Evolution in the Prokaryotic Grade
J. Peter Gogarten and Lorraine Olendzenski
OUTLINE
1. What is a prokaryote?
2. Archaea and Bacteria
3. Rooting the tree of life
4. Symbiosis, syntrophy, and eukaryotic origins
5. Horizontal gene transfer in the evolution of prokaryotes
6. Darwin’s coral of life
7. Biased gene transfer
8. Sex, recombination, and procreation
9. Transfer of genes within and between groups
10. Biochemical innovation as a result of horizontal gene transfer
Prokaryotes are defined as organisms that lack a double membrane-bounded nucleus, but comprise two separate evolutionary lineages, the Archaea (or Archaebacteria) and the Bacteria (or Eubacteria). In prokaryotes and in many single-celled eukaryotes, genes can be transferred between related and unrelated organisms. As a consequence, genes coexisting in a genome have different histories from one another, and organisms can acquire new traits not only through gradual modification of ancestral traits, but also through transfer of genetic material from unrelated organisms.
GLOSSARY
Aminoacyl tRNA Synthetases. Enzymes that charge the tRNA with their cognate amino acid.
Archaea. One of the three domains of life, distinguished based on ribosomal RNA sequence, RNA structure, ether-linked lipids in the cell membrane, and the absence of peptidoglycan in their cell walls. Also known as Archaebacteria.
ATPase/ATP Synthase. Multi-subunit enzymes that use an electrochemical transmembrane gradient of protons to synthesize ATP from ADP and inorganic phosphate. The reaction is reversible, and in some organisms sodium ions are used instead of protons.
Bacteria. One of the three domains of life, characterized by distinct ribosomal structure and rRNA sequence, and cell walls (if present) containing peptidoglycan.
Conjugation. Process of DNA transfer from one cell to another in which bacteria are joined by pili or other structures.
CRISPR Elements (Clustered Regularly Interspaced Short Palindromic Repeats). Regions found in some bacterial genomes and most archaeal genomes that confer immunity against exogenous genetic elements such as phages, plasmids, and other invading genetic elements.
Duplicated Genes. Sets of homologous genes that arose by evolution from each other in an individual or lineage. Also called paralogous genes.
Flagellins. Proteins that make up the bacterial and archaeal flagella.
Gene Transfer Agent (GTA). Particles evolved from phages that transfer host genetic material between prokaryotes and that lost the activity to propagate independently of the host genome.
Halophile. Organism that thrives at high salt concentrations.
Horizontal Gene Transfer (HGT). A process by which genes or gene fragments are transferred among closely or distantly related organisms that are not in a direct ancestor-descendant relationship. It differs from vertical inheritance, whereby offspring acquire genes from their parents. Also known as lateral gene transfer.
Hyperthermophile. Organisms that have optimal growth temperatures from 80 °C to up to 121 °C.
Inteins. Protein introns; intervening sequences similar to introns that are transcribed and translated together with the host protein and that splice out only after translation.
Phage. Virus that attacks Bacteria. Phages are able to introduce foreign genes into a cell.
Plasmids. Circular extrachromosomal DNA elements present in prokaryotes that can replicate independently of the main chromosome. Plasmids carry few nonessential genes.
Prokaryotes. Cellular organisms whose genetic material is not surrounded by a membrane to form a nucleus. They include two domains of life: Archaea and Bacteria.
Recombination. A process in which new DNA is incorporated into a genome. Incorporation happens at regions of similarity between the existing genomic DNA and the incoming DNA. Recombination can be reciprocal, where the introduced DNA replaces an already-existing gene, or illegitimate, whereby newly introduced DNA does not replace existing DNA.
RNA Polymerase. An enzyme that synthesizes RNA using DNA as template.
Rooted Tree of Life. A phylogenetic tree that shows the relationships among all major lineages and includes at its root the most recent common ancestor (MRCA) of all living organisms (also known as last universal common ancestor, or LUCA). The root can be determined using a gene family whose history includes an ancient gene duplication. The phylogeny of such a family including homologues from a variety of different organisms can be rooted by using the ancient paralogues as an out-group.
Thermoacidophile. An organism that is both a thermo- and an acidophile. Many thermoacidophiles thrive at temperatures between 60 and 90 °C and at pH <3. All currently known thermoacidophiles are Archaea.
Thermophile. Organism that grows optimally at temperatures between 50 and 80 °C.
Transformation. Uptake of free DNA from the environment and integration into the genome of a cell.
1. WHAT IS A PROKARYOTE?
Prokaryotes are microorganisms defined by the absence of a nucleus. In eukaryotes (domain Eukarya), the nucleus is created by a double membrane that forms a compartment separating most of the genetic material from the rest of the cell. This compartment exists for most of the cell cycle. The nucleus can be considered a character that evolved in the lineage leading to the eukaryotes. Thus the presence of a nucleus is a derived characteristic of the eukaryotes. The difference in cellular structure between prokaryotes and eukaryotes formed a long-standing perceived dichotomy in the tree of life. However, recognition of the Archaea as a lineage separate from the Bacteria by Carl Woese, George Fox, Otto Kandler, and colleagues forced the realization that prokaryotes comprise at least two major lineages that are superficially similar in cellular morphology, but that do not constitute a monophyletic group. Because the absence of the nucleus reflects the ancestral state, and a group defined by an ancestral state is considered paraphyletic (see chapter II.1), in a cladistic system of classification there is no domain “Prokaryota” (cf. below and the section on taxonomy). The term prokaryote thus describes an organizational level or a grade of evolution; it is useful for describing a group of organisms with similar features but should not be regarded as a valid taxonomic group in a cladistics system.
Prokaryotes are ubiquitous in nature, inhabiting almost all possible niches on earth. It is estimated that there are 1030 prokaryotes on our planet. A single human carries about 1014 prokaryotes, 10 times more than the number of eukaryotic cells in the human body. Prokaryotes are the oldest forms of life on earth; evidence for their existence in the form of laminated rocks called stromatolites stretches back to 3.5 billion years BP, and in the form of graphite-rich sediments to 3.8 billion years BP. They are abundant in ocean water and soil and often predominate in extreme environments where eukaryotes are rare, such as hydrothermal vents, hot springs, hypersaline basins, and the deep subsurface.
Typical unicellular prokaryotes usually range in size from 0.2 μm to about 2.0 μm in diameter. Among the largest is the bacterium Thiomargarita namibiensis, found in the deep sea, whose cells can reach up to 1 mm in diameter. Epulopiscium fishelsoni, a bacterium found in the gut of the surgeon fish, was originally classified as eukaryotic because its cells can be 500 μm long. The morphology of prokaryotes is typically limited to rods, spheres, and filaments; however, some, such as the caulobacters, cyanobacteria, myxococci, and actinobacteria, have evolved complex developmental stages, including multicellularity and cell differentiation. When filamentous forms occur, they are composed of flattened cells stacked together. Resting stages may occur in the form of spores and cysts.
Although the morphological diversity of prokaryotes is relatively limited, their metabolic diversity is a defining characteristic. Prokaryotes carry out unique biochemical processes not found elsewhere in the biosphere, for example, nitrogen fixation, methane production, ammonia oxidation, and various unusual forms of anaerobic respiration, including sulfate, nitrate, and nitrite reduction, as well as the reduction of a variety of metals including iron. Heterotrophic prokaryotes are able to utilize a vast array of organic carbon sources and contribute to the breakdown of complex organic compounds. The nutritional mode of heterotrophic prokaryotes is typically absorptive: they hydrolyze substrates externally and transport small molecules into their cells. Autotrophic prokaryotes are able to fix CO2 into cellular matter using either light energy (phototrophy) or redox energy obtained from reduced inorganic chemicals in a process termed chemoautotrophy. Phototrophy has been found in at least six different bacterial phyla, and all plastids found in eukaryotes can trace their ancestry back to an endosymbiotic event involving a primary endosymbiosis of a eukaryotic cell with a cyanobacterium. Chemoautotrophy is found in diverse phyla of both Bacteria and Archaea.
As a whole, prokaryotes have evolved the ability to couple the oxidation of almost any possible electron donor to the reduction of a huge variety of electron acceptors. In this way, they are able to drive the geochemical transformations of the major biologically reactive elements that make up the biogeochemical cycles of earth.
2. ARCHAEA AND BACTERIA
Initially designated as Eubacteria and Archaebacteria, the two prokaryote groups have been more recently renamed Bacteria and Archaea. Characters that support the division of prokaryotes into two separate lineages include ribosomal RNA sequences, promoter structure, membrane lipid and cell wall composition, antibiotic sensitivity, and the composition of flagellins, the proteins that make up prokaryotic flagella. Phylogenetic trees generated by using molecular sequences of ribosomal proteins, RNA polymerases, tRNAs, proton-pumping ATPases/ATP synthases, elongation factors, and aminoacyl tRNA synthetases typically show Bacteria and Archaea as two separate lineages, influenced by instances of gene transfer.
Archaea
The Archaea are dominated by two well-established phyla, the Euryarchaeota and the Crenarchaeota. Many of the known Euryarchaeota are found in extreme environments and include the halophiles and methanogens. Methanogens gain energy from converting hydrogen (H2) and carbon dioxide into methane. Thermophilic Euryarchaeota are those that grow optimally at temperatures greater than 50 °C and include the thermophilic Archaeoglobi, the thermoacidophilic Thermoplasmata, and the hyperthermophilic Thermococci. The halophilic Archaea, often referred to as haloarchaea, grow best in salt concentrations of 3.5–4.5 molar. Officially their class is still called Halobacteria because they were named before the Archaea were recognized as a distinct group and long before this domain was labeled Archaea.
The first Crenarchaeota were characterized as sulfur-dependent hyperthermophiles, but they have since been found in many other environments. Some theories on the origin of eukaryotes suggest the crenarchaeotes as ancestors of the eukaryotes. In these theories the Crenarchaeota are labeled Eocytes.
Nanoarchaea are extremely small hyperthermophilic cells that are symbiotic with the crenarcheaote Igniococcus and initially were suggested to represent a basal archaeal phylum, the Nanoarchaeota; however, more recent analyses suggest that they are a divergent group within the Euryarchaeota.
The mesophilic Thaumarchaeota, which include marine ammonia oxidizers, were originally seen as a lineage branching within the thermophilic Crenarchaeota. The Thaumarchaeota have been suggested to represent a third major phylum of the Archaea. Thaumarchaeota may possess several ancestral features of the Archaea that have diverged in Crenarchaeota and Euryarchaeota.
The Korarchaeota are another candidate phylum. At present they contain a group of uncultured thermophilic Archaea whose genes in molecular phylogenies often either branch with the Crenarchaeota, or constitute a branch deeper than the split between Crenarchaeota and Euryarchaeota.
Bacteria
More than 50 phyla belonging to the domain Bacteria have been recognized. Approximately half these lineages have no cultured representatives and are recognized solely on the basis of rRNA sequence data obtained from natural environments. Lineages that contain no cultured representatives are termed Candidate Phyla or Candidate Divisions. The Taxonomic Outline of the second edition of Bergey’s Manual of Systematic Bacteriology (Garrity et al. 2004) describes 24 phyla, each with cultured representatives. In August 2011, 30 phyla of Bacteria were recognized in the List of Prokaryotic Names with Standing in Nomenclature. While the phylogenetic analyses of bacterial phyla yields branching patterns among lineages that are unresolved or shrub-like, suggestive of a large simultaneous radiation at the base of the bacterial tree, the placement of individuals into phyla usually is unambiguous.
Many bacterial phylogenies contain two deep branching lineages, Thermotogae and Aquificae, that include mainly thermophiles (organisms that grow optimally at temperatures higher than 50 °C) and hyperthermophiles (organisms that grow optimally from 80 °C up to 121 °C). Other major bacterial lineages include Cyanobacteria, Firmicutes, Actinobacteria, Proteobacteria, Nitrospirae, Bacteriodetes, Chlorobi, Spirochetes, Chlamydiae, Planctomycetes, and Verrucomicrobia. Cyanobacteria are ubiquitous in marine and terrestrial habitats, can have complex morphologies (including filamentous forms with cell specialization), and generate oxygen during photosynthesis (oxygenic photosynthesis). They can be symbiotic with eukaryotic hosts and are the ancestors of eukaryotic plastids. The Firmicutes and Actinobacteria have a specialized cell wall structure characterized by multiple peptidoglycan layers. The major antibiotic-producing bacteria are found within this lineage. The Firmicutes include the endospore-forming Bacillus and Clostridia. Species of Clostridia cause tetanus and botulism. The nonspore-forming Lactobacillus found in fermented foods such as yogurt, pickles, and cheese, as well as probiotic supplements, are also members of the Firmicutes. The Actinobacteria include the filamentous actinomycetes, and nonfilamentous forms such as Mycobacterium, the genus that includes the causative agent of tuberculosis, Propionibacterium, which includes bacteria responsible for the holes in Swiss cheese and acne, and Bifidobacterium, species of which inhabit intestinal tracts and are used in probiotic supplements. One of the best-characterized lineages of Bacteria is Proteobacteria, which is divided into five classes: Alphaproteobacteria (containing nitrogen-fixing endosymbiotic rhizobia, plant-tumor-inducing Agrobacterium, and parasitic Rickettsia), Betaproteobacteria (some photosynthetic bacteria, nitrifying bacteria, and pathogens such as Neisseria, the causative agent of gonorrhea), Gammaproteobacteria (purple sulfur and iron phototrophic bacteria, sulfur-oxidizing bacteria, and the Enterobacteriaceae, such as E. coli and many other bacteria found in the human digestive tract), Deltaproteobacteria (including sulfur- and iron-reducing bacteria and the multicellular Myxobacteria), and Epsilonproteobacteria (Helicobacter, the bacteria that cause ulcers, and some marine sulfur-oxidizing bacteria). The heterotrophic Bacteroidetes and green sulfur-oxidizing phototrophic Chlorobi are species-rich phyla of obligate anaerobes found in many environments. Members of the Nitrospirae are nitrite- and iron-oxidizing bacteria. Spirochetes are responsible for syphilis, Lyme disease, and yaws, and are characterized by unique, sheathed spiral cells with internalized flagella although nonspiral members of this phylum have been characterized. The Chlamydiae are unusual obligate intracellular parasites and form a common sexually transmitted infection in humans, while the Planctomycetes are found in a variety of aquatic environments and exhibit internal membrane compartments reminiscent of nuclei. The Verrucomicrobia are poorly understood microbes that have cell projections containing tubulin, a protein normally found in eukaryotes and believed to have been acquired by these bacteria through horizontal transfer from a eukaryote.
3. ROOTING THE TREE OF LIFE
With the discovery that the Archaea are a separate lineage, a three-domain model of the tree of life emerged. The evolutionary relationship among these three groups has become one of the most important questions in studies of the early evolution of life. One question debated by some is the location of the root of the tree of life, corresponding to the Most Recent Common Ancestor (MRCA) of all extant organisms. While the existence of the Archaea was confirmed by comparing sequences of ribosomal RNA molecules, rRNA sequence analysis alone cannot be used to identify the root of the tree of life since no out-group exists when rRNAs from across all three domains are compared (see chapter II.2). However, sequences from protein families that have undergone ancient gene duplications can be used to root the tree of life, with each paralogue acting as an out-group for the other, producing a reciprocal rooting of the tree. These analyses generally support placing the root on the bacterial branch or within the bacterial domain; however, analyses of some proteins are interpreted to support a rooting along the eukaryal branch or within the Archaea.
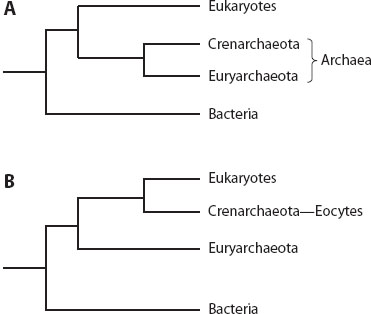
Analyses of most ancient duplicated genes and of the amino acid compositional bias created through the early expansion of the genetic code place the root on the bacterial branch and reveal the Archaea as the lineage most closely related to the eukaryotic nucleocytoplasm (figure 1A). The recognition of the Archaea as a distinct group specifically related to the eukaryotic nucleocytoplasm is widely accepted based on similarities of the transcription machinery, presence of N-linked glycoproteins, lack of formylmethionine, shared resistance or sensitivity to various antibiotics, presence of tRNA introns, histones in some Archaea and most eukaryotes, and similarity between the eukaryotic endomembrane energizing vacuolar ATPase and the archaeal ATP synthase.
Two major hypotheses for the topology of the tree of life are currently seriously debated. The “classical” three-domain model corresponds to a monophyletic Archaea (figure 1A), whereas the alternative “eocyte” model contains a paraphyletic archaeal grouping in which a subset of the Archaea (the Crenarchaeota, sometimes referred to as eocytes) is more closely related to the eukaryotes (figure 1B). Under the classical model (figure 1A), eukaryotes might have accumulated eukaryotic characteristics, such as the cytoskeleton, long before uptake of the mitochondria. The accumulation of numerous derived characters in the eukaryotic lineage, including evolution of the nuclear, cytoskeletal, and spliceosomal machinery, before the endosymbiotic event leading to the mitochondria, provides support for a deep-branching protoeukaryote lineage. However, if some of these derived eukaryotic features, such as the emergence of spliceosomal introns and the nuclear envelope, were triggered by the mitochondrial endosymbiont, or if the premitochondrial derivation of protoeukaryotes was rapid, then a more recent crenarchaeal origin of the Eukarya is plausible, consistent with a tree of life depicting a paraphyletic Archaea (figure 1B).
Figure 1. Possible relationships between the three domains of life. (A) The three-domain model shows a monophyletic archaeal domain (Crenarchaeota and Euryarchaeota). (B) In the eocyte version of the tree of life, the Archaea form a paraphyletic group, with only the Crenarchaeota/Eocytes forming the sister group to the eukaryotic nucleocytoplasm.
4. SYMBIOSIS, SYNTROPHY, AND EUKARYOTIC ORIGINS
Many prokaryotes live inside or on a larger host as mutualists, commensals, or pathogens. Such close symbiotic relationships of prokaryotes with larger organisms led to the acquisition of novel metabolic capabilities by the host, for example, nitrogen fixation in the root nodules of plants or chemoautotrophy in animals such as the giant tube worms and clams found at hydrothermal vents. Symbiosis with prokaryotes can lead to morphological evolution of the host; structures such as the cecum, colon, and rumen have evolved in different mammalian lineages to accommodate the prokaryotic communities that contribute to food digestion. Some deep-sea fish signal mates or attract prey using specialized structures that evolved solely to house luminescent bacteria. Some squid use luminscent bacteria contained in ventral light organs for counterillumination against light radiating from above, making the squid less visible to predators when viewed from below.
Prokaryotes also live in close associations with other prokaryotic members of their communities and engage in a type of metabolic interdependence known as syntrophy. Common in anaerobic environments, syntrophic prokaryotes take advantage of the metabolic abilities of their partners to break down compounds they cannot digest by themselves and overcome energy barriers, allowing them to survive using metabolisms that yield very little free energy for growth. Desulfovibrio vulgaris, which grows well when provided with sulfate, can ferment lactate in the absence of sulfate if the methanogen Methanococcus paludis is also present. When grown together, Desulfovibrio produces acetate, carbon dioxide, and hydrogen from the fermentation of lactate. The methanogen in turn uses the hydrogen produced by Desulfovibrio to reduce carbon dioxide and yield methane. The use of hydrogen by the methanogen drives the overall reaction, which otherwise would not yield energy because of the accumulation of hydrogen.
Syntrophic, endosymbiotic relationships are postulated to have played an important role in the origin of eukaryotes. All extant eukaryotes are the product of lineage fusion that took place at least once early in their evolution, when a bacterium belonging to the Alphaproteobacteria became an endosymbiont in the ancestor of all known eukaryotes. In other words, eukaryotes are derived from at least two distinct ancestors: an organism related to the Archaea was ancestor of the nucleocytoplasm, and an Alphaproteobacterium was the ancestor of the endosymbiont that evolved into the eukaryotic cell organelle known as the mitochondrion. Even eukaryotes that possess no functioning mitochondria have been shown to have evolved from ancestors that already had the alpha-proteobacterial endosymbiont (see chapter II.12). Additional bacterial lineages likely contributed to the origin and early evolution of the eukaryotes; however, the selective advantages that led to the formation and success of these symbioses are still open questions.
5. HORIZONTAL GENE TRANSFER IN THE EVOLUTION OF PROKARYOTES
While the tree of life depicts bifurcating lineages as the standard representation of species evolution, we now understand that evolution is not solely a steadily bifurcating process; lineages also exchange genetic material or fuse with one another to form a new line of descent. The comparison of whole genome sequences from a great variety of prokaryotes has underscored the importance of horizontal gene transfer (HGT) in their evolution.
Phylogenetic trees constructed using single genes can have radically different topologies from one another, depending on what gene is being analyzed. This pattern is consistent with the transfer of genes between unrelated lineages; as a result, prokaryotic genomes are mosaics: different parts of a microbial genome have differing evolutionary histories, largely as a result of horizontal gene transfer. Because of this, phylogenetic reconstruction of organismal evolution is more complicated than in animals, for example, where HGT is rare. For example, in the deeply branching bacterial lineages Aquificae and Thermotogae, the ribosomal protein and RNA gene trees place these lineages as sister to the rest of the Bacteria; however, the majority of genes in the Aquificae group them with the Epsilonproteobacteria, whereas most Thermotogae genes group them among the clostridia, members of the Firmicutes. This suggests large-scale HGT between particular bacterial lineages sometime in the past.
6. DARWIN’S CORAL OF LIFE
The reconstruction of prokaryotic evolution is based mainly on the study of extant organisms. In most molecular phylogenies, only the lineages leading to extant organisms are included. In one of his notebooks, Darwin contemplated that the image of a tree was not appropriate to depict evolution, because in a botanical tree the whole tree is alive, whereas in the tree of life only the top layer represents living organisms. This layer of extant life rests on extinct ancestors, a fact that Darwin noted would be more appropriately captured by the image of a coral. One misconception resulting from the tree image is that the Most Recent Common Ancestor (MRCA) of all extant organisms appears as a singular organism without coexisting lineages. If we take extinct lineages into consideration, it is clear that the MRCA was surrounded by many other lineages. Furthermore, a consequence of gene transfer is that different genes trace their history back to different MRCAs; that is, the MRCA of all extant ribosomes probably existed in a different organism and at a different time than the MRCA of all ATP-synthases, or the MRCA of all extant RNA polymerases.
7. BIASED GENE TRANSFER
Although all genes are subject to horizontal gene transfer, some are more frequently transferred than others, and not all groups of organisms experience gene transfer to the same extent. Such differential gene transfer may contribute to maintaining coherent groups such as species. More closely related organisms often exchange genes more frequently, thus creating a phylogenetic relationship or signal that mimics or reinforces that seen with vertical inheritance. Genes are transferred mainly within major bacterial groups with a transfer bias toward partners more closely related to each other, as judged by rRNA phylogeny. Both shared ancestry (vertical inheritance) and biased HGT seem to cause the observed phylogenetic tendencies observed in genome analyses. Horizontal transfers between divergent species may be draped over a phylogenetic backbone of vertical descent and do not necessarily affect the resolution attainable for a treelike vertical inheritance process of organismal evolution. In prokaryotic lineages, distinguishing the extent to which biased gene transfer contributes to a particular phylogenetic pattern as compared with that of vertical shared ancestry is ongoing.
8. SEX, RECOMBINATION, AND PROCREATION
Sex in most eukaryotes involves the pairing of homologous chromosomes that can then undergo reciprocal crossing-over during meiosis. Daughter cells of a set of meiotic cell divisions receive single copies of each chromosome; overall, the collection of daughter cells contains the full complement of genes in the parent cell, redistributed reciprocally among each homologous chromosome. A generally acknowledged advantage of sexual recombination is that beneficial mutations that arose in different individuals in a population can come together in a single individual in future generations. Without recombination, in order for two separately arising beneficial mutations to come together in the same individual, they would need to evolve successively in the same lineage; however, it must be noted that in many eukaryotic lineages sex is not required for reproduction. Many single-celled eukaryotes, for example, yeast, propagate mainly through mitosis. Even in animals, where reproduction is linked primarily to sex, asexual lineages occur where asexual propagation (parthenogenesis) is the norm (e.g., bdelloid rotifers), with sex occurring only when food becomes scarce (e.g., in the planktonic crustacean Daphnia). In many instances, eukaryotes that propagate asexually have been shown to facilitate recombination by mechanisms similar to those found in prokaryotes.
Recombination in prokaryotes uses different mechanisms from those found in meiotic recombination. DNA can be introduced to a genome via various mobile genetic elements, including plasmids, transposons, integrons, and integrative conjugative elements (ICEs) such as conjugative transposons. Transformation occurs when DNA is taken up from the environment by competent recipient cells. In conjugation, DNA is transferred between two cells that become physically joined by pili. Conjugation can lead to the transfer of either single genes, plasmids, or a substantial portion of the genome. In the cell fusion observed in haloarchaea, two complete genomes are brought together in a single cell, and recombination between the two genomes can occur. Additionally, genetic material can be packaged into phages, viruses that attack bacteria and can be delivered to a recipient cell via a process called transduction. Transduction by phage or virus results in the transfer of foreign DNA into a prokaryotic cell. Gene transfer agents (GTAs) first discovered in some marine Alphaproteobacteria seem to be derived from phage, but they function only in the transfer of genomic DNA between cells and no longer propagate independently from their host. Mechanisms for transferring genes in nature are abundant, but the frequency with which transferred DNAs overcome barriers to attain successful long-term integration into new genomes needs further elucidation.
Gene transfer in prokaryotes is usually unidirectional and not reciprocal. Either a new gene or gene fragment is added to the genome (illegitimate recombination), or it replaces a homologous piece of DNA previously present in the genome (homologous recombination). Usually the replaced fragment is not transferred back to the donor; however, the above-mentioned case of cell fusion in haloarchaea shows that reciprocal recombination in prokaryotes does occur.
Many of the mechanisms for gene transfer may have evolved for purposes unrelated to the acceleration of adaptive evolution through recombination: the immediate selective advantage of taking up DNA from the environment might have been for food; phages, viruses, and conjugative plasmids can thus be considered parasites that have been selected for their own propagation. However, once these mechanisms were in existence, they also allowed for recombination and gene flow within and between populations. Interestingly, virulence resistance via CRISPR elements (Clustered Regularly Interspaced Short Palindromic Repeats) is a trait that greatly decreases the interaction between the bacterial genome, phages, and other extrachromosomal elements; ironically, however, the CRISPR loci themselves are often acquired through the transfer of plasmids.
9. TRANSFER OF GENES WITHIN AND BETWEEN GROUPS
Gene transfer has many advantages for the evolution of groups of organisms. Homologous recombination allows beneficial mutations that occurred in different organisms to come together in a single individual. Similarly, a slightly deleterious mutation can be unlinked through recombination from a beneficial mutation that has occurred in the same individual. However, acquisition of DNA also can be detrimental to the recipient, because the acquired DNA can encode molecular parasites, such as transposons, self-splicing introns, or inteins (protein introns, intervening sequences similar to introns that are transcribed and translated together with the host protein and that splice out only after translation).
The probability of acquiring a molecular parasite is smaller if genes are traded only with close relatives followed by homologous recombination of only small pieces of transferred DNA. The specificity of phages and conjugative plasmids, and the use of recognition sequences by the machinery that imports naked DNA from the environment, decrease the probability of incorporating DNA from an unrelated organism; however, the specificity for within-group transfer is not strict. For example, GTAs can transfer genes to divergent recipients, even to members of different bacterial phyla. Agrobacterium tumefaciens uses its conjugation machinery to transfer tumor-inducing DNA into a plant genome, and many bacteria import DNA without the use of a species-specific recognition sequence.
Prokaryotes possess restriction endonucleases, proteins that act as a kind of immune system to cleave foreign DNA from a virus or phage that may invade a host cell. The smaller fragments created by restriction endonuclease from the foreign DNA can still undergo homologous recombination, but their digestion into smaller pieces decreases the probability of incorporating a complete parasitic genetic element. Methylated DNA is not cleaved by the restriction endonucleases and is thus recognized as belonging to the host. While restriction endonucleases protect against invasion by molecular parasites, the restriction endonuclease and protecting DNA methylase can be considered selfish mobile genetic elements. The protecting DNA methylase has a higher turnover than the restriction endonuclease, and an organism that loses the genes for the restriction/modification system will lose protection faster than the endonuclease activity decays. Under these conditions, the remaining endonuclease activity will attack the genomic DNA of the organism.
Small deletions occur frequently in prokaryotic genomes. If such a deletion inactivates a beneficial gene, the deletion will be negatively selected and removed from the population; however, if the deletion occurs in a parasitic genetic element, the element is inactivated and may be deleted through further deletions. The deletion pressure in prokaryotic genomes tends to inactivate genes that are not under purifying selection in the host; however, this piecewise deletion is circumvented by those selfish genes that lead to addiction, such as the methylating enzyme–restriction enzyme pairs described above.
Sex in eukaryotes restricts recombination to partners belonging to the same species. In contrast, prokaryotes are more promiscuous, frequently incorporating genes from very divergent donors. Despite being more selective in choosing with whom to exchange genes, eukaryotic genomes contain many more remnants of parasitic elements in their genomes than bacteria and archaea. The likely reason for this is the difference in population size. In the case of eukaryotes, the smaller population size allows for less efficient weeding out of mutations with a slight selective disadvantage. The remnant of a molecular parasite increases the amount of DNA and possibly protein that need to be synthesized by a tiny fraction that is too small to be subject to purifying selection in a small population, but that could be weeded out in the very large populations that characterize many prokaryotes.
10. BIOCHEMICAL INNOVATION AS A RESULT OF HORIZONTAL GENE TRANSFER
Horizontal gene transfer plays an important role in the creation of novel biochemical pathways. An existing pathway can be transferred between divergent organisms, providing an adaptive advantage to the recipient; however, in addition, the transfer of genes encoding individual enzymes can create novel pathways that did not previously exist in either the donor or the recipient. Pathways created through gene transfer have greatly impacted the biosphere. One example is acetoclastic methanogenesis in Methanosarcina, which arose through the acquisition of two enzymes from cellulolytic clostridia. These two enzymes in clostridia lead to the production of acetate. In Methanosarcina, these enzymes work in the opposite direction, producing acetate and introducing it into the methanogenic pathway. Today methanogenesis from acetate represents the major pathway for biological methane production, most of it occurring in members of the genus Methanosarcina. Oxygenic photosynthesis is another important process whose assembly is believed to have involved horizontal gene transfer. Chlorophyll-based photosynthesis occurs in several bacterial phyla. In all cases except cyanobacteria, light capture uses only a single photosystem and entails relatively reduced electron donors such as hydrogen sulfide. The photosystems in different phyla are very divergent and can be distinguished on the basis of their sequence and their primary electron acceptor. Only when two of these divergent photosystems came together in the ancestor of the cyanobacteria did it become possible for these photosystems to work in series—generating enough energy to raise electrons from the redox potential of water to that of nicotinamide adenine dinucleotide phosphate (NADP). This allowed cyanobacteria to use water, an effectively unlimited resource, as an electron source, and to generate molecular oxygen as a by-product. Thus, it is fair to say that the oxygen-rich atmosphere we enjoy today would not have arisen without HGT.
FURTHER READING
Bapteste, E., M. A. O’Malley, et al. 2009. Prokaryotic evolution and the tree of life are two different things. Biology Direct 4: 34. Provides the philosophical, scientific, and epistemological perspectives on the reasons a strictly bifurcating phylogenetic tree does not adequately describe evolution in the prokaryotic domains because of extensive chimerism and horizontal gene transfer.
Boekels-Gogarten, M., J. P. Gogarten, and L. Olendzenski, eds. 2009. Horizontal Gene Transfer: Genomes in Flux. Methods in Molecular Biology: 532. New York: Humana Press. Chapters summarizing definitions, philosophy, mechanisms, case studies, and analysis methods used in research on horizontal gene transfer and the impact of HGT on microbial, animal, and plant evolution.
Doolittle, W. F. 1999. Phylogenetic classification and the universal tree. Science 284: 2124–2129. Reviews the reasons a tree-based classification system may be unsuitable for microorganisms.
Dworkin, M., S. Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt, eds. 2006. The Prokaryotes: A Handbook on the Biology of Bacteria. 3rd ed. Vol. 1–7. New York: Springer. Organismal approach to prokaryotic diversity, providing information on habitat, culture techniques, biochemistry, and unique attributes of each group of prokaryotes.
Fournier, G. P., A. A. Dick, D. Williams, and J. P. Gogarten. 2011. Evolution of the Archaea: Emerging views on origins and phylogeny. Research in Microbiology 162: 92–98. Discusses the origin of eukaryotes and their relation to the archaeal phyla.
Fournier, G. P., and J. P. Gogarten. 2010. Rooting the ribosomal tree of life. Molecular Biology and Evolution 27: 1792–1801. Using a phylogenetic tree of concatenated ribosomal proteins, this analysis of amino acid compositional bias detects a strong and unique signal associated with the early expansion of the genetic code, placing the root of the translation machinery along the bacterial branch.
Garrett, R. A., and H.-P. Klenk, eds. 2007. Archaea: Evolution, Physiology, and Molecular Biology. Malden, MA: Blackwell. Documents the early development of the field of study of Archaea by researchers active in their discovery.
Garrity, G. M., et al., eds. 2001–2012. Bergey’s Manual of Systematic Bacteriology. 2nd ed. Vol. 1–5. New York: Springer. Survey of bacterial and archaeal phyla, including information on phylogeny, morphology, ecology, and physiology of cultured species.
Gogarten, J. P., W. F. Doolittle, et al. 2002. Prokaryotic evolution in light of gene transfer. Molecular Biology and Evolution 19: 2226–2238. Discusses biased gene transfer as a force creating exchange groups whose members appear similar to one another, regardless of their shared ancestry.
Margulis, L. 1993. Symbiosis in Cell Evolution: Microbial Communities in the Archean and Proterozoic Eons. 2nd ed. New York: W. H. Freeman. A classic text on the role of endosymbiosis in the evolution of eukaryotic cells.
Oren, A., and R. T. Papke, eds. 2010. Molecular Phylogeny of Microorganisms. Norfolk, UK: Caister Academic Press. Methods and concepts of phylogeny and their usefulness in reconstructing evolutionary relationships of microorganisms, including rooting of the tree of life and impact of horizontal gene transfer on formation of microbial groups.
Pace, N. R. 2009. Mapping the tree of life: Progress and prospects. Microbiology and Molecular Biology Reviews 73: 565–576. Describes current understanding of the structure of the Bacteria and Archaea phylogenetic tree constructed using ribosomal RNA sequences.
Woese, C. R. 1987. Bacterial evolution. Microbiological Reviews 51: 221–271. Summarizes the shortcomings of prior taxonomies and gives an overview of the use of ribosomal RNAs for constructing phylogenetic relationships among Bacteria and Archaea. It also includes an overview of the major groups known at the time.