Adaptive Radiation
Peter R. Grant
OUTLINE
1. Biodiversity
2. Origin and development of the concept
3. The ecological theory
4. Speciation
5. Ecological opportunity
6. Species interactions
7. Intrinsic factors: Key innovations
8. Hybridization
9. Testing the ideas
10. Future prospects
The world has millions of species, and they display an astonishing variety of size, color, and behavior. Adaptive radiations comprise groups of distinctive yet closely related species that have evolved from a common ancestor in a relatively short time. Studies of these radiations help reveal the causes of their evolution. As a result of natural selection during and after speciation, descendant species differ morphologically or physiologically in the way they exploit different environments. Adaptive differentiation also depends on the absence of constraints from competitor species. The guiding force of natural environments is revealed in the observation that the same evolutionary pathway is often taken by different organisms in the same environment. Taxonomic groups vary in their intrinsic potential to diversify because they possess traits that are key evolutionary innovations or because they readily exchange genes through hybridization. Invasion of an underexploited environment allows species to initially multiply at a high rate, and diversify morphologically and ecologically. The fossil record and reconstructions from molecular phylogenies show that both speciation and diversification rates later decline. Experiments in the laboratory with bacteria replicate the pattern of diversification through observable time. Bacteria respond to ecological opportunity by diversifying into a maximum number of ecologically differentiated types.
GLOSSARY
Allopatry. The occurrence of species that occupy separate environments, such as islands.
Competition. The struggle between two or more individuals or species for a resource in limited supply that they jointly consume (e.g., nutrients for a plant, prey for a predator, or places for prey to avoid being eaten). Competition may take the form of an aggressive interaction such as fighting, or differential depletion of a resource by the competitors.
Disparity. The degree of phenotypic difference among individuals or species in one or more traits.
Ecomorph. A recognizable association between morphology of individuals or species and use of the environment.
Evolvability. The genetic and developmental properties of members of a species that determine the likelihood that it will undergo evolutionary change.
Introgressive Hybridization. The interbreeding of two species or genetically divergent populations and subsequent breeding of the offspring with members of one of the parental populations, resulting in the transfer of genes.
Parapatry. The occurrence of species in adjacent or contiguous distributional ranges.
Sympatry. The occurrence of species that occupy the same area.
1. BIODIVERSITY
For many evolutionary biologists the most important pair of questions that need to be answered are: Why do so many species exist on this planet? And why do they differ so greatly from one another? Species number in the millions, varying in size from viruses to whales and from algae to trees; varying in color from bright butterflies to dull and cryptic moths; varying in behavior from solitary polar bears to highly social honey bees; and varying in numerous other ways in exploiting the environment for food, avoiding their enemies, and reproducing. How is all this variation to be explained?
Adaptive radiations provide rich material for seeking answers to these questions because they comprise groups of distinctive yet closely related species. An adaptive radiation is the product of differentiation of an ancestral species into an array of descendant species that differ in the way they exploit the environment. When the differentiation has proceeded rapidly, the evolutionary transitions from one state to another can readily be characterized and strongly interpreted.
Angiosperm plants, dinosaurs, and marsupial mammals are typical examples at high taxonomic levels. Typical examples at lower levels are Darwin’s finches on the Galápagos Islands, honeycreeper finches, Drosophila, spiders, the silversword alliance of plants in the Hawaiian archipelago, cichlid fish in the Great Lakes of Africa, and Anolis lizards in the Caribbean. These examples have the following in common: (1) they comprise several to many species, (2) the species vary morphologically in conspicuous ways, and relatedly, (3) they occupy a diversity of ecological niches. Most of the species were (4) derived from a single ancestor in their current environment, and (5) most diverged relatively rapidly. Cichlid fish in Lake Malawi are an outstanding example. Hundreds of species—the exact number is unknown—were derived from one or a few common ancestors in the last 2 million years, and they have diversified into many trophic forms, including algae-, insect-, snail-, and fish-eating specialists. Their mouth and teeth morphologies reflect their diets, and for this reason the variation is inferred to be adaptive, that is to say, the product of diverse natural selection. One group alone, the rock-dwelling “Mbuna” of the genus Tropheops, comprises 230 species. Ole Seehausen has calculated that one new species arose every 46 years!
2. ORIGIN AND DEVELOPMENT OF THE CONCEPT
The term adaptive radiation was coined in 1902 by a paleontologist, H. F. Osborn, and the phenomenon it refers to was popularized by another, G. G. Simpson, about 50 years later. Simpson viewed the evolutionary radiation of a major group of animals, such as marsupial mammals, as various lines of descent from a common ancestor arising more or less simultaneously and diverging in different morphological and ecological directions, rather like spokes radiating from the hub of a wheel. This image is powerful yet fails to represent the correct evolutionary pathway of bifurcating branches in a treelike structure. Nonetheless, the term has stuck. The adaptive adjective is applied because the products of a radiation are conjectured or known to be adapted to exploiting the environment in different ways.
In the last 20 or 30 years the range of extant organisms that have been studied in detail has increased dramatically, owing largely to the availability of molecular phylogenies for inferring relatedness among species and the pattern and rates of diversification. With these studies has come increasing scrutiny of the term itself, and debate on definitions. Should an unusually high rate of diversification be an essential ingredient of the definition? How is an adaptive radiation to be distinguished from a nonadaptive radiation? These questions become important in comparative studies when generalizations are sought across a broad taxonomic range of organisms. There are no simple answers because there is no clear line of demarcation or break point between adaptive radiations—defined by numbers of species, variety or rates of diversification—and all others. As used in this chapter the term adaptive radiation is most usefully applied to those groups that have diversified rapidly and interpretably, such as the ones cited earlier.
3. THE ECOLOGICAL THEORY
By placing the occurrence of fossils within a time frame, paleontologists like Simpson were able to detect a pattern in the history of a radiation. A radiation was seen to begin with rapid multiplication of species as well as diversification of morphological types. As the radiation proceeded, both species proliferation and morphological evolution slowed down. The observed pattern gave rise to an inferred process, as follows. Invasion of an underexploited environment allowed species to multiply at a high rate. At the same time they diversified phenotypically and ecologically. Eventually, both speciation and diversification rates declined as competition increased for a diminishing variety of unexploited or underexploited resources. This is an explicitly ecological interpretation. On one occasion Simpson used T. H. Huxley’s phrase “filling the ecological barrel” to capture the essence of a limited environment. The greatest opportunity for occupancy occurs at the beginning, when the barrel is empty, and by implication there is an increasing difficulty for newcomers to fit as the barrel fills.
Dolph Schluter converted a coherent explanation into a theory by identifying all the elements and framing them as hypotheses to be tested by their predictions. The key elements of the theory are (1) phenotypic differentiation caused by natural selection arising from differences among environments, (2) competition for resources, and (3) speciation governed by both processes. The theory is not one to be rejected by the first contrary observation so much as an organizational framework for investigating causes in individual cases. This framework has stimulated a large amount of quantitative analysis of large data sets: literally hundreds of species in the case of cichlid fish.
A few words are warranted here about the domain of applicability of the theory. Some groups diversify without radiating. For example, Plethodon salamanders in eastern North America diversified rapidly early in their history, and more slowly later, as in classical adaptive radiations, but speciation was largely allopatric, resulting in little morphological diversity and, as far as is known, relatively little adaptation. The 46 extant species owe their existence to few niches but many habitat fragments and a long history, and not, with some notable exceptions, to competitive interactions. Similarly, speciation may occur repeatedly through diversification of mating signals by sexual selection, such as the color of cichlid fish or petals of flowers (e.g., Mimulus), vocalizations of birds, pheromones of moths, and the stridulation of crickets and leafhoppers. In these and suggested examples of diversification through random drift or founder effects it is difficult to rule out the role of ecological factors in speciation, and for this reason the distinction between adaptive and nonadaptive causes of species proliferation is sometimes blurred. Instead, nonadaptive processes are likely to contribute to an adaptive radiation, being complementary rather than a strict alternative to adaptation. This means that a particular radiation may have heterogeneous causes: adaptive and nonadaptive processes, natural selection and sexual selection, competition and predation, and so forth.
4. SPECIATION
In an adaptive radiation one species gives rise to many. Derived species either do not interbreed or interbreed with a limited degree of genetic exchange (see chapter VI.1). It is easy to envisage how reproductive isolation could arise in an archipelago where populations are isolated by physical barriers and undergo independent evolution: they adapt to different environments. Divergence of the signaling and response system deployed in mate choice occurs as a correlated effect of adaptive divergence. A sympatric phase of the speciation process then follows the allopatric phase, when a derived species disperses into the environment of another. Coexistence in the same habitat, as described by the term sympatry, depends on ecological and reproductive differences having arisen largely or completely in allopatry.
If this were a universal route to species multiplication, those environments without physical barriers should have fewer species, area for area, than fragmented environments, but this is not always observed. For example, relatively homogeneous lakes contain a very high diversity of fish species. In the case of African Great Lake cichlids, their diversity traces back to one or a very few colonizing species, so they must have evolved in broad-scale geographic sympatry within the lake, though possibly in local parapatry or allopatry. Just how a population splits into two under these circumstances is not clear. The question is, How does reproductive isolation evolve through disruptive selection in the face of gene flow counteracting divergence? (See chapter VI.6.) A solution in plants but rare in animals is polyploidy: autopolyploidy through chromosomal doubling, or allopolyploidy through hybridization.
Speciation and adaptive radiation should not be equated. Adaptive radiation usually implies speciation, whereas speciation implies neither adaptation nor a radiation.
5. ECOLOGICAL OPPORTUNITY
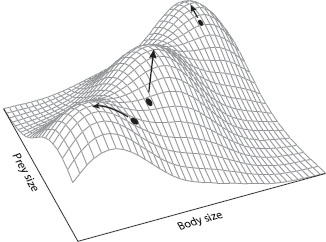
Adaptive differentiation depends on both the availability of ecological resources (niches) to sustain a variety of organisms, and properties of the organisms that facilitate their evolution. Adaptive landscapes provide a way of visualizing opportunities for diversification provided by the environment. The adaptive landscape is an abstraction first developed by Sewall Wright in 1932 to explore possibilities and limits to change of genotype frequencies in a population. Transformed for more general usage by Simpson and others, an adaptive landscape represents variation in fitness in relation to combinations of traits or environmental conditions (figure 1). There are hills and valleys in the landscape. Fitness is at a maximum when a population occupies the top of a hill, be it flat or peaked. Each hill can be equated with ecological opportunity. The spatial distribution of hills reflects another property of environments: some are closer and more within reach of evolving organisms than others, and proximity governs the sequence in which hills can be climbed through the action of natural selection. Neither the environment nor the landscape it represents should be thought of as static. Over time, hills gain or lose height, become more or less peaked, and move closer together or farther apart.
Figure 1. The adaptive landscape. Mean fitness for a phenotype with a given combination of body size and prey size is indicated by the height of the surface. Arrows indicate paths of steepest fitness ascent for three populations in the vicinity of three peaks. (From Schluter 2000.)
The guiding force of natural environments is revealed by such observations as the same evolutionary pathway being taken by different organisms in the same environment. For example, flowering plants from a diverse array of families have invaded freshwater from the land at least 50 times, with repeated evolution of floating, bladderlike structures in the leaves. Dioecy, the presence of two separate sexes in a population, has evolved repeatedly in different lineages of plants colonizing the Hawaiian archipelago. Algal-scraping, mollusk-crushing, fish-scale-scraping cichlid fish and other ecomorphs seen in Lake Malawi are also seen in Lakes Victoria and Tanganyika, even though their origins differ and their evolution has been independent. Thus organisms diversify in response to ecological opportunities. Hand in hand with the opportunity is a challenge from the environment in many cases. For example, coping with ultraviolet radiation and predation from fish and invertebrates is a challenge faced by planktonic Daphnia in the upper waters of lakes. The repeated response of different Daphnia lineages has been the evolution of melanism in the face of the first challenge, and head shields and spines when confronted by the second.
Evolution in similar environments is thus predictable to a degree, because ecological opportunities are similar. An example is provided by the adaptive radiation of Caribbean Anolis lizards. Their history has been reconstructed from a molecular phylogeny. It shows that the four largest islands of the Greater Antilles (Cuba, Hispaniola, Jamaica, and Puerto Rico) were colonized by different species, which then underwent parallel evolutionary diversification into the same ecotypes occupying the same spatial niches (ground, tree trunk, branches, and twigs). Subsequent evolutionary diversification on a given island took the form of variation on these four themes.
Evolutionary history repeats itself at the higher taxonomic levels, too. As pointed out many years ago by Osborn, the marsupial radiation led to convergence in many phenotypic and ecological traits with eutherian mammals: browsing, burrowing, and gliding habits and associated morphological adaptations evolved independently in the two groups, on separate continents. Parallel radiation within two continents yielded convergence between them. This is strong indirect evidence for the driving force of environmental factors in adaptive radiations.
6. SPECIES INTERACTIONS
An important factor in rapid diversification in a new environment is the absence of competitors—typically, related species. In fact, when the concept of adaptive radiation was first developed, the absence of competitors was emphasized as a key facilitating or predisposing factor. For example, islands were viewed as empty environments when the first colonists arrived, and diversification proceeded until all ecological niches were filled, one species per niche. The Gaussian principle that no two species can occupy the same ecological niche is, in modern language, no two species can occupy the same adaptive peak, for reasons of competitive inequality. Therefore, if all peaks in a landscape are occupied, there is no ecological opportunity for a radiation to occur, or if one has begun, there is no opportunity for it to continue.
The objective reality of adaptive peaks can be demonstrated only with environmental data. For granivorous Darwin’s finches in the Galápagos the peaks have been quantified on several islands by measuring food supply, and fitness has been estimated for each island with a given seed size profile by measuring population sizes in relation to beak sizes. The adaptive landscape for these finches is rugged, not smooth. In agreement with expectation, only one species is ever associated with one peak. Interestingly, different species are sometimes associated with the same peak on different islands: they are interchangeable, although ecologically incompatible.
Competitive interactions between species lead either to the exclusion of one species by another or to evolutionary adjustments to each other. Competitive displacement in food-related body size and shape among sticklebacks has been demonstrated experimentally under controlled conditions in ponds. Limnetic species are smaller and more slender than benthic species. A solitary species intermediate in size and shape between them suffered slower growth in the experimental presence of a limnetic species, and as a result, their average body sizes diverged.
Under natural conditions character displacement has been observed in Darwin’s finches. During a drought large members of the medium ground finch population died at a disproportionately high rate. They were outcompeted by a larger, more efficient species, the large ground finch, when feeding on a diminishing supply of large and hard seeds; as a result, the average beak size of the medium ground finch population decreased. Beak size is heritable; therefore, the change produced by natural selection gave rise to evolution: average beak size remained low in the generations produced in the following six years.
7. INTRINSIC FACTORS: KEY INNOVATIONS
Ecological opportunity is one side of a coin; organism responsiveness or intrinsic evolvability is the other. The importance of intrinsic factors is hinted at by differences among taxonomic groups in how far and how fast they radiate. For example, speciation in lacustrine fish appears to be faster than speciation in terrestrial birds and arthropods; time required for speciation has been estimated to be 15,000 to 300,000 years for fish, and half a million to1 million years for birds and arthropods on islands. These values are calculated from simply knowing the number of existing species and then estimating time since they were derived from a common ancestor.
Diversification of cichlid fish has been rampant, but most of the other fish taxa in apparently similar circumstances in African lakes have failed to diversify beyond a few species. Since different environmental factors are not clearly implicated, the contrast suggests there are some intrinsic genetic, developmental, or physiological factors that differ among the groups. Intrinsic factors may potentiate evolutionary change, or they may constrain it. All that is known is that the radiating groups of these fish cluster together in the grand African phylogeny, implying a common inheritance of one or more predisposing factors for radiating. In general, these factors are poorly known, if at all. Identifying them is an important challenge for future studies of adaptive radiations.
The most striking evidence of intrinsic potential is correlative and therefore indirect: it is the association between the evolution of a novel trait and a large number of related species that share the trait. The trait is described as a key evolutionary innovation, facilitating a novel and diverse way of exploiting the environment (see chapter VI.15). Prominent among such novel traits are the pharyngeal jaws of cichlid fish. These are plates with toothlike projections in the roof of the mouth that enable their possessors to split the functions of procuring and processing food into oral and pharyngeal regions of the mouth. Fish with pharyngeal jaws have diversified greatly in oral jaws and associated diets, as different as grazing algae and catching other fish. However, pharyngeal jaws are not sufficient to explain cichlid radiation, because groups that have not radiated also possess them.
The evolution of antifreeze glycoproteins by fish in the suborder Notothenioidei is another example of a key innovation that apparently facilitated invasion of Antarctic waters and subsequent diversification in a relatively empty environment. Detailed comparative work is needed to determine whether a key innovation resulted in an enhanced diversification rate. For example, nectar spurs in columbines (Aquilegia) vary in shape and color, which affect reproductive isolation by attracting different pollinators, and apparently facilitated speciation, because there are more species in clades with than without spurs. The evolutionary invention of resin canals by some plants, constituting a defense against chewing insects, is another example.
8. HYBRIDIZATION
One possible potentiating factor is introgressive hybridization. An exchange of genes can result in enhanced genetic variation and evolutionary potential, and under the right ecological circumstances it can lead to the formation of a new species (see chapter VI.6). The work of Loren Rieseberg on sunflowers (Helianthus) and Tom Whitham on poplars (Populus) in western North America, and several others elsewhere, has shown that introgression of certain genes is selective according to the nature of the environment (soil, microclimate, herbivores, etc.). Populations of sunflower hybrids in peripheral and ecologically extreme environments undergo large-scale genome reorganization, leading to reproductive isolation from the parental populations. These findings have a bearing on the early stages of major radiations on the assumption that what we observe now reveals what happened in the past at the beginning of those radiations. For example, introgressive hybridization is widespread in many young radiations, including the silverswords of Hawaii, Darwin’s finches of the Galápagos, African cichlids, and Heliconius butterflies in South and Central America, but is almost absent in older radiations such as Hawaiian honeycreeper finches and Caribbean Anolis lizards.
Seehausen has suggested that gene mixing through hybridization does more than accompany a radiation: it creates a hybrid swarm and thereby facilitates a radiation. The evidence of ancient hybridization is a mismatch between phylogenetic reconstructions of the same organisms based on nuclear and cytoplasmic genes. For example, the cichlids of Lake Victoria, with a history of 15,000 years, are far younger than the fish in Lake Malawi and correspondingly have much lower mitochondrial gene diversity, yet they have the same level of nuclear diversity as their Lake Malawi counterparts. Similar evidence of incongruence between nuclear and chloroplast genes is found in one lineage of Hawaiian silverswords. The pattern is even repeated in human history. More modern studies of hybridization like the sunflower research are needed to further explore these ideas on diversity generation.
9. TESTING THE IDEAS
Many adaptive radiations happened over millions of years. Their study is therefore retrospective, and interpretations are necessarily inferences that to be plausible, should be consistent with known biological processes. How can the interpretations be tested? For example, in the absence of fossils, how can we tell whether extinction has been important or trivial? An indirect method (see the following section) is to devise alternative models and to use statistical analyses of data to see which model fits the data best. Taxa comprising large numbers of species are necessary for this method to be effective, and well-resolved molecular phylogenies are desirable. And as described later, direct tests can be performed experimentally with microorganisms in the laboratory.
Timing of Speciation and Diversification
The first major pattern of adaptive radiation is an increase in number of species or higher taxa, at a diminishing rate. The second major pattern is an increase in phenotypic disparity, that is, an increase in morphological differences among the species.
The fossil record shows a pattern of high extinction rates at times of geophysical perturbation, followed by speciation that is initially rapid and subsequently slower (see chapter VI.13). Fossils are missing for most living taxa; therefore, molecular phylogenies have been used in their place to see whether the “early burst” phenomenon with subsequent decline is a consistent element of adaptive radiations. A phylogeny describes how the number of species at any one time has increased from the starting point. Accumulation of species on the y-axis is plotted against time on the x-axis to give a lineage-through-time plot. Since the same pattern of accumulation of species diversity can result from a declining speciation rate, as expected, or an increasing extinction rate, it is necessary to decompose diversity into its speciation and extinction components, via mathematical models. For example, Daniel Rabosky and Richard Glor constructed 12 different ways in which species multiplication might be expected to occur as continuous processes of birth (speciation) and death (extinction). Using a molecular phylogeny of Caribbean Anolis lizards to reconstruct the pattern of diversification, they found that one model outperformed others in fitting the data. In this model, speciation rates declined toward a presumed equilibrium on three out of four islands.
However, it is still not known whether the major radiations of the last few million years—which currently are receiving much attention—reflect approximately constant but different speciation and extinction rates, or hidden pulses of each, occurring either synchronously or asynchronously. Agamid lizards in Australia, warblers and Lampropeltine snakes in North America, and pythons in the Australo-Papuan region all declined in rate of diversification. The data fit models with varying speciation rate better than models with varying extinction rate. Conversely, in the most comprehensive analysis to date of many radiations, Jonathan Losos and colleagues found that most of the groups they studied did not fit the early burst pattern. One of several possible reasons is that the broad aspects of the environment do not remain constant, contrary to assumption. For example, Darwin’s finch species increased in parallel with an increase in the number of islands of the Galápagos archipelago. In this case, diversification was facilitated, if not actually driven, by expanding spatial and temporal ecological opportunities. Seehausen found a different pattern of diversification in a changing environment. Species diversity of the rapidly evolving cichlid fish in Africa increased in pulses interspersed with periods of relative stasis, as might be expected if the environment is occasionally perturbed in some significant way. Thus speciation and extinction may not vary in a coordinated manner through time; either or both may change systematically or erratically.
With ecological opportunity being maximal at the beginning of a radiation, initially high rates of phenotypic diversification are expected, which then decrease as the environment becomes progressively filled with a large number of ecologically diverse exploiters. This expectation has been tested with the large number of species that make up the Caribbean Anolis faunas. The expectation was met: rates of diversification of two ecologically important traits, body size and limb length, decreased with increasing radiation, and decreased as the number of inferred potential competitors increased. These results matched paleontological evidence over larger time spans and taxonomic categories: morphological disparity increased among hard-bodied invertebrates in the so-called Cambrian explosion and in certain groups of organisms following the end-Permian and end-Cretaceous mass extinctions.
To what extent are speciation and morphological change coupled? This question is difficult to answer. Branching points in a phylogeny can be estimated, but morphological (and ecological) change independent of speciation can be dated only with fossils, and then with difficulty. Morphological change is likely to accompany speciation (cladogenesis) and be a vital part of it, but change may nonetheless continue without further speciation (anagenesis). Anagenesis may have contributed to diversification of Lampropeltine snakes in North America because morphological disparity increased prior to the Pliocene, close to the time when rates of speciation decreased. Selective extinction through competitive interactions is another process that leads to the same enhancement of morphological disparity.
Experimental Adaptive Radiations
Some of the problems involved in inferring history can be circumvented with studies of living organisms. Direct tests of radiation theory can be performed experimentally with microorganisms in the laboratory; bacteria have the enormous advantage of short generations and rapid evolution. Introduced to a plate of agar, an inoculum of the asexual bacterium Pseudomonas fluorescens diversifies into three main morphological types. Diversification in a microcosm is an analogue of the radiations of sexually reproducing organisms into many species over millennia or more. For example, in the longest-running experiment performed so far, genetic changes affecting metabolism of Escherichia coli have been investigated for more than 50,000 generations. Another advantage of microbes is they can be stored in the freezer, then taken out years later to compare their performance with that of the descendants of the parent population to test the hypothesis that competitive ability evolves. It does.
The Pseudomonas fluorescens experiment shows a repeated pattern of evolutionary diversification into three main ecotypes recognized by their distinctive morphology. A colonizing type (“smooth”) gives rise to two more: a biofilm-producing “wrinkly spreader” and a “fuzzy spreader.” They are specialized on different parts of their environment (spatial niches). Through repeated mutation they give rise to (asexual) clones that are morphological and metabolic variations on these three main themes. The experiments show that the variants within a spatial niche compete for resources and replace one another. The number of morphologically distinct clones (richness) reaches a peak through time and then declines, thus overshooting the long-term carrying capacity of the environment.
The range of morphological variation—the disparity—shows a different pattern: it rises to a maximum and remains there. Thus the bacteria, in the absence of predation and competitor species, respond to ecological opportunity by diversifying into a maximum of ecologically differentiated types. What happens then depends on the environment. Either it remains fixed, as imposed by the investigators, or changes with concomitant changes in the community. Little is known in the laboratory or nature about how a gradually changing environment affects the course of an adaptive radiation, or how adaptive radiation of one group facilitates further radiation through positive feedback from other organisms with which it interacts (e.g., mutualisms). An increase in aridity in the last couple of million years altered the speciation-extinction balance in Caribbean birds. It opened up new niches in the Galápagos, and Darwin’s finches responded by evolving seed-eating specialists.
10. FUTURE PROSPECTS
The study of adaptive radiations is becoming increasingly quantitative, experimental, and comparative. The goal is to understand (1) general properties and (2) differences, according to time of occurrence, taxonomic group, and particular environment. Greater understanding of the causes of diverse adaptive radiations will come from a variety of sources. One is the discovery of new systems. Vertebrates and some plant groups have dominated investigations of extant groups so far, although the recent exploitation of microcosms for experimental investigation has revealed an enormous potential residing in microorganisms. Additional experimental potential at the level of ecological communities has scarcely been tapped. A second source is genetics—specifically, gene expression of ecologically important traits during development—for an understanding of comparative evolvability in different lineages. A third is speciation, introgressive hybridization, and the interrelationship of the two. Experimental investigations have a larger role to play in both revealing and testing the causal factors that observations imply. And inferences about how radiations unfold will improve as analytical methods are refined. Eventually, the knowledge obtained from studying adaptive radiations will be integrated with what will be learned from all the rest of evolution, environments, and earth history, to provide a more comprehensive understanding of the richness of the biological world.
FURTHER READING
Gavrilets, S., and J. B. Losos. 2009. Adaptive radiation: Contrasting theory with data. Science 323: 732–737. Illustrates the interplay of theoretical and empirical studies in the development of understanding how adaptive radiations proceed.
Gavrilets, S. A., and A. Vose. 2005. Dynamic patterns of adaptive radiation. Proceedings of the National Academy of Sciences USA 102: 18040–18045. A theoretical analysis of adaptive radiation.
Givnish, T. J., and K. J. Sytsma, eds. 1997. Molecular Evolution and Adaptive Radiation. Cambridge: Cambridge University Press. Many chapters provide a molecular genetic basis for reconstructing phylogenies and using them for describing and interpreting patterns of adaptive radiation.
Grant, P. R., and B. R. Grant. 2008. How and Why Species Multiply: The Radiation of Darwin’s Finches. Princeton, NJ: Princeton University Press. Darwin’s finches are used as a model system for understanding how species form in adaptive radiations.
Losos, J. B. 2009. Lizards in an Evolutionary Tree. Berkeley: University of California Press. An exceptionally comprehensive account of one of the most impressive and diverse adaptive radiations.
Meyer, J. R., S. E. Schoustra, J. Lachapelle, and R. Kassen. 2011. Overshooting dynamics in a model adaptive radiation. Proceedings of the Royal Society B 278: 392–398. A good example of the power of experimental microcosms for testing in the laboratory ideas generated by observations in nature.
Rabosky, D. L., and R. E. Glor 2010. Equilibrium speciation dynamics in a model adaptive radiation of island lizards. Proceedings of the National Academy of Sciences USA 107: 22178–22183. A comprehensive modeling approach to questions of how adaptive radiations increase in size and complexity through time.
Seehausen, O. 2006. African cichlid fish: A model system in adaptive radiation research. Proceedings of the Royal Society B 273: 1987–1998. A good overview of the most diverse and rapid radiation known.
Schluter, D. 2000. The Ecology of Adaptive Radiation. Oxford: Oxford University Press. The standard modern work on adaptive radiation.
Simpson, G. G. 1953. The Major Features of Evolution. New York: Columbia University Press. A general discussion of evolutionary radiations that integrates population genetics with paleontology.