Sexual Selection: Male-Male Competition
Christine W. Miller
It is certain that amongst almost all animals there is a struggle between the males for the possession of the female. This fact is so notorious that it would be superfluous to give examples. —Charles Darwin
OUTLINE
1. Why are males most often the competing sex and females the choosy sex?
2. The processes of sexual selection
3. Male-male competition in the big and small
4. Weapon evolution
5. Additional forms of male-male competition
6. Male-male competition in plants
7. Total sexual selection
8. Sexual selection and ecological context
Males commonly compete for access to potential mates. This chapter addresses these competitive interactions among males, including alternative mating strategies and sperm competition.
GLOSSARY
Aggression. Offensive action, particularly in regard to defending resources. Aggression in defending females and other resources should be favored by selection when there are fewer resources than competitors and when an individual can boost fitness by forcibly removing another individual.
Intersexual Selection. Sexual selection due to interactions between the sexes. Mate choice by males or females is intersexual selection.
Intrasexual Selection. Sexual selection due to interactions within one sex. Male-male competition for access to mates is the major form of intrasexual selection. In some cases females may also compete for mates, especially in sex-role reversed species.
Polygyny. Mating systems characterized by high variance in male reproductive success; a few males mate with many females, and many males mate with few or no females.
Sperm Competition. Competition between the ejaculates of different males to fertilize the ova of a particular female.
Males in many species compete with one another for access to potential mates, a phenomenon called male-male competition. Sexual selection, or selection due to variation in reproductive success, is traditionally divided into the processes of intrasexual selection—that is, selection between members of one sex for reproductive access to the other sex—and intersexual selection, or mate choice, in which selection of mates is nonrandom. Both sexes can directly compete for matings and exert mate choice, but male-male competition is more apparent than female-female competition, and females are often choosier in selecting mates than are males. Indeed, across species, males are usually the sex with the more elaborated weapons (used in intrasexual competition) and ornaments (used to attract females). When females do compete with one another for males, it is usually in situations where males offer resources such as food, paternal care, or a suitable location to raise offspring.
1. WHY ARE MALES MOST OFTEN THE COMPETING SEX AND FEMALES THE CHOOSY SEX?
The asymmetry between males and females is due to the factors that limit reproductive success for each sex. Individual male reproductive success increases with the number of mating events. Thus, a male can achieve greater reproductive success by competing with other males to access as many females as possible. Females, however, have an upper limit to their potential reproductive success owing to their greater investment in each individual offspring. Thus, it benefits females to choose mates carefully, to ensure that the offspring in which they invest are high quality. When males invest relatively more in caring for offspring than do females, such as is the case in jacana birds, seahorses, giant water bugs, and other sex-role reversed species, females then become the competing sex and males the choosy sex. Furthermore, in monogamous species in which both parents care for offspring and are limited in reproductive potential, both male-male competition and female-female competitions may exist, as well as mate choice by both sexes. See also chapter VII.6.
2. THE PROCESSES OF SEXUAL SELECTION
In The Descent of Man and Selection in Relation to Sex (1874) Charles Darwin noted that “if each male secures two or more females, many males cannot pair.” This observation highlights why sexual selection causes the divergence of male and female traits and explains why male-male competition for reproduction is not unusual. To achieve reproductive opportunities, males must (1) fight other males for access to females or guard the resources females need for survival and reproduction and/or (2) attract females with sexual ornaments and/or direct benefits, such as food. The first situation involves intrasexual selection among males. The second situation describes intersexual selection, involving female choice for indirect (genetic) and direct benefits (see chapter VII.6). These mechanisms of sexual selection rarely operate independently, and thus understanding the entire process of sexual selection requires simultaneous consideration of all the agents that result in differential mating success. Furthermore, because mating success does not perfectly translate into reproductive success, attention to those factors resulting in differential fertilization success (sperm competition and cryptic female choice) is also necessary. In the past, intrasexual selection and intersexual selection have garnered varying amounts of attention each, even though both play central roles in the process of sexual selection. Here, we consider these forces of sexual selection separately to better highlight the important aspects of each to the bigger picture.
3. MALE-MALE COMPETITION IN THE BIG AND SMALL
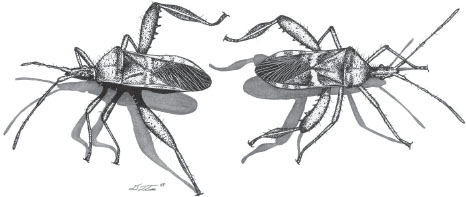
Male-male competition is most commonly observed in polygynous mating systems and where males compete over females (female defense polygyny) or the resources that females need (resource defense polygyny). Examples of male-male competition include the dramatic head butting by male bighorn sheep and fights between roosters. Male elk use elaborate antlers to lock and push one another during the mating season, vying for access to females. But antlers are not limited to ungulates; some male flies also have antlers and competitive behaviors that bear a striking resemblance to those of elk. Another group of flies, the stalk-eyed flies, has eyes that teeter out on the end of long stalks. Males with the longest eyestalks are more likely to win in competitions with other males, and they are also more attractive to females. Leaf-footed cactus bug males engage in wrestling matches by turning around end to end (figure 1), wrapping their elaborated hindlegs around their opponent’s body, and squeezing. These are only a few examples of the wide array of male-male competitive behaviors and associated morphologies found in the natural world.
Figure 1. In an escalated competition male leaf-footed cactus bugs line up end to end, wrap their hindlegs around one another, and press their femur spines into the abdomen of their opponent. (Drawing by David Tuss.)
4. WEAPON EVOLUTION
Male-male competition has resulted in the evolution of a tremendous diversity of weapons across taxa, including spurs, tusks, antlers, horns, and mandibles. In nearly all species that possess sexually selected weapons, essential resources are concentrated in space or time and are thus economically defensible. The costs of investing in weapons can presumably be offset by the benefits of increased reproductive opportunities.
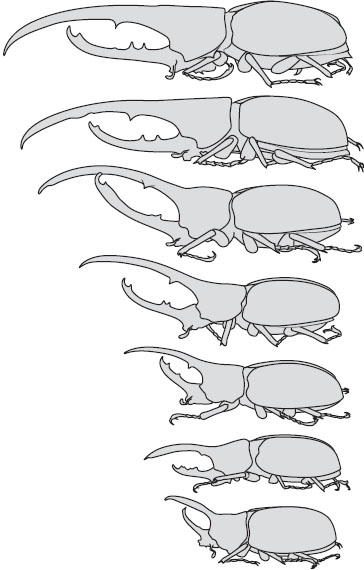
Weapon size can be one of the most variable morphological traits among individuals of the same population (figure 2). Large males or those in good condition generally invest in the largest or most complex weapons, while smaller males often have disproportionately reduced traits. There may be a high cost for a small male to develop a large weapon, both in terms of energetic investment and in terms of severe consequences during escalated competitions. Therefore, these traits are thought to serve as honest indicators of male quality, both prior to and during physical encounters.
Figure 2. Weapon size and shape are often extremely variable within populations. Pictured here: variation in male Dynastes hercules horn size. (Adapted from C. Champy. 1924. Les Caractéres Sexuels Considérés comme Phénomènes de Développement et dans Leurs Rapports avec l’Hormone Sexuelle. Paris: Gaston Doin.)
Species with complex weapons and other indicators of male quality may actually have a reduced, not increased, danger of injury from physical combat. Theoretical models suggest that males should assess the potential of their rivals using visual, chemical, and audible signals and avoid competing with unbeatable opponents. Weapons may serve a dual function as visual signals: the more complex the weapon, the more clearly it may signal a male’s competitive ability. In fact, reconstructions of weapon evolution within clades is beginning to reveal that weapons initially evolve as small, dangerous traits that later evolve to serve more as signals of quality.
Complex weapons may also facilitate male-male contests that are highly ritualized with low risk of lethality. In animals such as elk, dynastid beetles, and even triceratops dinosaurs, male weapons connect in a specific way, allowing for protracted pushing contests that assess strength with little chance of physical damage. For example, baradine weevils have deep pockets in their thorax that accommodate the paired horns of opponent males during contests.
Weapons sometimes exist in both sexes. For example, female cape buffalo have prominent horns, and female tusked wasps have branched facial outgrowths. In most cases, female weapons are smaller than those of their male counterparts and may serve as defense against predators or in competitions with other females over nest sites and food. Existing evidence suggests that most weapons initially evolve owing to natural selection, and male weapons are further elaborated by sexual selection. Evolution commonly proceeds in such a manner—existing features are co-opted for new uses over evolutionary time.
5. ADDITIONAL FORMS OF MALE-MALE COMPETITION
Alternative Mating Strategies
Male-male competition is not limited to obvious direct confrontations. Within a population, some males are able to access females by evading aggressive, dominant males. Males may employ various alternative techniques to come into contact with females, such as superficially resembling females or employing sneaking behaviors. Some of the better-studied examples of species using alternative mating strategies include isopods, ruffs (a wading bird), dung beetles, and Pacific salmon. Males employing alternative mating strategies often also have complementary male morphologies that support the behavioral differences.
When should alternative mating strategies evolve? The answer can be found by examining the average and variance in male mating success for a population. If some males are able to achieve disproportionate access to females, then the average number of mates per male and the variance in male mating success will increase. Sexual selection in this scenario will be much stronger than in a monogamous population, because some males will leave many descendants, while many will leave none. Thus, polygyny can lead to a “mating niche” for males employing unconventional mating behaviors. Males who avoid direct confrontations with other males may still be able to find and mate with females using alternative means.
Males employing alternative mating strategies and achieving surreptitious matings may yield only a fraction of the fertilization success gained by the conventional males, but if they manage to sire even a few offspring, then they will still have greater mating success than conventional males who are unable to secure mates. The average mating success of conventional and unconventional males is often similar, though the variance is often quite different.
Three genetically distinct male types coexist in the marine isopod Paracerceis sculpta. Large alpha males defend females within intertidal sponges, beta males mimic females, and tiny gamma males hide within large harems. All male types have equal mating success, with the beta males and gamma males sneaking copulations. While this mating system is female-defense polygyny, only one of the three male types attempts to defend females. Thus, understanding sexual selection in this species necessitates recognition of alternative mating strategies.
Sperm Competition
Darwin distinguished two contexts for sexual selection: male-male competition and mate choice. He apparently viewed sexual selection as occurring only prior to copulation and that a male’s success could be measured in his ability to obtain copulations. Since Geoff Parker’s work (1970) it has become clear that this view is incomplete. Females commonly mate with more than one male during a reproductive cycle, thus setting the stage for the postcopulatory equivalent of male-male competition, sperm competition, and the postcopulatory equivalent of mate choice, termed cryptic female choice. Sperm competition is the competition between the ejaculates of different males to fertilize the ova of an individual female. Cryptic female choice occurs when females can discriminate in their reproductive tract among the sperm of different males, and at least 21 possible mechanisms for this form of discrimination have been described (see chapter VII.6). Sperm competition and cryptic female choice create powerful selective pressures and have shaped many life-history characteristics including body size, reproductive morphology, physiology, and behavior.
Male Adaptations to Sperm Competition
The single most important factor determining the fertilization success of a male is the number of sperm a male inseminates. Thus, across taxa, males with the potential for more intense sperm competition tend to have larger testes and larger sperm storage organs. Some also have larger accessory glands, which produce the seminal fluid in which the sperm are transported. Accessory glands may produce seminal substances that form copulatory plugs that impede further insemination by other males. Seminal fluids can also contain cocktails of chemicals that increase male fertilization success. In the fruit fly Drosophila, seminal products have been shown to poison previously inseminated sperm and elevate female egg production. These chemicals raise the likelihood of male paternity but come at a cost (to females) of earlier mortality.
Mate guarding is a common and straightforward means of preventing or minimizing sperm competition. Males may guard females before or after mating, or through prolonged copulation. For example, in golden dung flies (Sepsis cynipsea), males locate female on dung pats, guard individual females as they lay eggs, and then attempt to copulate with females after they leave the dung. In squash bugs (Anasa tristis), copulations can last for days, during which the large females pull the small males around by their genitalia. Many males suffer injuries owing to these prolonged copulations, but if greater fertilization success is the result, injury and early death may be well worth the investment.
6. MALE-MALE COMPETITION IN PLANTS
Sexual selection, including both mate choice and male-male competition, is not limited to animals. Plants also compete for mating opportunities. First, male plants must have effective means for moving their pollen to the stigma of a female flower, and they do so using wind, water, or animals. Greater pollen production can help plants disperse their pollen and access ovules. In animal-pollinated plants, male-male competition for the attraction of effective pollinators has played an important role in the evolution of flowers.
Once pollen has reached a stigma, it also must then compete with other pollen for fertilization success, akin to sperm competition in animals. Not all pollen has an equal likelihood of siring seeds. For example, in wild radish (Raphanus sativus), use of genetic markers has shown that unequal siring of seeds by pollen donors is very common. This differential fertilization success may be due to the speed and effectiveness of pollen in reaching ovules, but it may also reflect properties of the female plant that discriminate among potential mates.
7. TOTAL SEXUAL SELECTION
Male-male competition is often studied separately from female mate choice. Experimental isolation of the individual agents of sexual selection is convenient and can provide keen insights into how each component functions, but these studies provide only limited insights into the whole picture. A complete understanding of sexual selection requires simultaneous consideration of male-male competition, female mate choice, and the other agents that affect differential fertilization success in populations, including sperm competition, cryptic female choice, and alternative mating strategies. Mate choice is dealt with in chapter VII.6 but is considered here in relation to its effects on competition for mates.
An example of a dynamic interplay between male-male competition and female mate choice can be seen in the pronghorn antelope, a species with a female defense polygyny mating system. Male pronghorn compete with other males for groups of females, called harems. However, females are not passively herded into harems. Instead, females invest considerable energy sampling many males before choosing a male with which to reproduce. As a result, harem composition fluctuates, and male mating success is associated with the ability to maintain large harems across rut. Sexual selection on males is strong, with only a small subset of males in a population able to achieve mating success in a given year.
As seen in pronghorn, male-male competition and female mate choice commonly select linearly and in the same direction on male traits, such as defensive ability, body size, and weapon size. In some instances, male-male competition and female mate choice may have identical outcomes. For example, females may directly observe male-male competition and mate with the winner of the competitions. However, the agents of sexual selection can also be partially or completely in opposition. For example, in the cockroach Nauphoeta cinerea, a male pheromone is used both in establishing male dominance and in attracting females. However, the blend of compounds most useful for establishing dominance is not the same as the blend found most attractive to females. Thus, there are opposing selection pressures on the composition of a male trait used both in male-male competition and in female mate choice. Such opposing selection pressures in this and other species may lead to the maintenance of genetic variation in sexually selected traits.
What determines whether male-male competition and female mate choice are reinforcing, in opposition, or somewhere in between? One factor appears to be the mating system of a species. For example, in social species, such as primates and cockroaches, males may form dominance hierarchies through male-male competition. Dominant males may have only limited control over female mating decisions, and females may, at times, be able to mate with whomever they prefer. Conversely, in resource defense mating systems, females may be able to reproduce only with the subset of males successful in guarding food and nest sites or those successful in sneaking access to females. Thus, in some species, females may not have the opportunity to fully exercise their mating preferences.
The preceding discussion focused primarily on those situations in which male-male competition and female mate choice act on the same traits in males. Body size is an example of a male trait that is often under selection for both male-male competition and female mate choice. However, the agents of sexual selection may also select for distinct traits. Females may be rather unconcerned about male weapons such as male horns, and males vying for status may ignore ornaments such as colorful plumage. In this case, ornaments and weapons may evolve somewhat independently, albeit with potential trade-offs in investment.
8. SEXUAL SELECTION AND ECOLOGICAL CONTEXT
Theory on mating systems has long recognized that ecological variables—in particular, the concentration of resources—shape the form, strength, and even direction of sexual selection for entire populations (see chapter VII.4). However, this perspective has not fully permeated to considerations of the level of the day-to-day interactions among individuals. The availability and quality of resources and other environmental factors can change quickly, and as they do, individual animals may adaptively change their mate choice and male-male competitive behaviors. For example, in times of resource limitation, otherwise-dominant N. cinerea cockroaches reduce their aggression and dominance behaviors, while subordinate cockroaches become more aggressive. Thus, the structure of dominance hierarchies, access to females, and the resulting sexual selection pressures may vary according to the amount of resources available.
An ecological view of sexual selection necessitates, first and foremost, a keen understanding of the natural history of study organisms, best gained through careful observations in nature. The vast majority of investigations on mate choice and male-male competition have been conducted in only one or a small range of environmental contexts, and often these contexts are wholly artificial. Such experiments allow for fine-tuned analysis of how sexual selection can operate, but they do not provide an ecologically relevant picture of how mate choice and male-male competition function, and fluctuate, in real-world settings. Studies are beginning to demonstrate that sexual selection is indeed variable over time and space. Over the next decades, it will be exciting to learn how environmental variability alters the processes, and outcomes, of sexual selection.
FURTHER READING
Andersson, M. 1994. Sexual Selection. Princeton, NJ: Princeton University Press. A wide-ranging review of sexual selection and mating systems.
Emlen, D. J. 2008. The evolution of animal weapons. Annual Review of Ecology and Systematics 39: 387–413. An extremely readable and fascinating description of animal weapons, including those used by dinosaurs.
Hunt, J., C. J. Breuker, J. A. Sadowski, and A. J. Moore. 2009. Male-male competition, female mate choice and their interaction: Determining total sexual selection. Journal of Evolutionary Biology 22: 12–26. An important reminder that male-male competition and female mate choice may exert opposing or reinforcing selection pressures on male sexually selected traits.
Parker, G. A. 1970. Sperm competition and its evolutionary consequences in the insects. Biological Reviews 45: 525–567. Seminal work by the founder and developer of modern sperm competition theory.
Shuster, S. M., and M. J. Wade. 2003. Mating Systems and Strategies. Princeton, NJ: Princeton University Press. A modern and quantitative synthesis of mating systems and alternative mating strategies.