The Fossil Record
Noel A. Heim and Dana H. Geary
OUTLINE
1. Fossilization and taphonomy
2. The nature of the fossil record
3. Marine diversity in the Phanerozoic
4. The value of the fossil record
The fossil record documents the history of life over the course of the past 3.5 billion years, demonstrates that evolution has occurred, and provides otherwise inaccessible insights into the evolutionary process. This chapter outlines briefly how the fossil record has been formed, and explores the nature of the fossil record in relation to its central role in understanding evolution. Evolutionary biology is a historical science, and the process of evolution is often played out over intervals of time much too long for direct observation. Thus the fossil record provides the dimension of time that is essential for a complete understanding of the process that unites all of biology.
GLOSSARY
Body Fossil. The fossilized remains of a once-living organism. Body fossils represent the actual organism and are distinct from trace fossils.
Stratigraphy. The study of how and why rocks are deposited in their observed vertical and lateral successions. A key stratigraphic concept is that of superposition, where vertical successions of rock are ordered with the oldest strata (layers) at the bottom and the youngest at the top. Although younger rocks are always deposited on top of older rocks, primary “layer cake” stratigraphic successions can be altered through tectonic folding and faulting.
Taphonomy. The study of the ways in which the dead remains of once-living organisms become preserved as fossils. Much of taphonomy is concerned with understanding which biological information is preserved by fossils and which is lost.
Time Averaging. The mixing of noncontemporaneous individuals into a single sedimentary or fossil assemblage.
Trace Fossil. The record of behavior preserved in the sedimentary record. Trace fossils include footprints, burrows, feeding traces, coprolites, and insect leaf damage. Trace fossils are frequently not attributable to a specific species.
Unconformity. A surface separating two stratigraphic units that represents “missing” geologic time. Unconformities are formed through the erosion of previously deposited sediments, prolonged intervals of nondeposition, or a combination of the two.
Uniformitarianism. The idea that the processes that are observable and operating today, and only those processes, can be used to explain the geological and biological evolution of the earth as preserved in the geological and fossil records.
The value of the fossil record is perhaps most clearly illuminated if we try to imagine our knowledge of evolution without it. Plenty of evidence would indicate that evolution had occurred, but the rich history of life would simply be a matter for conjecture. Fossils reveal our history in amazing and often-unpredictable ways: giant ground sloths and saber-toothed cats roaming the landscapes of our present-day cities just a few millennia ago, diverse genera of camels, horses, and rhinos not long before that, and earlier still, flightless birds that preyed on tiny horses and flying reptiles the size of airplanes, to name just a few. We would know that whales are mammals, and we could use molecular evidence to determine their closest living relatives, but how much richer the story becomes when we can actually find their ancestors on the ancient shores of a warm tropical ocean and touch the diminutive leg bones of a 15 m Basilosaurus. We would likely suppose that great calamities had occurred, but who could imagine a global deep freeze complete with tropical glaciers, followed by the most remarkable blossoming of life in history?
The fossil record, however, is not only a catalog of wonderful organisms. It also demonstrates without a doubt that evolution happened. It provides us with transitional organisms between major groups that demonstrate clearly how one evolved from the other (e.g., land vertebrates from fishes, birds from dinosaurs, mammals from reptiles, and many more). The fossil record gives us the ability to determine the actual time frame of evolutionary change, and importantly, it continues to provide valuable information about how evolution occurs.
Evolution can be slow on human timescales. Thus the sequence of evolutionary events preserved in the fossil record is essential for testing the validity of existing evolutionary theory and modifying or expanding our set of ideas on how the process works. For example, the fossil record demonstrates that rates of evolution need be neither slow nor constant, even within lineages. Paleontologists have also shown that evolutionary history has been marked by various types of extinction events (e.g., mass extinctions, mass depletions, turnover events; see chapter VI.13). The field of macroevolution, the study of evolution at or above the species level, is deeply grounded in the fossil record (see chapters VI.12 and VI.13).
1. FOSSILIZATION AND TAPHONOMY
Fossils are the remains or traces of ancient organisms preserved in the rock record. Fossils are found in sedimentary rocks, including sandstones, siltstones, and limestones—rocks that have formed by the accumulation of particles of sediment and/or the skeletal remains of organisms. Sedimentary rocks form in surficial or near-surface environments, especially aquatic ones such as lake and ocean bottoms, and thus record the occurrence and activities of living organisms for the past 3.5 billion years.
The study of the ways in which freshly dead organisms are incorporated into the fossil record is called taphonomy. Taphonomy explores a broad variety of processes, including decay, postmortem transport, fossilization, time averaging, and postburial alteration. Taphonomic studies can reveal not only how certain organisms become preserved as fossils but also why certain other organisms are not readily fossilizable.
The chance of an individual organism becoming a fossil is vanishingly small, yet museums and outcrops abound with fossil remains. In large part, our abundant fossil record results from the fact that life has such a rich and deep history—countless trillions of organisms have lived and died on our earth. Aside from a good dose of luck, several reasonably predictable factors are key in determining the likelihood of an individual becoming a fossil. The path to becoming a fossil is logically divided into two phases: those processes that happen between death and burial in sediment (collectively termed biostratinomy) and those that occur after burial (collectively termed diagenesis).
Biostratinomic processes are easily visualized because most people have witnessed at least some of them. The death of an organism may be followed by partial or complete consumption by a predator, an interval of scavenging and decay (scavenging on a small scale), mechanical abrasion, chemical dissolution, and bioerosion, as the remains wash back and forth in the surf, roll along in a river, or rest on the ground or the seafloor. Naturally then, a significant fraction of organisms disappear forever in these ways, their atoms eventually recycled in a multitude of other organisms but leaving no remains to fossilize. Biostratinomic processes are not entirely destructive, however; they also control the nature of many fossil deposits through the winnowing, sorting, and concentrating of hard parts by normal sediment transport in water or wind.
It is intuitively obvious that hard parts such as shells, bones, and teeth will withstand postmortem degradation better than soft tissue. It is not surprising, then, that clams have a better fossil record than do slugs, or that more is known about the teeth of ancient mammals than about their livers. Even hard parts have an organic matrix, however, and the ratio of mineral to organic material can be very important in preservation. The cuticle of trilobites, for example, contained a higher proportion of calcium carbonate than does that of a lobster or a crab, which explains why trilobites have a much better fossil record.
Perhaps the single most critical factor in becoming a fossil is having the good fortune to be buried as quickly as possible in sediment of some kind. Rapid burial avoids the ravages imposed by physical and biological processes at the surface. The habitat occupied by an organism has a very important effect on its potential for burial. In general, most terrestrial habitats are areas of net sediment erosion, whereas oceans, lakes, and some river systems are more likely to be areas of net sediment accumulation (although it is geological [tectonic and sea level] factors that govern how much sediment can accumulate, not the habitat per se).
Once the remains of an organism become permanently buried, a variety of biological, chemical, and physical processes may predominate. Enormous spans of time may be involved, of course, and the burial history of an object may be complex. Naturally there are many ways for buried remains to be rendered forever unrecognizable: from complete dissolution in the shallow subsurface to destruction by extreme heat and pressure at depth. Here we focus on those processes that result in fossilization.
Waters that percolate through a deposit play a critical role in determining the fate of the buried remains. These fluids may dissolve skeletal material away or may precipitate minerals, or both. When mineral-laden waters deposit precipitates in preexisting spaces in skeletal material, the result is permineralization. Wood and bone, with their abundant natural pores, are often fossilized in this way, sometimes with fine structural details preserved. The minerals deposited are typically silicates, pyrite, or carbonates. Petrifaction is a closely related process in which organic matter is completely replaced by precipitating minerals. In other cases, both the skeletal and soft tissue components of an organism may be replaced by precipitating minerals (replacement). The dissolution of tissues or hard parts and subsequent precipitation of minerals in their place may happen virtually simultaneously, thereby preserving fine detail, or these events may be widely spaced in time and record only the basic shape of the organism. Fossils may form in more unusual ways as well, including freezing, desiccation, or entrapment in sap (with eventual modification into amber). Under particular conditions (including low oxygen and pH), tissues may be preserved as phosphate, occasionally with spectacular results (e.g., cellular preservation in the Doushantuo Formation [590–565 Ma], arthropod larvae in the Orsten Formation [501–488 Ma]).
The fossilization processes described above, in which various parts of ancient organisms become preserved, generate what are called body fossils. Trace fossils, on the other hand, record the activities of ancient organisms and may include footprints, feeding traces, borings, burrows, and even fossilized feces (coprolites). Trace fossils often provide a wealth of information that would be unavailable from even the best of body fossils, including such important things as feeding behavior, diet, and locomotion.
Taphonomic processes have acted and continue to act as a filter for the preservation of organic remains in the geological record; not all living individuals, species, or higher taxa have an equal chance of preservation. Paleontologists sometimes estimate “paleontological completeness” as the probability of sampling any member of a given taxon within its stratigraphic range (the total amount of geologic time between the oldest- and youngest-known fossils of that taxon). For example, in an interval of about 5 million years, estimates of the probability of finding any genus belonging to a particular group range from 5 percent for polychaetes to 90 percent for brachiopods, trilobites, graptolites, conodonts, and cephalopods (Foote and Sepkoski 1999). Other well-known marine animal taxa fall between, including sponges, corals, crinoids, bivalves, and gastropods, all with probabilities of approximately 40–50 percent.
Fortunately, the taphonomic filter is imperfect and the geological record is scattered with deposits of exquisitely preserved fossils from organisms that would typically be destroyed before fossilization. Lagerstätten is the term generally reserved for fossil deposits with exceptionally well-preserved soft tissues. The importance of these relatively few deposits for understanding the history of life has been profound. Certainly the most well-known lagerstätten is the Middle Cambrian Burgess Shale, discovered in the Canadian Rockies by C. D. Walcott in 1909. The Burgess Shale deserves its fame; it provides a window on an entire community of organisms, including a spectacular variety of soft-bodied forms, from a critical interval near the early expansion of animal life. Of the more than 100 species described from the Burgess Shale, probably fewer than 15 percent have hard parts that would be preserved under more typical conditions of fossilization. Our knowledge of the early history of animal life has been dramatically enhanced in recent years by discovery of other lagerstätten, each containing otherwise-unknown organisms, and many taking the origins of important groups further back in time.
Lagerstätten are not restricted to any particular time interval. The Messel Oil Shale near Frankfurt, Germany, preserves animals and plants from an Eocene lake and its surroundings, including numerous fish, birds, mammals (e.g., primates, bats, pygmy horses, and hedgehogs), and insects (the latter with distinct coloration still present). Other lagerstätten include the Hunsruck Slate (Devonian of Germany), which has revealed much about the nature of trilobite limbs and other soft parts, the Rhynie Chert (Devonian of Scotland) with its early vascular plants, and various Cretaceous localities in China known for their feathered dinosaurs and early flowering plants.
Aside from their exceptional preservation, lagerstätten do not share a particular mode of fossilization. Each deposit differs in the mineralogy of its specimens and in the particular suite of physicochemical conditions that led to their preservation. The general themes that prevail are the following: (1) postmortem exposure to aerobic (oxygenated) conditions is simply not possible: decay processes that occur in the presence of oxygen destroy organic remains very quickly; (2) rapid burial is usually important, although environments with low sedimentation rates may allow for preservation if bottom waters lack oxygen; (3) the chemistry of ambient fluids is key to the mineralization of remains (e.g., pyritization requires low oxygen and high iron concentrations).
Fossil material with well-preserved DNA is not impossible, but it is more rare than one would imagine, given the coverage it receives in the popular press. Most confirmed cases of DNA preservation involve frozen or desiccated organisms less than 1 million years old. Other chemical signatures of life are much more durable, however. Sponge-specific sterane biomarkers from Precambrian sediments of the Arabian Peninsula, for example, indicate that basal metazoans first evolved nearly 100 million years before their earliest known body fossils.
2. THE NATURE OF THE FOSSIL RECORD
Many important questions about evolution can be addressed only by the fossil record. For most of these questions, the issue of temporal resolution is fundamental. How long do individual species persist? How rapidly do they replace one another? (We note that paleontologists use a morphological species concept, which can be supported by data on paleoecology, the geographic and temporal ranges of particular forms, and quantitative methods to objectively differentiate morphological groups.) What is the pace of morphological change within a lineage? How rapid and how simultaneous are mass extinctions? How quickly can communities recover from a major environmental catastrophe? What is the relationship between the timing of a particular group’s first appearance and the timing of major geological events such as continental fragmentation? Before addressing these and other basic questions, a paleontologist must establish the basic temporal parameters of the collections with which he or she is working.
The amount of time represented by a given sample is of critical importance, and a wide spectrum of possibilities exists. A set of fossils found in geographic and stratigraphic proximity may represent a true ecological snapshot; that is, it may include only organisms that lived at the same time and in the same place. The Miocene Ashfall Fossil Beds in Nebraska come close to these criteria; this deposit preserves complete skeletons of multiple species of rhinos, horses, camels, dogs, birds, turtles, and many more organisms that apparently fell victim to the effects of a sudden influx of volcanic ash. All of the animals preserved likely died within a few days or weeks of one another. At the opposite end of the spectrum, a fossil sample may contain individuals that lived at widely separated time intervals. For instance, many modern coastlines include rocky outcrops, often bearing fossils that are Pleistocene or older in age. As these fossils weather out of the nearshore rocks (or unconsolidated sediment), they mingle with the remains of recently dead organisms; the resultant mixture of shells that lived thousands or even millions of years apart may eventually become a fossil assemblage. This mixing of noncontemporaneous material into a single fossil deposit is known as time averaging. Aside from erosion and redeposition (as just described), time averaging may be caused by sediment winnowing or low sedimentation rates, coupled with the accumulation of skeletal material. Once remains are buried, they are still subject to time averaging through bioturbation, the mixing of the upper layers of sediment by burrowing organisms.
Time averaging has been quantified in modern marine settings, primarily on collections of mollusks. Radiocarbon and amino acid racemization dating techniques have shown that most shells are fewer than 3000 years old in modern nearshore settings and fewer than 10,000 years old on the continental shelves.
Although time averaging may seem like a hindrance to understanding ancient ecology and evolution, it has its advantages, too. A fossil collection that contains individuals accumulated over a few thousand years is certainly not an ecological snapshot, but the noise caused by short-term environmental and other fluctuations is reduced. Time averaging in the fossil record permits evolutionary change to be examined without conflating responses to ephemeral or short-period phenomena (e.g., seasonality) with more lasting evolutionary change. For example, two samples of living gastropods collected 100 m apart in the same bay are likely to be more different in species composition and relative abundances than the death assemblages collected at the same locations. This is because localized faunas change rapidly over time through stochasticity in birth, death, recruitment, and locomotion; thus, fewer fossil samples are needed to adequately capture the overall composition of an area when time averaging has occurred.
The fidelity of the fossil record can also be assessed via live-dead comparisons, in which the relative abundance of species in living communities is compared with the relative abundances in nearby accumulations of remains (that, given time, might become a fossil collection). Studies by taphonomists have shown that life assemblages are generally well represented by their corresponding death assemblages in a variety of marine settings (Kidwell and Flessa 1996). These studies have focused primarily on mollusks, which are readily fossilizable and ecologically important, but the results are very likely applicable to a broad spectrum of taxonomic groups. Live-dead comparisons have also demonstrated that most fossil accumulations only very rarely involve long-distance transport of skeletal material, and when they do there is abundant independent evidence that transport has occurred, including size-sorting, fragmentation, imbrication, and/or sedimentological context.
The resolution of a paleontological study is determined by a number of factors: the mode of fossilization, time averaging, and the amount of geologic time missing between fossil collections. Careful examination of modern ecosystems, lagerstätten, and modern sedimentary environments can provide a wealth of information on the first two factors. But the issue of stratigraphic resolution, or missing time, is one of the most commonly criticized aspects of paleontological studies, beginning with Charles Darwin himself.
Darwin famously devoted a chapter in The Origin to the imperfection of the fossil record. This chapter ended with a metaphor:
… The geological record as a history of the world imperfectly kept, and written in a changing dialect; of this history we possess the last volume alone, relating only to two or three countries. Of this volume, only here and there a short chapter has been preserved; and of each page, only here and there a few lines. Each word of the slowly-changing language, more or less different in the successive chapters, may represent the forms of life, which are entombed in our consecutive formations, and which falsely appear to have been abruptly introduced. (Darwin 1872, chapter 10)
Darwin’s main concern with geological completeness, or the lack thereof, was his conviction that natural selection required species to evolve slowly and continuously, leading to an expectation of multiple intermediate forms between ancestral and descendant species. Instead, just as with living species, Darwin found significant morphological gaps between closely related fossil species. The conclusion was inevitable; the fossil record is incomplete.
Darwin was, of course, not a naive geologist, having studied geology at the University of Edinburgh and the University of Cambridge. It was common geological knowledge then as now that no single place records the entirety of earth history. Darwin’s extreme view of geological incompleteness, however, was not entirely based on geological observations but rather colored by the failure of the fossil record to conform to his theory: “But I do not pretend that I should ever have suspected how poor was the record in the best preserved geological sections, had not the absence of innumerable transitional links between the species which lived at the commencement and close of each [geological] formation, pressed so hardly on my theory” (Darwin 1872, chapter 10). Darwin faced a choice when the geological record did not produce the innumerable intermediate forms between species his theory had predicted: reject his theory of evolution by natural selection or attribute the failure of his hypothesis to the poor quality of the fossil record. He chose the latter.
Darwin’s choice was based on strict adherence to the principle of uniformitarianism, which was first proposed by James Hutton in 1788 and fully developed by Charles Lyell in The Principles of Geology. One of the key tenets of uniformitarianism to which both Lyell and Darwin adhered was that rates of natural phenomena have not changed over geologic time—volcanoes erupt with unvarying frequency, lakes fill with sediment at a constant rate, within-lineage speciation rates are invariant, etc. Darwin’s worldview was informed by a strict adherence to constant rates and precluded him from seriously considering the possibility that distinct species appeared rapidly in geologic time.
The geologic timescale was understood in a general way before Darwin wrote The Origin; the Paleozoic, Mesozoic, and Cenozoic eras and nearly all the geological periods were named in the first half of the nineteenth century. Geologists have been working diligently ever since to refine these subdivisions, which form the basis for correlations of rocks of different ages across the globe. The most significant development since Darwin’s day, however, has been the application of radiometric dating to provide numerical ages for the timescale (plate 1). Darwin could only guess at the age of the earth or the amount of time represented by any given sequence of sedimentary rocks (or the gaps contained therein). Our ability to quantify these time intervals now, combined with the wealth of modern paleontological and biological studies of species-level change, has changed our expectations of the way evolution should appear in the fossil record.
We now appreciate that stratigraphically adjacent paleontological samples will commonly be separated in time by tens of thousands of years. It is clear from considerable biological work that such time spans are more than enough time for distinctive new species to be generated. Recognizing these facts, paleontologists no longer have an expectation of geologically gradual sequences: transitions from ancestral to descendant species that play out continuously over hundreds of thousands or even millions of years. Such changes are occasionally preserved, but it is more common that a given stratigraphic sequence provides a snapshot of morphology every 30,000–50,000 years, and that the changes representing the evolution of a new species have occurred between adjacent samples. A new species will therefore appear to have arisen “suddenly,” but our understanding of the time elapsed defines this “geological instant” as, typically, some tens of thousands of years. This sort of rapidity is what Stephen Jay Gould and Niles Eldredge refer to in their theory of punctuated equilibrium, first proposed in the early 1970s. As they argue, the apparently rapid changes in fossil sequences should be expected, given our knowledge of the absolute timing of the accumulation of sediments forming the rock record and the time intervals typically required for species-level change.
Although Darwin’s bleak assessment of the fossil record is now recognized as overly pessimistic, paleontologists take care to approach the fossil record realistically, knowing that stratigraphic gaps can alter our perceptions of evolutionary patterns and processes (Kidwell and Holland 2002). The recognition of unconformities in the field is therefore critical to the interpretation of fossil sequences. Fortunately, recognition of these gaps between sedimentary strata has occupied the time of a great many stratigraphers who have shown that unconformities can be readily identified and commonly fall at predictable positions within stratigraphic successions.
Thus, the fossil record is neither a complete nor unbiased archive of past life; however, the nature of the fossil record is fairly well understood, and it is possible to design meaningful evolutionary studies using fossil data that are not dominated by the fossil record’s inherently nonrandom structure.
3. MARINE DIVERSITY IN THE PHANEROZOIC
A major focus of paleobiological research over the past 35 years has been the documentation and explanation of biodiversity changes over time. Precambrian life was chemically diverse and often locally abundant, but it generally would not have been much to look at with the naked eye. It is sometimes referred to as the Age of Slime; microbes were dominant, multicellular organisms evolved only after billions of years and for the most part remained tiny; therefore, most paleobiologists interested in the geological history of biodiversity focus on the Phanerozoic eon (the past 542 million years). The name Phanerozoic means obvious life, and indeed the interval is represented by voluminous, widely distributed, fossiliferous sedimentary rocks. The Cambrian explosion occurred in the eponymous earliest period of the Phanerozoic and marks the geologically sudden appearance of large metazoans with readily fossilizable hard parts. Thus, although Cambrian diversity was far from zero, it does mark the Phanerozoic low and provides a good temporal starting point for understanding biodiversity dynamics over evolutionary timescales.
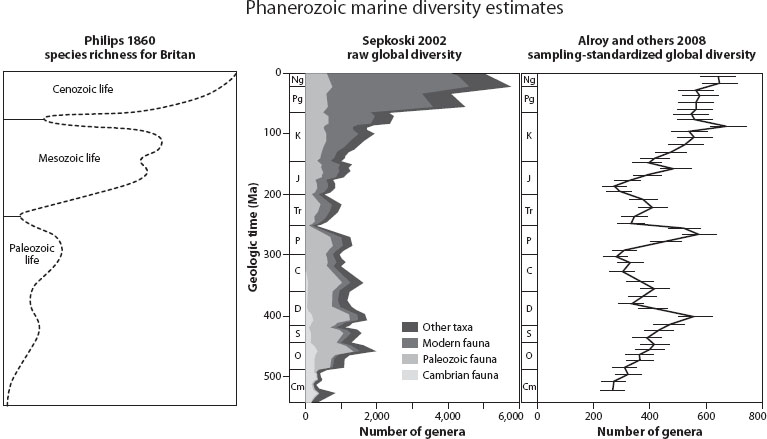
John Phillips, an English geologist of the Victorian era, published one of the first estimates of marine diversity over the Phanerozoic. Although his compilation reflected only taxa known in Great Britain and the diversity axis of his seminal figure bears no numbers, his plot shares many features of more recent compilations. Specifically, Phillips’s figure (figure 1) shows increasing diversity from the earliest Paleozoic to the Holocene, with major drops in diversity at the end-Permian and the end-Cretaceous (recognized today as two of the great mass extinctions, although Phillips did not use that term). These extinction events not only record sharp drops in diversity, but they mark major shifts in the composition of global faunas. It is no accident that these events mark the most fundamental divisions of the Phanerozoic: the Paleozoic, Mesozoic and Cenozoic eras.
Figure 1. Modeled diversity curves showing the number of marine animal taxa through the Phanerozoic. Phillips’s diversity curve is an estimate of the number of species based on observed fossils and the number of outcrops of a given age within Great Britain. Sepkoski’s curve is a global estimate of the number of genera based on times of first and last occurrences. Alroy’s curve is a sampling-standardized global estimate of the number of genera based on the number of genera actually sampled in each time interval; the total ranges of genera are not considered.
In the late twentieth century, new compilations of global marine diversity, particularly by the late J. Jack Sepkoski Jr., were used to explore biodiversity dynamics. Sepkoski trolled the paleontological literature to compile a data set containing the times of first and last appearance of 2800 marine fossil families (later updated to include >36,000 marine genera). The data were global in scope, although with a bias toward North America and Western Europe, where fossil collecting has been most intense. Sepkoski’s family- and genus-level compilations showed very similar diversity trends: an initial slow increase through the Cambrian, a rapid Ordovician radiation, a plateau in diversity through the remainder of the Paleozoic, followed by the major end-Permian mass extinction, with subsequent apparently exponential increase toward the Holocene. Twice during the Paleozoic plateau and twice during the Mesozoic-Cenozoic increase, mass extinctions removed 11–14 percent of the families, including the well-known event at the end of the Cretaceous that defined the extinction of the large terrestrial dinosaurs. Sepkoski’s iconic plot of Phanerozoic diversity (figure 1) is probably the single most frequently used figure in paleontology presentations at national meetings.
Sepkoski did not simply tabulate Phanerozoic marine diversity, he also proposed an evolutionary model to explain the observed patterns. Sepkoski identified three evolutionary faunas: the Cambrian, Paleozoic, and Modern. Each of these three statistically defined evolutionary faunas includes taxa with similar diversity histories and broadly similar ecologies; the Cambrian Fauna is dominated by trilobites and a variety of other small grazers or deposit feeders, the Paleozoic Fauna is characterized by brachiopods, crinoids, and certain types of corals and bryozoans, most of which lived on the sediment surface and fed on suspended material in the water column, and the Modern Fauna (dominant ever since the global ecosystem recovered from the end-Permian mass extinction) is characterized by snails and clams, many of which live below the sediment surface, and a host of predators (including crabs, lobsters, fish, mammals, and reptiles) capable of feeding on organisms with hard skeletons.
Sepkoski used a coupled logistic model of diversification to explain these diversity patterns. In this model, each evolutionary fauna undergoes logistic (density-dependent) growth, meaning that diversification rates are initially exponential but then decline in response to the accumulating diversity. The coupled logistic growth model accounts well for many features of the empirical diversity curve, suggesting that global diversity may move toward equilibrium and that cohorts of higher taxa share common macroevolutionary histories.
Currently, the most controversial aspect of the Sepkoski diversity curve is the shape and magnitude of the Mesozoic-Cenozoic radiation. Sepkoski’s compendium suggests that it is exponential, implying that if the coupled logistic model is correct, the Modern Fauna has not yet reached equilibrium. Others, most notably John Alroy of Macquarie University, have argued that the exponential increase toward the Holocene is a sampling bias. Their primary argument is that the age distribution of sedimentary rocks at the earth’s surface is nonrandom and nonuniform, and that paleontological sampling intensity is dictated by this distribution. (In other words, diversity simply appears to be higher in intervals that have more rocks exposed at the surface.) To test this hypothesis, John Alroy and Charles Marshall created the Paleobiology Database (PaleoDB; paleodb.org) to catalog fossil occurrences from around the globe. Thanks to an ongoing community-wide effort, the PaleoDB currently includes more than 110,000 individual fossil collections comprising more than 950,000 fossil occurrences. The main difference between the PaleoDB and Sepkoski’s data is that Sepkoski reported only the time intervals of a taxon’s first and last appearance, while the PaleoDB records all individual occurrences with the total stratigraphic range (as defined by the oldest and youngest occurrences). Paleontological sampling intensity within each interval can therefore be accounted for in the PaleoDB. The PaleoDB diversity curve (figure 1) shares many of the broad features of the Sepkoski curve, but the rise toward the recent is not exponential and appears to have reached an equilibrium. Although compelling, the PaleoDB diversity curve is certainly not the final “answer.” The nuances of Phanerozoic marine diversity will continue to emerge as the PaleoDB grows, new subsampling techniques are found, and paleoenvironmental data become more comprehensive; however, it seems clear that our overall understanding of past marine diversity is generally good and that logistic (density-dependent) growth models consistently provide the most accurate description of diversification trends. Furthermore, each of the diversity curves produced by Phillips, Sepkoski, and Alroy suggest the possibility that the equilibrium state of diversity has changed over the Phanerozoic.
The other great macroevolutionary advance to come from Sepkoski’s global compendium was recognition of the “big five” mass extinctions and a myriad of smaller extinctions. The earth’s biosphere experienced major mass extinctions at or near the end of the Ordovician, Devonian, Permian, Triassic, and Cretaceous periods. These events, which are covered in more detail in chapter VI.13, were relatively rapid, taxonomically devastating, and global in extent.
4. THE VALUE OF THE FOSSIL RECORD
This chapter offers a very brief introduction to the nature of the fossil record and its importance for understanding the history of life and the evolutionary processes that have shaped that history. Molecular genetics, developmental biology, comparative morphology, and the fossil record all provide powerful and independent evidence for evolution. The beauty of the evidence offered by the fossil record is its accessibility. No matter where you are, a fossiliferous outcrop is likely not too far away. A fossil hunter cannot expect museum-quality specimens at every roadcut, but pulling from the earth a small part of a very ancient trilobite or tree fern or turtle can stir the imagination and awaken a connection to our deep ancestry.
FURTHER READING
Ager, D. V. 1993. The Nature of the Stratigraphical Record. 3rd ed. New York: John Wiley & Sons. An excellent introduction to stratigraphy, including issues of completeness.
Alroy, J. 2010. Geographical, environmental and intrinsic biotic controls on Phanerozoic marine diversification. Palaeontology 53: 1211–1235. The most recent summary of PaleoDB diversity and methods.
Foote, M. 2010. The geological history of biodiversity. In M. A. Bell et al., eds., Evolution since Darwin: The First 150 Years. Sunderland, MA: Sinauer. A review of Phanerozoic marine animal diversity focusing on geological completeness and diversity dependence.
Foote, M., and A. I. Miller. 2007. Principals of Paleontology. 3rd ed. New York: W. H. Freeman. A recent revision of a classic paleobiology textbook.
Gould, S. J. 1989. Wonderful Life: The Burgess Shale and the Nature of History. New York: W. W. Norton. An enjoyable account of the Burgess Shale, with emphasis on evolution as a historical science. Some ideas presented here sparked a controversy over just how strange these organisms were.
Gould, S. J. 2002. The Structure of Evolutionary Theory. Cambridge, MA: Harvard University Press. Gould’s final book. Remarkably long, but with much of value.
Kidwell, S. M., and K. W. Flessa. 1996. The quality of the fossil record: Populations, species, and communities. Annual Reviews in Earth and Planetary Sciences 24: 433–464. A review of the fidelity of the fossil record.
Kidwell, S. M., and S. M. Holland. 2002. The quality of the fossil record: Implications for evolutionary analyses. Annual Review of Ecology and Systematics 33: 561–588. A review of the ways in which stratigraphy influences perceptions of the fossil record.
Knoll, A. H. 2003. Life on a Young Planet: The First Three Billion Years of Evolution on Earth. Princeton, NJ: Princeton University Press. The finest popular book on the fossil record and early life.
Miller, W., III, ed. 2007. Trace Fossils: Concepts, Problems, Prospects. New York: Elsevier. An excellent edited volume on the history, methods, and importance of ichnology, the study of trace fossils.
Raup, D. M. 1979. Biases in the fossil record of species and genera. Bulletin of the Carnegie Museum of Natural History 13: 85–91. A summary of the biases in the fossil record written by one of the most important paleontologists of the last 50 years.
Sepkoski, D., and M. Ruse, eds. 2009. The Paleobiological Revolution: Essays on the Growth of Modern Paleontology. Chicago: University of Chicago Press. Includes some very thoughtful pieces on the development of paleobiology.
Smith, J. M. 1984. Paleontology at the high table. Nature 309: 401–402. An important comment on paleontology by a nonpaleontologist.