoted his energies in the late 1800s to historical biogeography, and is frequently identified as “the father of zoogeography.” He wrote three major books on the subject—The Malay Archipelago (1869), The Geographical Distribution of Animals (1876), and Island Life (1880)—and famously identified the biogeographic discontinuity between the Australian and Oriental faunas now known as Wallace’s line. Other key figures at that time included the British botanist Joseph Dalton Hooker (1817–1911), who, in contrast to Wallace and Darwin, favored the rise and fall of land bridges (“extensionism”) as an explanation for major intercontinental disjunctions, and the American botanist Asa Gray (1810–1888), who focused special attention on disjunctions in temperate forests around the Northern Hemisphere. The British ornithologist Phillip Sclater (1829–1913) first circumscribed the world’s major terrestrial biogeographic regions, and Edward Forbes (1815–1854) likewise identified marine biogeographic realms.
In the first half of the twentieth century, William Diller Matthew (1871–1930) emphasized “centers of origin,” proposing that primitive forms would occupy more inaccessible areas. John Willis (1868–1958) formulated the “age and area” hypothesis, whereby the geographic range of a taxon depended on its age. Stanley A. Cain (1902–1995) provided a badly needed critique of the center of origin concept, and the criteria by which these “centers” were recognized in practice.
George Gaylord Simpson (1902–1984) and Phillip Darlington (1904–1983) were prominent among zoological biogeographers in the Wallace-Darwin-Matthew tradition. They drew special attention to migration routes of different types (“corridors,” “sweepstakes routes”), envisioning movements over a more or less fixed landscape. At first they dismissed continental drift as a factor, although a dynamic earth clearly provided a completely new interpretation for patterns at the level of whole biotas. Later they reconsidered this stance, though Darlington, in particular, cautioned that drifting continents might be too ancient to be relevant to many modern geographic patterns.
Beginning with E. V. Wulff (1885–1941), phytogeographers connected plant disjunction patterns to past continental movements, and this oriented Peter Raven and Daniel Axelrod’s masterful synthesis in 1974. In the meantime, led by Sherwin Carlquist, botanists continued to focus on dispersal biology and the colonization of oceanic islands. A significant development during this period was Leon Croizat’s “panbiogeography” (see Croizat 1964). Importantly, Croizat emphasized the search for general patterns (“generalized tracks”), vicariance explanations (“baselines”), and the merging of biotas (“nodes”). To this day Croizat’s approach has attracted a small but vocal following, though many fault his approach for shunning phylogenetic information.
2. CLADISTIC BIOGEOGRAPHY
With the rise of phylogenetic systematics came the use of phylogenetic trees in historical biogeography. Willi Hennig (1966) featured the placement of trees on maps, and the interpretation in terms of lineage movements (e.g., the “progression rule” to identify a center of origin). Another dipterist and contemporary, Lars Brundin, had the more profound influence through his empirical work on the chironomid midges of the Southern Hemisphere. Brundin compared multiple phylogenies of midge lineages overlaid on a map of the southern continents, seeking general patterns caused, perhaps, by drifting continents. Brundin’s work, combined with Croizat’s outlook, spawned the development by Gareth Nelson and colleagues of what was initially known as vicariance biogeography or, later, cladistic biogeography (e.g., Platnick and Nelson 1978). This reflected a strong negative reaction to the one-off dispersal scenarios for individual groups that had come to dominate the biogeographic literature. Instead, the focus was squarely on vicariance-causing events (especially continental movements) that could impact multiple lineages, and on inferring relationships among areas of endemism as a way to elucidate earth history.
Methodologically, vicariance biogeography focused almost exclusively on the comparison of cladograms for different groups of organisms, but with the names of the terminal taxa replaced by the areas of endemism they occupy (so-called area cladograms). The goal was to derive a general area cladogram that best summarized the geographic relations among the areas in an underlying set of area cladograms (e.g., Wiley 1988). This was easy in the simple three-area cases used repeatedly to illustrate the basic approach, but proved far more difficult in practice, especially when species were widespread (found in more than one area), when an area appeared in more than one place in an underlying area cladogram, when particular areas were missing from any of the area cladograms, or with a combination of these factors. One set of methods focused on tallying shared geographic “components,” employing various assumptions about widespread taxa, etc. In recent years, this general approach has spawned several others, including “three-area analysis” and “paralogy-free subtree analysis,” which variously focus on subsets of the underlying area cladograms. Another popular method, known as Brook’s parsimony analysis (BPA), decomposed each area cladogram into a set of binary characters representing the area connections, and compiled these into a matrix that was analyzed by parsimony to derive a single tree showing area relationships.
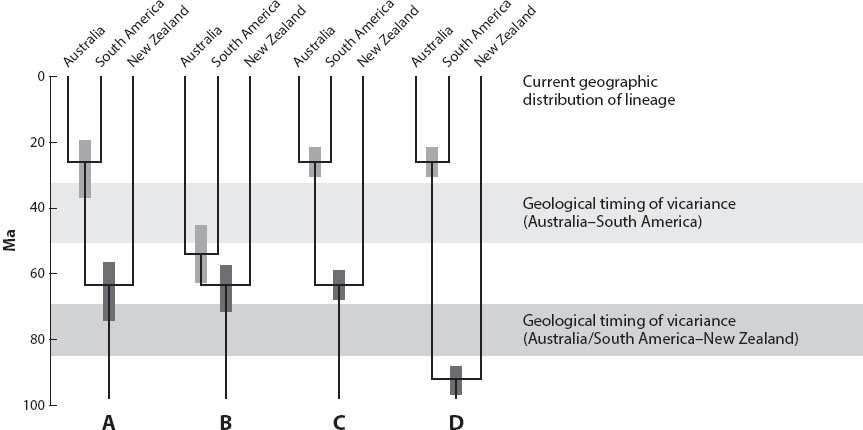
Much of the emphasis in the cladistic biogeography literature was on philosophical and methodological issues, which were reviewed repeatedly, but with few breakthroughs. Real-world cases were typically complicated enough that area cladograms alone lacked the power to confidently resolve area relationships. Among other problems, cases of pseudocongruence came to light, in which events that took place at different times in the past yielded the same area cladogram (Donoghue and Moore 2003; figure 1). By the mid-1990s even the leaders of the vicariance movement had become frustrated by the lack of progress. Unfortunately, however, positions hardened. For example, although information on the timing of lineage-splitting events has been widely viewed as potentially useful in sorting out competing biogeographic explanations (e.g., these might be far too old or too young in relation to a particular earth history event; figure 1), such information has been dismissed by some cladistic biogeographers as being too questionable or distracting from the mission of identifying general patterns in area cladograms.
Figure 1. Four clades showing the same area cladogram: New Zealand (Australia + South Africa). The inferred ages for the divergences in Clades A and B (confidence limits indicated by bars at the nodes) are consistent with vicariance caused first by the separation of New Zealand from Australia + South America, and then by the separation of Australia and South America. In contrast, the inferred divergence times in clades C and D are inconsistent with these geological causes; these clades illustrate instances of pseudocongruence. (Modified from Trends in Ecology and Evolution, 26/2, Michael D. Crisp, Steven A. Trewick, and Lyn G. Cook, "Hypothesis Testing in Biogeography," p. 69 [2011], with permission from Elsevier.)
3. INFERRING ANCESTRAL AREAS
One of the hallmarks of the development of cladistic biogeography was the dismissal of the problem of inferring areas of origin for individual lineages or centers of origin for entire biotas. Such pursuits had been central in the history of biogeography, but were denigrated by cladistic biogeographers as fundamentally unscientific. Given such strong objections, it came as a pleasant surprise when the issue resurfaced in the 1990s, now in a phylogenetic framework. This development began with an inference that the common ancestor of the plant clade Asteraceae had probably lived in South America and, following a healthy debate, culminated in the development of a new optimization procedure designed specifically for biogeographic problems (Ronquist 1997). This procedure entailed the key realization that reconstructing ancestral geographic areas was not the same as reconstructing ancestral character states. Area reconstruction needed to place values (or costs) on the events that specifically mattered to historical biogeography, namely speciation, extinction, dispersal, and vicariance. The resulting “event-based” approach, implemented in the software package DIVA, was easily applied to real-world data sets and quickly became the method of choice for biogeographic studies focused on individual clades.
As this discussion proceeded, it was recognized that the problem of reconstructing the geographic evolution of clades was parallel in many respects to the problem of deciphering gene trees within species trees, or of parasites on their hosts (e.g., Page and Charleston 1998). All three of these problems can be conceived as involving trees of different sorts that track one another with greater or lesser fidelity (e.g., in historical biogeography, trees of geographic areas and trees of the taxa in those areas), and it was natural to identify parallel processes in the different systems. For example, vicariant speciation could be equated with codiversification in the case of gene and species trees, speciation within an area to gene duplication, extinction to lineage sorting, and dispersal to lateral gene transfer. The hope was that methods developed in one area could be ported to problems in the other areas, and that a general theory would emerge; however, despite the similarities, there are also disanalogies. For example, in biogeography the repeated movement, at different times, often among a limited set of areas via the same corridor (e.g., iterated movement of lineages through the Bering Land Bridge), might yield many more instances of pseudocongruence than in the other systems. Nevertheless, it is noteworthy that maximum likelihood and Bayesian approaches began to be applied to the gene-tree and host-parasite cases, primarily to evaluate more complex models specifically including time as a variable.
The development of maximum likelihood methods for the inference of ancestral areas got under way with Ree et al. (2005), who developed what was later dubbed the dispersal-extinction-cladogenesis (DEC) model. The parsimony-based approach in DIVA was unable to take into account the timing of splitting events in the underlying lineage tree, or changes in the likelihood of movements among areas through time (e.g., the existence of a corridor during some period but not in others, or the increasing distance between two continents). The underlying model of diversification (how daughter species inherit geographic ranges) is another important concern, and the method developed by Ree et al. (2005) specifically provided for maintenance of a widespread species through a speciation event within a subregion. Initially, likelihood calculations were made using simulations, but an analytical solution (calculating probabilities of dispersal and extinction as functions of time using a rate matrix) was soon implemented in the software package Lagrange. Such approaches are open ended, and recent efforts have incorporated additional relevant information such as the physical sizes of geographic regions and the distances that separate them.
4. A FRESH LOOK AT OLD PATTERNS
Over the past decade, considerable progress has been made in understanding several classical biogeographic patterns. To give the flavor of such findings, I will briefly highlight below just three intercontinental disjunction patterns; similar advances have been made within several continents and biogeographic regions (e.g., the Mediterranean, Australia, and southern Africa), and in some island systems (e.g., Hawaii and the Canary Islands). In general, new dating information—both on lineages and on geological and climatic events—is having an important impact on our interpretations, and concerted movements and long-distance dispersal are once again being considered as possible causes alongside continental movements (de Queiroz 2005).
Aside from insights into intercontinental disjunction patterns, historical biogeography has increasingly entered discussions of global biodiversity patterns and conservation. A good example is provided by the latitudinal species-richness gradient. Many ecological explanations have been put forward for this gradient, but it is possible that it is largely explained by the initial diversification of many lineages in the tropical climates that were widespread in the Paleocene and Eocene, followed by more recent movements into temperate, boreal, and arid biomes with global cooling since the Eocene (e.g., Wiens and Donoghue 2004). A similar explanation may also hold for the equivalent latitudinal gradient seen in marine organisms.
Laurasian connections. Disjunctions have long been evident between eastern Asia and eastern North America, and these are embedded in a broader Laurasian distribution pattern that also involves endemics in Europe and western North America. Dated phylogenies imply that there have been multiple movements of lineages around the Northern Hemisphere, and that many of these were movements through Beringia (as opposed to the North Atlantic Land Bridge) at times when climates were accommodating. It appears that Asia has been a source area for a number of lineages, but movement has occurred in both directions. Fossil evidence indicates that a number of lineages now confined to Asia were more widespread around the Northern Hemisphere in the Eocene and Oligocene, and that ranges have become restricted with the cooling and/or drying of climates in some regions (Europe and western North America, in particular). Collectively, these patterns provide evidence of niche conservatism. Many Northern Hemisphere plant lineages, for example, diversified solely within temperate forests throughout the Cenozoic, though a few radiated into drier regions as these have spread since the Miocene in western North America and around the Mediterranean basin.
Gondwanan connections. Gondwanan disjunction patterns have long attracted attention, with an emphasis on the role of past continental movements. Although there are disjunctions that may well have been caused by the drifting of the southern continents, new information on divergence times cautions against the universal application of such an explanation. Indeed, many Africa/South America plant disjunctions that were once thought to be the result of the breakup of Gondwanaland, are being reinterpreted in light of evidence that their divergences are far younger than 100 million years ago. Disjunctions dated to the Eocene are now often interpreted as reflecting movements through the Northern Hemisphere when climates were warmer and connections across southern Laurasia more continuous. Other recent studies have featured “west-wind drift” as an explanation for significant connections between Australia and New Zealand; these may have been established through repeated dispersal with the prevailing winds after the breakup of the southern continents. Finally, contrary to earlier theories emphasizing the southward migration of groups that originated in the north, a number of studies have argued for a Southern Hemisphere origin of some major, now-cosmopolitan, groups, including birds and the sunflowers and their relatives (Asteraceae).
New World connections. It has long been appreciated that South America existed in relative geographic isolation through much of the Cenozoic, and that regular north-south connections were established only more recently (especially with the formation of the Isthmus of Panama), setting off “the great American interchange.” Recent studies have reported a wider than expected range of dates for disjunctions between North and South America, with plant lineages in particular showing a number of older splits, perhaps reflecting more frequent long-distance dispersal than in terrestrial vertebrates. At the same time, however, new geological evidence is suggesting the possibility of earlier corridors between the two landmasses (e.g., in the Eocene-Oligocene some 35–33 Ma), and that the docking of the continents may have taken place considerably earlier than the previously accepted time frame of about 3 million years ago. Regarding directionalities, recent studies have largely substantiated the view that more tropical plant clades moved north from South America, while more high-elevation clades moved from north to south. A number of studies have documented the movement of alpine elements into the Andes from the north, accompanied by greatly elevated rates of diversification. In mammals, where it has been possible to integrate an extensive fossil record, it appears that more lineages moved from North America to South America than in the opposite direction, and the northern elements tended to radiate more extensively in the south (e.g., deer, canids, sigmodontine rodents) than did the southern elements in the north (e.g., sloths, agoutis, and opossums).
5. BEYOND THE STANDOFF
Over the last several decades a counterproductive chasm opened between two broad camps focused on different aspects of historical biogeography. One group has attempted, primarily using area cladograms as evidence, to discern general patterns in disjunctions across multiple lineages, with the aim of assessing past connections among areas of endemism. Another group has attempted to infer ancestral areas and pathways of movement within particular lineages using phylogenetic trees and methods that incorporate varying amounts of other relevant information, such as the absolute timing of splitting events. Both these objectives are not only perfectly legitimate, but also, within limits imposed by the data, attainable. Most importantly, they are complementary and jointly necessary to fully understand historical biogeography. If nothing else, they could directly benefit one another. On the one hand, the search for general patterns would benefit from information on the timing of events. For example, this information would help weed out instances of pseudocongruence that plague the analysis of area cladograms. On the other hand, the inference of past biogeographic movements within particular lineages would benefit greatly from knowledge of patterns and processes in other groups of organisms. Methodologically, it should be possible to bring all this information together in a likelihood or Bayesian framework, and we might even realize the long-imagined feedbacks between biogeography and the earth sciences. For example, information on the history of areas could be incorporated into estimations of divergence times within clades, or biogeographic information could be used in choosing among competing geological models.
Fortunately, a new generation of historical biogeographers has the opportunity for a fresh start, identifying and synthesizing the best ideas and methods that have been developed. The resulting integrative historical biogeography will yield a far richer understanding of the spatial history of lineages and of the earth itself, and will add even greater value to the study of ecology, evolutionary biology, and biodiversity conservation.
FURTHER READING
Crisp, M. D., S. A. Treweck, and L. G. Cook. 2011. Hypothesis testing in biogeography. Trends in Ecology & Evolution 211: 66–72. A recent critique of the field, with an emphasis on framing testable hypotheses.
Croizat, L. 1964. Space, Time, Form: The Biological Synthesis. Published by the author. Caracas, Venezuela. One of several prodigious self-published volumes, this provides an insight into the iconoclastic views of Croizat, the founder of “panbiogeography.”
de Queiroz, A. 2005. The resurrection of oceanic dispersal in historical biogeography. Trends in Ecology & Evolution 20: 68–73. An argument that the importance of long-distance dispersal across oceans was underestimated in the era of vicariance biogeography.
Donoghue, M. J., and B. R. Moore. 2003. Toward an integrative historical biogeography. Integrative and Comparative Biology 43: 261–270. An argument for integrating the timing of lineage-splitting events into biogeographic analyses, especially to identify instances of pseudocongruence.
Hennig, W. 1966. Phylogenetic Systematics. Urbana: University of Illinois Press. The founding document of phylogenetic biology, which includes examples of the use of phylogenetic trees in relation to biogeography.
Lomolino, M. V., B. R. Riddle, R. J. Whittaker, and J. H. Brown. 2010. Biogeography. 4th ed. Sunderland, MA: Sinauer. A textbook treatment of the entire discipline of biogeography.
Lomolino, M. V., D. F. Sax, and J. S. Brown. 2004. Foundations of Biogeography. Classic Papers and Commentaries. Chicago: University of Chicago Press. A compilation and analysis of key historical writings on biogeography; an excellent starting point for understanding the history of the discipline.
Morrone, J. J. 2009. Evolutionary Biogeography: An Integrative Approach with Case Studies. New York: Columbia University Press. A recent overview of the wide variety of methods in use in historical biogeography.
Page, R.D.M., and M. A. Charleston. 1998. Trees within trees: Phylogeny and historical associations. Trends in Ecology & Evolution 13: 356–359. A comparison of historical biogeography with the gene-tree/species-tree problem and the host-parasite codiversification problem.
Platnick, N. I., and G. Nelson. 1978. A method of analysis for historical biogeography. Systematic Zoology 27: 1–16. A key early paper on the vicariance/cladistic biogeographic approach.
Ree, R. H., B. R. Moore, C. O. Webb, and M. J. Donoghue. 2005. A likelihood framework for inferring the evolution of geographic range on phylogenetic trees. Evolution 59: 2299–2311. The introduction of a maximum likelihood method for historical biogeography, taking into account the timing of lineage-splitting events and changes in the likelihood of geographic movements through time.
Ronquist, F. 1997. Dispersal-vicariance analysis: A new approach to the quantification of historical biogeography. Systematic Biology 46: 195–203. The introduction of a parsimony-based approach to the inference of ancestral geographic ranges that minimizes the number of implied dispersal and extinction events.
Wiens, J. J., and M. J. Donoghue. 2004. Historical biogeography, ecology, and species richness. Trends in Ecology & Evolution 19: 639–644. An argument for the reintegration of historical biogeography and ecology in explaining global distribution patterns such as the latitudinal species-richness gradient.
Wiley, E. O. 1988. Vicariance biogeography. Annual Review of Ecology and Systematics 19: 513–542. An overview of the concepts and methods of vicariance biogeography, after the first decade, with simple explanations of component analysis and Brooks parsimony analysis.