
Somewhere beyond the edge of our solar system, hurtling into interstellar space, is a phonograph and a golden record with hieroglyphic instructions on the sleeve. They are attached to the Voyager 2 space probe, launched in 1977 to transmit photographs and data back to us from the outer planets in our solar system. Now that it has flown by Neptune and its thrilling scientific mission is over, it serves as an interplanetary calling card from us to any spacefaring extraterrestrial that might snag it.
The astronomer Carl Sagan was the record producer, and he chose sights and sounds that captured our species and its accomplishments. He included greetings in fifty-five human languages and one “whale language,” a twelve-minute sound essay made up of a baby’s cry, a kiss, and an EEG record of the meditations of a woman in love, and ninety minutes of music sampled from the world’s idioms: Mexican mariachi, Peruvian panpipes, Indian raga, a Navajo night chant, a Pygmy girl’s initiation song, a Japanese shakuhachi piece, Bach, Beethoven, Mozart, Stravinsky, Louis Armstrong, and Chuck Berry singing “Johnny B. Goode.”
The disk also bore a message of peace from our species to the cosmos. In an unintended bit of black comedy, the message was recited by the secretary-general of the United Nations at the time, Kurt Waldheim. Years later historians discovered that Waldheim had spent World War II as an intelligence officer in a German army unit that carried out brutal reprisals against Balkan partisans and deported the Jewish population of Salonika to Nazi death camps. It is too late to call Voyager back, and this mordant joke on us will circle the center of the Milky Way galaxy forever.
The Voyager phonograph record, in any case, was a fine idea, if only because of the questions it raised. Are we alone? If not, do alien life forms have the intelligence and the desire to develop space travel? If so, would they interpret the sounds and images as we intended, or would they hear the voice as the whine of a modem and see the line drawings of people on the cover as showing a race of wire frames? If they understood it, how would they respond? By ignoring us? By coming over to enslave us or eat us? Or by starting an interplanetary dialogue? In a Saturday Night Live skit, the long-awaited reply from outer space was “Send more Chuck Berry.”
These are not just questions for late-night dorm-room bull sessions. In the early 1990s NASA allocated a hundred million dollars to a ten-year Search for Extraterrestrial Intelligence (SETI). Scientists were to listen with radio antennas for signals that could have come only from intelligent extraterrestrials. Predictably, some congressmen objected. One said it was a waste of federal money “to look for little green men with mis-shapen heads.” To minimize the “giggle factor,” NASA renamed the project the High-Resolution Microwave Survey, but it was too late to save the project from the congressional ax. Currently it is funded by donations from private sources, including Steven Spielberg.
The opposition to SETI came not just from the know-nothings but from some of the world’s most distinguished biologists. Why did they join the discussion? SETI depends on assumptions from evolutionary theory, not just astronomy—in particular, about the evolution of intelligence. Is intelligence inevitable, or was it a fluke? At a famous conference in 1961, the astronomer and SETI enthusiast Frank Drake noted that the number of extraterrestrial civilizations that might contact us can be estimated with the following formula:
(1) (The number of stars in the galaxy) ×
(2) (The fraction of stars with planets) ×
(3) (The number of planets per solar system with a life-supporting environment) ×
(4) (The fraction of these planets on which life actually appears) ×
(5) (The fraction of life-bearing planets on which intelligence emerges) ×
(6) (The fraction of intelligent societies willing and able to communicate with other worlds) ×
(7) (The longevity of each technology in the communicative state).
The astronomers, physicists, and engineers at the conference felt unable to estimate factor (6) without a sociologist or a historian. But they felt confident in estimating factor (5), the proportion of life-bearing planets on which intelligence emerges. They decided it was one hundred percent.
Finding intelligent life elsewhere in the cosmos would be the most exciting discovery in human history. So why are the biologists being such grinches? It is because they sense that the SETI enthusiasts are reasoning from a pre-scientific folk belief. Centuries-old religious dogma, the Victorian ideal of progress, and modern secular humanism all lead people to misunderstand evolution as an internal yearning or unfolding toward greater complexity, climaxing in the appearance of man. The pressure builds up, and intelligence emerges like popcorn in a pan.
The religious doctrine was called the Great Chain of Being—amoeba to monkey to man—and even today many scientists thoughtlessly use words like “higher” and “lower” life forms and the evolutionary “scale” and “ladder.” The parade of primates, from gangly-armed gibbon through stoop-shouldered caveman to upright modern man, has become an icon of pop culture, and we all understand what someone means when she says she turned down a date because the guy is not very evolved. In science fiction like H. G. Wells’ The Time Machine, episodes of Star Trek, and stories from Boy’s Life, the momentum is extrapolated to our descendants, shown as bald, varicose-veined, bulbous-brained, spindly-bodied homunculi. In The Planet of the Apes and other stories, after we have blown ourselves to smithereens or choked in our pollutants, apes or dolphins rise to the occasion and take on our mantle.
Drake expressed these assumptions in a letter to Science defending SETI against the eminent biologist Ernst Mayr. Mayr had noted that only one of the fifty million species on earth had developed civilizations, so the probability that life on a given planet would include an intelligent species might very well be small. Drake replied:
The first species to develop intelligent civilizations will discover that it is the only such species. Should it be surprised? Someone must be first, and being first says nothing about how many other species had or have the potential to evolve into intelligent civilizations, or may do so in the future. … Similarly, among many civilizations, one will be the first, and temporarily the only one, to develop electronic technology. How else could it be? The evidence does suggest that planetary systems need to exist in sufficiently benign circumstances for a few billion years for a technology-using species to evolve.
To see why this thinking runs so afoul of the modern theory of evolution, consider an analogy. The human brain is an exquisitely complex organ that evolved only once. The elephant’s trunk, which can stack logs, uproot trees, pick up a dime, remove thorns, powder the elephant with dust, siphon water, serve as a snorkel, and scribble with a pencil, is another complex organ that evolved only once. The brain and the trunk are products of the same evolutionary force, natural selection. Imagine an astronomer on the Planet of the Elephants defending SETT, the Search for Extraterrestrial Trunks:
The first species to develop a trunk will discover that it is the only such species. Should it be surprised? Someone must be first, and being first says nothing about how many other species had or have the potential to evolve trunks, or may do so in the future. … Similarly, among many trunk-bearing species, one will be the first, and temporarily the only one, to powder itself with dust. The evidence does suggest that planetary systems need to exist in sufficiently benign circumstances for a few billion years for a trunk-using species to evolve.…
This reasoning strikes us as cockeyed because the elephant is assuming that evolution did not just produce the trunk in a species on this planet but was striving to produce it in some lucky species, each waiting and hoping. The elephant is merely “the first,” and “temporarily” the only one; other species have “the potential,” though a few billion years will have to pass for the potential to be realized. Of course, we are not chauvinistic about trunks, so we can see that trunks evolved, but not because a rising tide made it inevitable. Thanks to fortuitous preconditions in the elephants’ ancestors (large size and certain kinds of nostrils and lips), certain selective forces (the problems posed by lifting and lowering a huge head), and luck, the trunk evolved as a workable solution for those organisms at that time. Other animals did not and will not evolve trunks because in their bodies and circumstances it is of no great help. Could it happen again, here or elsewhere? It could, but the proportion of planets on which the necessary hand has been dealt in a given period of time is presumably small. Certainly it is less than one hundred percent.
We are chauvinistic about our brains, thinking them to be the goal of evolution. And that makes no sense, for reasons articulated over the years by Stephen Jay Gould. First, natural selection does nothing even close to striving for intelligence. The process is driven by differences in the survival and reproduction rates of replicating organisms in a particular environment. Over time the organisms acquire designs that adapt them for survival and reproduction in that environment, period; nothing pulls them in any direction other than success there and then. When an organism moves to a new environment, its lineage adapts accordingly, but the organisms who stayed behind in the original environment can prosper unchanged. Life is a densely branching bush, not a scale or a ladder, and living organisms are at the tips of the branches, not on lower rungs. Every organism alive today has had the same amount of time to evolve since the origin of life—the amoeba, the platypus, the rhesus macaque, and, yes, Larry on the answering machine asking for another date.
But, a SETI fan might ask, isn’t it true that animals become more complex over time? And wouldn’t intelligence be the culmination? In many lineages, of course, animals have become more complex. Life began simple, so the complexity of the most complex creature alive on earth at any time has to increase over the eons. But in many lineages they have not. The organisms reach an optimum and stay put, often for hundreds of millions of years. And those that do become more complex don’t always become smarter. They become bigger, or faster, or more poisonous, or more fecund, or more sensitive to smells and sounds, or able to fly higher and farther, or better at building nests or dams—whatever works for them. Evolution is about ends, not means; becoming smart is just one option.
Still, isn’t it inevitable that many organisms would take the route to intelligence? Often different lineages converge on a solution, like the forty different groups of animals that evolved complex designs for eyes. Presumably you can’t be too rich, too thin, or too smart. Why wouldn’t humanlike intelligence be a solution that many organisms, on this planet and elsewhere, might converge on?
Evolution could indeed have converged on humanlike intelligence several times, and perhaps that point could be developed to justify SETI. But in calculating the odds, it is not enough to think about how great it is to be smart. In evolutionary theory, that kind of reasoning merits the accusation that conservatives are always hurling at liberals: they specify a benefit but neglect to factor in the costs. Organisms don’t evolve toward every imaginable advantage. If they did, every creature would be faster than a speeding bullet, more powerful than a locomotive, and able to leap tall buildings in a single bound. An organism that devotes some of its matter and energy to one organ must take it away from another. It must have thinner bones or less muscle or fewer eggs. Organs evolve only when their benefits outweigh their costs.
Do you have a Personal Digital Assistant, like the Apple Newton? These are the hand-held devices that recognize handwriting, store phone numbers, edit text, send faxes, keep schedules, and many other feats. They are marvels of engineering and can organize a busy life. But I don’t have one, though I am a gadget-lover. Whenever I am tempted to buy a PDA, four things dissuade me. First, they are bulky. Second, they need batteries. Third, they take time to learn to use. Fourth, their sophistication makes simple tasks, like looking up a phone number, slow and cumbersome. I get by with a notebook and a fountain pen.
The same disadvantages would face any creature pondering whether to evolve a humanlike brain. First, the brain is bulky. The female pelvis barely accommodates a baby’s outsize head. That design compromise kills many women during childbirth and requires a pivoting gait that makes women biomechanically less efficient walkers than men. Also, a heavy head bobbing around on a neck makes us more vulnerable to fatal injuries in accidents such as falls. Second, the brain needs energy. Neural tissue is metabolically greedy; our brains take up only two percent of our body weight but consume twenty percent of our energy and nutrients. Third, brains take time to learn to use. We spend much of our lives either being children or caring for children. Fourth, simple tasks can be slow. My first graduate advisor was a mathematical psychologist who wanted to model the transmission of information in the brain by measuring reaction times to loud tones. Theoretically, the neuron-to-neuron transmission times should have added up to a few milliseconds. But there were seventy-five milliseconds unaccounted for between stimulus and response—“There’s all this cogitation going on, and we just want him to push his finger down,” my advisor grumbled. Lower-tech animals can be much quicker; some insects can bite in less than a millisecond. Perhaps this answers the rhetorical question in the sporting equipment ad: The average man’s IQ is 107. The average brown trout’s IQ is 4. So why can’t a man catch a brown trout?
Intelligence isn’t for everyone, any more than a trunk is, and this should give SETI enthusiasts pause. But I am not arguing against the search for extraterrestrial intelligence; my topic is terrestrial intelligence. The fallacy that intelligence is some exalted ambition of evolution is part of the same fallacy that treats it as a divine essence or wonder tissue or all-encompassing mathematical principle. The mind is an organ, a biological gadget. We have our minds because their design attains outcomes whose benefits outweighed the costs in the lives of Plio-Pleistocene African primates. To understand ourselves, we need to know the how, why, where, and when of this episode in history. They are the subject of this chapter.
One evolutionary biologist has made a prediction about extraterrestrial life—not to help us look for life on other planets, but to help us understand life on this planet. Richard Dawkins has ventured that life, anywhere it is found in the universe, will be a product of Darwinian natural selection. That may seem like the most overreaching prognosis ever made from an armchair, but in fact it is a straightforward consequence of the argument for the theory of natural selection. Natural selection is the only explanation we have of how complex life can evolve, putting aside the question of how it did evolve. If Dawkins is right, as I think he is, natural selection is indispensable to understanding the human mind. If it is the only explanation of the evolution of little green men, it certainly is the only explanation of the evolution of big brown and beige ones.
The theory of natural selection—like the other foundation of this book, the computational theory of mind—has an odd status in modern intellectual life. Within its home discipline, it is indispensable, explaining thousands of discoveries in a coherent framework and constantly inspiring new ones. But outside its home, it is misunderstood and reviled. As in Chapter 2, I want to spell out the case for this foundational idea: how it explains a key mystery that its alternatives cannot explain, how it has been verified in the lab and the field, and why some famous arguments against it are wrong.
Natural selection has a special place in science because it alone explains what makes life special. Life fascinates us because of its adaptive complexity or complex design. Living things are not just pretty bits of bric-a-brac, but do amazing things. They fly, or swim, or see, or digest food, or catch prey, or manufacture honey or silk or wood or poison. These are rare accomplishments, beyond the means of puddles, rocks, clouds, and other nonliving things. We would call a heap of extraterrestrial matter “life” only if it achieved comparable feats.
Rare accomplishments come from special structures. Animals can see and rocks can’t because animals have eyes, and eyes have precise arrangements of unusual materials capable of forming an image: a cornea that focuses light, a lens that adjusts the focus to the object’s depth, an iris that opens and closes to let in the right amount of light, a sphere of transparent jelly that maintains the eye’s shape, a retina at the focal plane of the lens, muscles that aim the eyes up-and-down, side-to-side, and in-and-out, rods and cones that transduce light into neural signals, and more, all exquisitely shaped and arranged. The odds are mind-bogglingly stacked against these structures’ being assembled out of raw materials by tornados, landslides, waterfalls, or the lightning bolt vaporizing swamp goo in the philosopher’s thought experiment.
The eye has so many parts, arranged so precisely, that it appears to have been designed in advance with the goal of putting together something that sees. The same is true for our other organs. Our joints are lubricated to pivot smoothly, our teeth meet to sheer and grind, our hearts pump blood—every organ seems to have been designed with a function in mind. One of the reasons God was invented was to be the mind that formed and executed life’s plans. The laws of the world work forwards, not backwards: rain causes the ground to be wet; the ground’s benefiting from being wet cannot cause the rain. What else but the plans of God could effect the teleology (goal-directedness) of life on earth?
Darwin showed what else. He identified a forward-causation physical process that mimics the paradoxical appearance of backward causation or teleology. The trick is replication. A replicator is something that can make a copy of itself, with most of its traits duplicated in the copy, including the ability to replicate in turn. Consider two states of affairs, A and B. B can’t cause A if A comes first. (Seeing well can’t cause an eye to have a clear lens.)
But let’s say that A causes B, and B in turn causes the protagonist of A to make a copy of itself—let’s call it AA. AA looks just like A, so it appears as if B has caused A. But it hasn’t; it has only caused AA, the copy of A. Suppose there are three animals, two with a cloudy lens, one with a clear lens. Having a clear lens (A) causes an eye to see well (B); seeing well causes the animal to reproduce by helping it avoid predators and find mates. The offspring (AA) have clear lenses and can see well, too. It looks as if the offspring have eyes so that they can see well (bad, teleological, backward causation), but that’s an illusion. The offspring have eyes because their parents’ eyes did see well (good, ordinary, forward causation). Their eyes look like their parents’ eyes, so it’s easy to mistake what happened for backward causation.
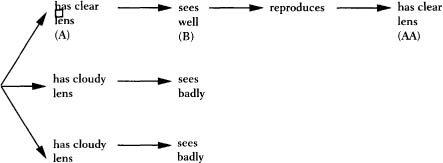
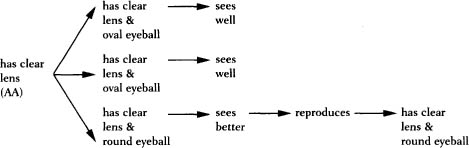
There’s more to an eye than a clear lens, but the special power of a replicator is that its copies can replicate, too. Consider what happens when the clear-lensed daughter of our hypothetical animal reproduces. Some of her offspring will have rounder eyeballs than others, and the round-eyed versions see better because the images are focused from center to edge. Better vision leads to better reproduction, and the next generation has both clear lenses and round eyeballs. They, too, are replicators, and the sharper-visioned of their offspring are more likely to leave a new generation with sharp vision, and so on. In every generation, the traits that lead to good vision are disproportionately passed down to the next generation. That is why a late generation of replicators will have traits that seem to have been designed by an intelligent engineer (see figure on page 158).
I have introduced Darwin’s theory in an unorthodox way that highlights its extraordinary contribution: explaining the appearance of design without a designer, using ordinary forward causation as it applies to replicators. The full story runs as follows. In the beginning was a replicator. This molecule or crystal was a product not of natural selection but of the laws of physics and chemistry. (If it were a product of selection, we would have an infinite regress.) Replicators are wont to multiply, and a single one multiplying unchecked would fill the universe with its great-great-great-…-great-grandcopies. But replicators use up materials to make their copies and energy to power the replication. The world is finite, so the replicators will compete for its resources. Because no copying process is one hundred percent perfect, errors will crop up, and not all of the daughters will be exact duplicates. Most of the copying errors will be changes for the worse, causing a less efficient uptake of energy and materials or a slower rate or lower probability of replication. But by dumb luck a few errors will be changes for the better, and the replicators bearing them will proliferate over the generations. Their descendants will accumulate any subsequent errors that are changes for the better, including ones that assemble protective covers and supports, manipulators, catalysts for useful chemical reactions, and other features of what we call bodies. The resulting replicator with its apparently well-engineered body is what we call an organism.
Natural selection is not the only process that changes organisms over time. But it is the only process that seemingly designs organisms over time. Dawkins stuck out his neck about extraterrestrial evolution because he reviewed every alternative to selection that has been proposed in the history of biology and showed that they are impotent to explain the signature of life, complex design.
The folk theory that organisms respond to an urge to unfold into more complex and adaptive forms obviously won’t do. The urge— and, more important, the power to achieve its ambitions—is a bit of magic that is left unexplained.
The two principles that have come to be associated with Darwin’s predecessor Jean Baptiste Lamarck—use and disuse, and the inheritance of acquired characteristics—are also not up to the job. The problem goes beyond the many demonstrations that Lamarck was wrong in fact. (For example, if acquired traits really could be inherited, several hundred generations of circumcision should have caused Jewish boys today to be born without foreskins.) The deeper problem is that the theory would not be able to explain adaptive complexity even if it had turned out to be correct. First, using an organ does not, by itself, make the organ function better. The photons passing through a lens do not somehow wash it clear, and using a machine does not improve it but wears it out. Now, many parts of organisms do adjust adaptively to use: exercised muscle bulks up, rubbed skin thickens, sunlit skin darkens, rewarded acts increase and punished ones decrease. But these responses are themselves part of the evolved design of the organism, and we need to explain how they arose: no law of physics or chemistry makes rubbed things thicken or illuminated surfaces darken. The inheritance of acquired characteristics is even worse, for most acquired characteristics are cuts, scrapes, scars, decay, weathering, and other assaults by the pitiless world, not improvements. And even if a blow did lead to an improvement, it is mysterious how the size and shape of the helpful wound could be read off the affected flesh and encoded back into DNA instructions in the sperm or egg.
Yet another failed theory is the one that invokes the macromutation: a mammoth copying error that begets a new kind of adapted organism in one fell swoop. The problem here is that the laws of probability astronomically militate against a large random copying error creating a complex functioning organ like the eye out of homogeneous flesh. Small random errors, in contrast, can make an organ a hit more like an eye, as in our example where an imaginable mutation might make a lens a tiny bit clearer or an eyeball a tiny bit rounder. Indeed, way before our scenario begins, a long sequence of small mutations must have accumulated to give the organism an eye at all. By looking at organisms with simpler eyes, Darwin reconstructed how that could have happened. A few mutations made a patch of skin cells light-sensitive, a few more made the underlying tissue opaque, others deepened it into a cup and then a spherical hollow. Subsequent mutations added a thin translucent cover, which subsequently was thickened into a lens, and so on. Each step offered a small improvement in vision. Each mutation was improbable, but not astronomically so. The entire sequence was not astronomically impossible because the mutations were not dealt all at once like a big gin rummy hand; each beneficial mutation was added to a set of prior ones that had been selected over the eons.
A fourth alternative is random genetic drift. Beneficial traits are beneficial only on average. Actual creatures suffer the slings and arrows of outrageous fortune. When the number of individuals in a generation is small enough, an advantageous trait can vanish if its bearers are unlucky, and a disadvantageous or neutral one can take over if its bearers are lucky. Genetic drift can, in principle, explain why a population has a simple trait, like being dark or light, or an inconsequential trait, like the sequence of DNA bases in a part of the chromosome that doesn’t do anything. But because of its very randomness, random drift cannot explain the appearance of an improbable, useful trait like an ability to see or fly. The required organs need hundreds or thousands of parts to work, and the odds are astronomically stacked against the required genes accumulating by sheer chance.
Dawkins’ argument about extraterrestrial life is a timeless claim about the logic of evolutionary theories, about the power of an explanans to cause the explanandum. And indeed his argument works against two subsequent challenges. One is a variant of Lamarckism called directed or adaptive mutation. Wouldn’t it be nice if an organism could react to an environmental challenge with a slew of new mutations, and not wasteful, random ones, but mutations for traits that would allow it to cope? Of course it would be nice, and that’s the problem—chemistry has no sense of niceness. The DNA inside the testes and ovaries cannot peer outside and considerately mutate to make fur when it’s cold and fins when it’s wet and claws when there are trees around, or to put a lens in front of the retina as opposed to between the toes or inside the pancreas. That is why a cornerstone of evolutionary theory—indeed, a cornerstone of the scientific worldview—is that mutations are indifferent overall to the benefits they confer on the organism. They cannot be adaptive in general, though of course a tiny few can be adaptive by chance. The periodic announcements of discoveries of “adaptive mutations” inevitably turn out to be laboratory curiosities or artifacts. No mechanism short of a guardian angel can guide mutations to respond to organisms’ needs in general, there being billions of kinds of organisms, each with thousands of needs.
The other challenge comes from the fans of a new field called the theory of complexity. The theory looks for mathematical principles of order underlying many complex systems: galaxies, crystals, weather systems, cells, organisms, brains, ecosystems, societies, and so on. Dozens of new books have applied these ideas to topics such as AIDS, urban decay, the Bosnian war, and, of course, the stock market. Stuart Kauffman, one of the movement’s leaders, suggested that feats like self-organization, order, stability, and coherence may be an “innate property of some complex systems.” Evolution, he suggests, may be a “marriage of selection and self-organization.”
Complexity theory raises interesting issues. Natural selection presupposes that a replicator arose somehow, and complexity theory might help explain the “somehow.” Complexity theory might also pitch in to explain other assumptions. Each body has to hang together long enough to function rather than fly apart or melt into a puddle. And for evolution to happen at all, mutations have to change a body enough to make a difference in its functioning but not so much as to bring it to a chaotic crash. If there are abstract principles that govern whether a web of interacting parts (molecules, genes, cells) has such properties, natural selection would have to work within those principles, just as it works within other constraints of physics and mathematics like the Pythagorean theorem and the law of gravitation.
But many readers have gone much further and conclude that natural selection is now trivial or obsolete, or at best of unknown importance. (Incidentally, the pioneers of complexity theory themselves, such as Kauffman and Murray Gell-Mann, are appalled by that extrapolation.) This letter to the New York Times Book Review is a typical example:
Thanks to recent advances in nonlinear dynamics, nonequilibrium thermodynamics and other disciplines at the boundary between biology and physics, there is every reason to believe that the origin and evolution of life will eventually be placed on a firm scientific footing. As we approach the 21st century, those other two great 19th century prophets—Marx and Freud—have finally been deposed from their pedestals. It is high time we freed the evolutionary debate from the anachronistic and unscientific thrall of Darwin worship as well.
The letter-writer must have reasoned as follows: complexity has always been treated as a fingerprint of natural selection, but now it can be explained by complexity theory; therefore natural selection is obsolete. But the reasoning is based on a pun. The “complexity” that so impresses biologists is not just any old order or stability. Organisms are not just cohesive blobs or pretty spirals or orderly grids. They are machines, and their “complexity” is functional, adaptive design: complexity in the service of accomplishing some interesting outcome. The digestive tract is not just patterned; it is patterned as a factory line for extracting nutrients from ingested tissues. No set of equations applicable to everything from galaxies to Bosnia can explain why teeth are found in the mouth rather than in the ear. And since organisms are collections of digestive tracts, eyes, and other systems organized to attain goals, general laws of complex systems will not suffice. Matter simply does not have an innate tendency to organize itself into broccoli, wombats, and ladybugs. Natural selection remains the only theory that explains how adaptive complexity, not just any old complexity, can arise, because it is the only nonmiraculous, forward-direction theory in which how well something works plays a causal role in how it came to be.

Because there are no alternatives, we would almost have to accept natural selection as the explanation of life on this planet even if there were no evidence for it. Thankfully, the evidence is overwhelming. I don’t just mean evidence that life evolved (which is way beyond reasonable doubt, creationists notwithstanding), but that it evolved by natural selection. Darwin himself pointed to the power of selective breeding, a direct analogue of natural selection, in shaping organisms. For example, the differences among dogs—Chihuahuas, greyhounds, Scotties, Saint Bernards, shar-peis—come from selective breeding of wolves for only a few thousand years. In breeding stations, laboratories, and seed company greenhouses, artificial selection has produced catalogues of wonderful new organisms befitting Dr. Seuss.
Natural selection is also readily observable in the wild. In a classic example, the white peppered moth gave way in nineteenth-century Manchester to a dark mutant form after industrial soot covered the lichen on which the moth rested, making the white form conspicuous to birds. When air pollution laws lightened the lichen in the 1950s, the then-rare white form reasserted itself. There are many other examples, perhaps the most pleasing coming from the work of Peter and Rosemary Grant. Darwin was inspired to the theory of natural selection in part by the thirteen species of finches on the Galápagos islands. They clearly were related to a species on the South American mainland, but differed from them and from one another. In particular, their beaks resembled different kinds of pliers: heavy-duty lineman’s pliers, high-leverage diagonal pliers, straight needle-nose pliers, curved needle-nose pliers, and so on. Darwin eventually reasoned that one kind of bird was blown to the islands and then differentiated into the thirteen species because of the demands of different ways of life on different parts of the islands, such as stripping bark from trees to get at insects, probing cactus flowers, or cracking tough seeds. But he despaired of ever seeing natural selection happen in real time: “We see nothing of these slow changes in progress, until the hand of time has marked the lapse of ages.” The Grants painstakingly measured the size and toughness of the seeds in different parts of the Galápagos at different times of the year, the length of the finches’ beaks, the time they took to crack the seeds, the numbers and ages of the finches in different parts of the islands, and so on—every variable relevant to natural selection. Their measurements showed the beaks evolving to track changes in the availability of different kinds of seeds, a frame-by-frame analysis of the movie that Darwin could only imagine. Selection in action is even more dramatic among faster-breeding organisms, as the world is discovering to its peril in the case of pesticide-resistant insects, drug-resistant bacteria, and the AIDS virus in a single patient.
And two of the prerequisites of natural selection—enough variation and enough time—are there for the having. Populations of naturally living organisms maintain an enormous reservoir of genetic variation that can serve as the raw material for natural selection. And life has had more than three billion years to evolve on earth, complex life a billion years, according to a recent estimate. In The Ascent of Man, Jacob Bronowski wrote:
I remember as a young father tiptoeing to the cradle of my first daughter when she was four or five days old, and thinking, “These marvelous fingers, every joint so perfect, down to the fingernails. I could not have designed that detail in a million years.” But of course it is exactly a million years that it took me, a million years that it took mankind … to reach its present stage of evolution.
Finally, two kinds of formal modeling have shown that natural selection can work. Mathematical proofs from population genetics show how genes combining according to Gregor Mendel’s laws can change in frequency under the pressure of selection. These changes can occur impressively fast. If a mutant produces just 1 percent more offspring than its rivals, it can increase its representation in a population from 0.1 percent to 99.9 percent in just over four thousand generations. A hypothetical mouse subjected to a selection pressure for increased size that is so weak it cannot be measured could nonetheless evolve to the size of an elephant in only twelve thousand generations.
More recently, computer simulations from the new field of Artificial Life have shown the power of natural selection to evolve organisms with complex adaptations. And what better demonstration than everyone’s favorite example of a complex adaptation, the eye? The computer scientists Dan Nilsson and Susanne Pelger simulated a three-layer slab of virtual skin resembling a light-sensitive spot on a primitive organism. It was a simple sandwich made up of a layer of pigmented cells on the bottom, a layer of light-sensitive cells above it, and a layer of translucent cells forming a protective cover. The translucent cells could undergo random mutations of their refractive index: their ability to bend light, which in real life often corresponds to density. All the cells could undergo small mutations affecting their size and thickness. In the simulation, the cells in the slab were allowed to mutate randomly, and after each round of mutation the program calculated the spatial resolution of an image projected onto the slab by a nearby object. If a bout of mutations improved the resolution, the mutations were retained as the starting point for the next bout, as if the slab belonged to a lineage of organisms whose survival depended on reacting to looming predators. As in real evolution, there was no master plan or project scheduling. The organism could not put up with a less effective detector in the short run even if its patience would have been rewarded by the best conceivable detector in the long run. Every change it retained had to be an improvement.
Satisfyingly, the model evolved into a complex eye right on the computer screen. The slab indented and then deepened into a cup; the transparent layer thickened to fill the cup and bulged out to form a cornea. Inside the clear filling, a spherical lens with a higher refractive index emerged in just the right place, resembling in many subtle details the excellent optical design of a fish’s eye. To estimate how long it would take in real time, rather than in computer time, for an eye to unfold, Nilsson and Pelger built in pessimistic assumptions about heritability, variation in the population, and the size of the selective advantage, and even forced the mutations to take place in only one part of the “eye” each generation. Nonetheless, the entire sequence in which flat skin became a complex eye took only four hundred thousand generations, a geological instant.
I have reviewed the modern case for the theory of natural selection because so many people are hostile to it. I don’t mean fundamentalists from the Bible Belt, but professors at America’s most distinguished universities from coast to coast. Time and again I have heard the objections: the theory is circular, what good is half an eye, how can structure arise from random mutation, there hasn’t been enough time, Gould has disproved it, complexity just emerges, physics will make it obsolete someday.
People desperately want Darwinism to be wrong. Dennett’s diagnosis in Darwin’s Dangerous Idea is that natural selection implies there is no plan to the universe, including human nature. No doubt that is a reason, though another is that people who study the mind would rather not have to think about how it evolved because it would make a hash of cherished theories. Various scholars have claimed that the mind is innately equipped with fifty thousand concepts (including “carburetor” and “trombone”), that capacity limitations prevent the human brain from solving problems that are routinely solved by bees, that language is designed for beauty rather than for use, that tribal people kill their babies to protect the ecosystem from human overpopulation, that children harbor an unconscious wish to copulate with their parents, and that people could just as easily be conditioned to enjoy the thought of their spouse being unfaithful as to be upset by the thought. When advised that these claims are evolutionarily improbable, they attack the theory of evolution rather than rethinking the claim. The efforts that academics have made to impugn Darwinism are truly remarkable.
One claim is that reverse-engineering, the attempt to discover the functions of organs (which I am arguing should be done to the human mind), is a symptom of a disease called “adaptationism.” Apparently if you believe that any aspect of an organism has a function, you absolutely must believe that every aspect has a function, that monkeys are brown to hide amongst the coconuts. The geneticist Richard Lewontin, for example, has defined adaptationism as “that approach to evolutionary studies which assumes without further proof that all aspects of the morphology, physiology and behavior of organisms are adaptive optimal solutions to problems.” Needless to say, there is no such madman. A sane person can believe that a complex organ is an adaptation, that is, a product of natural selection, while also believing that features of an organism that are not complex organs are a product of drift or a by-product of some other adaptation. Everyone acknowledges that the redness of blood was not selected for itself but is a by-product of selection for a molecule that carries oxygen, which just happens to be red. That does not imply that the ability of the eye to see could easily be a by-product of selection for something else.
There also are no benighted fools who fail to realize that animals carry baggage from their evolutionary ancestors. Readers young enough to have had sex education or old enough to be reading articles about the prostate may have noticed that the seminal ducts in men do not lead directly from the testicles to the penis but snake up into the body and pass over the ureter before coming back down. That is because the testes of our reptilian ancestors were inside their bodies. The bodies of mammals are too hot for the production of sperm, so the testes gradually descended into a scrotum. Like a gardener who snags a hose around a tree, natural selection did not have the foresight to plan the shortest route. Again, that does not mean that the entire eye could very well be useless phylogenetic baggage.
Similarly, because adaptationists believe that the laws of physics are not enough to explain the design of animals, they are also imagined to be prohibited from ever appealing to the laws of physics to explain anything. A Darwin critic once defiantly asked me, “Why has no animal evolved the ability to disappear and instantly reappear elsewhere, or to turn into King Kong at will (great for frightening predators)?” I think it is fair to say that “not being able to turn into King Kong at will” and “being able to see” call for different kinds of explanations.
Another accusation is that natural selection is a sterile exercise in after-the-fact storytelling. But if that were true, the history of biology would be a quagmire of effete speculation, with progress having to wait for today’s enlightened anti-adaptationists. Quite the opposite has happened. Mayr, the author of a definitive history of biology, wrote,
The adaptationist question, “What is the function of a given structure or organ?” has been for centuries the basis of every advance in physiology. If it had not been for the adaptationist program, we probably would still not yet know the functions of thymus, spleen, pituitary, and pineal. Harvey’s question “Why are there valves in the veins?” was a major stepping stone in his discovery of the circulation of blood.
From the shape of an organism’s body to the shape of its protein molecules, everything we have learned in biology has come from an understanding, implicit or explicit, that the organized complexity of an organism is in the service of its survival and reproduction. This includes what we have learned about the nonadaptive by-products, because they can be found only in the course of a search for the adaptations. It is the bald claim that a feature is a lucky product of drift or of some poorly understood dynamic that is untestable and post hoc.
Often I have heard it said that animals are not well engineered after all. Natural selection is hobbled by shortsightedness, the dead hand of the past, and crippling constraints on what kinds of structures are biologically and physically possible. Unlike a human engineer, selection is incapable of good design. Animals are clunking jalopies saddled with ancestral junk and occasionally blunder into barely serviceable solutions.
People are so eager to believe this claim that they seldom think it through or check the facts. Where do we find this miraculous human engineer who is not constrained by availability of parts, manufacturing practicality, and the laws of physics? Of course, natural selection does not have the foresight of engineers, but that cuts both ways: it does not have their mental blocks, impoverished imagination, or conformity to bourgeois sensibilities and ruling-class interests, either. Guided only by what works, selection can home in on brilliant, creative solutions. For millennia, biologists have discovered to their astonishment and delight the ingenious contrivances of the living world: the biomechanical perfection of cheetahs, the infrared pinhole cameras of snakes, the sonar of bats, the superglue of barnacles, the steel-strong silk of spiders, the dozens of grips of the human hand, the DNA repair machinery in all complex organisms. After all, entropy and more malevolent forces like predators and parasites are constantly gnawing at an organism’s right to life and do not forgive slapdash engineering.
And many of the examples of bad design in the animal kingdom turn out to be old spouses’ tales. Take the remark in a book by a famous cognitive psychologist that natural selection has been powerless to eliminate the wings of any bird, which is why penguins are stuck with wings even though they cannot fly. Wrong twice. The moa had no trace of a wing, and penguins do use their wings to fly—under water. Michael French makes the point in his engineering textbook using a more famous example:
It is an old joke that a camel is a horse designed by a committee, a joke which does grave injustice to a splendid creature and altogether too much honour to the creative power of committees. For a camel is no chimera, no odd collection of bits, but an elegant design of the tightest unity. So far as we can judge, every part is contrived to suit the difficult role of the whole, a large herbivorous animal to live in harsh climates with much soft going, sparse vegetation and very sparse water. The specification for a camel, if it were ever written down, would be a tough one in terms of range, fuel economy and adaptation to difficult terrains and extreme temperatures, and we must not be surprised that the design that meets it appears extreme. Nevertheless, every feature of the camel is of a piece: the large feet to diffuse load, the knobbly knees that derive from some of the design principles of Chapter 7 [bearings and pivots], the hump for storing food and the characteristic profile of the lips have a congruity that derives from function and invests the whole creation with a feeling of style and a certain bizarre elegance, borne out by the beautiful rhythms of its action at a gallop.
Obviously, evolution is constrained by the legacies of ancestors and the kinds of machinery that can be grown out of protein. Birds could not have evolved propellers, even if that had been advantageous. But many claims of biological constraints are howlers. One cognitive scientist has opined that “many properties of organisms, like symmetry, for example, do not really have anything to do with specific selection but just with the ways in which things can exist in the physical world.” In fact, most things that exist in the physical world are not symmetrical, for obvious reasons of probability: among all the possible arrangements of a volume of matter, only a tiny fraction are symmetrical. Even in the living world, the molecules of life are asymmetrical, as are livers, hearts, stomachs, flounders, snails, lobsters, oak trees, and so on. Symmetry has everything to do with selection. Organisms that move in straight lines have bilaterally symmetrical external forms because otherwise they would go in circles. Symmetry is so improbable and difficult to achieve that any disease or defect can disrupt it, and many animals size up the health of prospective mates by checking for minute asymmetries.
Gould has emphasized that natural selection has only limited freedom to alter basic body plans. Much of the plumbing, wiring, and architecture of the vertebrates, for example, has been unchanged for hundreds of millions of years. Presumably they come from embryological recipes that cannot easily be tinkered with. But the vertebrate body plan accommodates eels, cows, hummingbirds, aardvarks, ostriches, toads, gerbils, seahorses, giraffes, and blue whales. The similarities are important, but the differences are important, too! Developmental constraints only rule out broad classes of options. They cannot, by themselves, force a functioning organ to come into being. An embryological constraint like “Thou shalt grow wings” is an absurdity. The vast majority of hunks of animal flesh do not meet the stringent engineering demands of powered flight, so it is infinitesimally unlikely that the creeping and bumping cells in the microscopic layers of the developing embryo are obliged to align themselves into bones, skin, muscles, and feathers with just the right architecture to get the bird aloft—unless, of course, the developmental program had been shaped to bring about that outcome by the history of successes and failures of the whole body.
Natural selection should not be pitted against developmental, genetic, or phylogenetic constraints, as if the more important one of them is, the less important the others are. Selection versus constraints is a phony dichotomy, as crippling to clear thinking as the dichotomy between innateness and learning. Selection can only select from alternatives that are growable as carbon-based living stuff, but in the absence of selection that stuff could just as easily grow into scar tissue, scum, tumors, warts, tissue cultures, and quivering amorphous protoplasm as into functioning organs. Thus selection and constraints are both important but are answers to different questions. The question “Why does this creature have such-and-such an organ?” by itself is meaningless. It can only be asked when followed by a compared-to-what phrase. Why do birds have wings (as opposed to propellers)? Because you can’t grow a vertebrate with propellers. Why do birds have wings (as opposed to forelegs or hands or stumps)? Because selection favored ancestors of birds that could fly.
Another widespread misconception is that if an organ changed its function in the course of evolution, it did not evolve by natural selection. One discovery has been cited over and over in support of the misconception: the wings of insects were not originally used for locomotion. Like a friend-of-a-friend legend, that discovery has mutated in the retelling: wings evolved for something else but happened to be perfectly adapted for flight, and one day the insects just decided to fly with them; the evolution of insect wings refutes Darwin because they would have had to evolve gradually and half a wing is useless; the wings of birds were not originally used for locomotion (probably a misremembering of another fact, that the first feathers evolved not for flight but for insulation). All one has to do is say “the evolution of wings” and audiences will nod knowingly, completing the anti-adaptationist argument for themselves. How can anyone say that any organ was selected for its current function? Maybe it evolved for something else and the animal is only using it for that function now, like the nose holding up spectacles and all that stuff about insect wings that everyone knows about (or was it bird wings?).
Here is what you find when you check the facts. Many organs that we see today have maintained their original function. The eye was always an eye, from light-sensitive spot to image-focusing eyeball. Others changed their function. That is not a new discovery. Darwin gave many examples, such as the pectoral fins of fishes becoming the forelimbs of horses, the flippers of whales, the wings of birds, the digging claws of moles, and the arms of humans. In Darwin’s day the similarities were powerful evidence for the fact of evolution, and they still are. Darwin also cited changes in function to explain the problem of “the incipient stages of useful structures,” perennially popular among creationists. How could a complex organ gradually evolve when only the final form is usable? Most often the premise of unusability is just wrong. For example, partial eyes have partial sight, which is better than no sight at all. But sometimes the answer is that before an organ was selected to assume its current form, it was adapted for something else and then went through an intermediate stage in which it accomplished both. The delicate chain of middle-ear bones in mammals (hammer, anvil, stirrup) began as parts of the jaw hinge of reptiles. Reptiles often sense vibrations by lowering their jaws to the ground. Certain bones served both as jaw hinges and as vibration transmitters. That set the stage for the bones to specialize more and more as sound transmitters, causing them to shrink and move into their current shape and role. Darwin called the earlier forms “pre-adaptations,” though he stressed that evolution does not somehow anticipate next year’s model.
There is nothing mysterious about the evolution of birds’ wings. Half a wing will not let you soar like an eagle, but it will let you glide or parachute from trees (as many living animals do), and it will let you leap or take off in bursts while running, like a chicken trying to escape a farmer. Paleontologists disagree about which intermediate stage is best supported by the fossil and aerodynamic evidence, but there is nothing here to give comfort to a creationist or a social scientist.
The theory of the evolution of insect wings proposed by Joel Kingsolver and Mimi Koehl, far from being a refutation of adaptationism, is one of its finest moments. Small cold-blooded animals like insects struggle to regulate their temperature. Their high ratio of surface area to volume makes them heat up and cool down quickly. (That is why there are no bugs outside in cold months; winter is the best insecticide.) Perhaps the incipient wings of insects first evolved as adjustable solar panels, which soak up the sun’s energy when it is colder out and dissipate heat when it’s warmer. Using thermodynamic and aerodynamic analyses, Kingsolver and Koehl showed that proto-wings too small for flight are effective heat exchangers. The larger they grow, the more effective they become at heat regulation, though they reach a point of diminishing returns. That point is in the range of sizes in which the panels could serve as effective wings. Beyond that point, they become more and more useful for flying as they grow larger and larger, up to their present size. Natural selection could have pushed for bigger wings throughout the range from no wings to current wings, with a gradual change of function in the middle sizes.
So how did the work get garbled into the preposterous story that one day an ancient insect took off by flapping unmodified solar panels and the rest of them have been doing it ever since? Partly it is a misunderstanding of a term introduced by Gould, exaptation, which refers to the adaptation of an old organ to a new function (Darwin’s “pre-adaptation”) or the adaptation of a non-organ (bits of bone or tissue) to an organ with a function. Many readers have interpreted it as a new theory of evolution that has replaced adaptation and natural selection. It’s not. Once again, complex design is the reason. Occasionally a machine designed for a complicated, improbable task can be pressed into service to do something simpler. A book of cartoons called 101 Uses for a Dead Computer showed PCs being used as a paperweight, an aquarium, a boat anchor, and so on. The humor comes from the relegation of sophisticated technology to a humble function that cruder devices can fulfill. But there will never be a book of cartoons called 101 Uses for a Dead Paperweight showing one being used as a computer. And so it is with exaptation in the living world. On engineering grounds, the odds are against an organ designed for one purpose being usable out of the box for some other purpose, unless the new purpose is quite simple. (And even then the nervous system of the animal must often be adapted for it to find and keep the new use.) If the new function is at all difficult to accomplish, natural selection must have revamped and retrofitted the part considerably, as it did to give modern insects their wings. A housefly dodging a crazed human can decelerate from rapid flight, hover, turn in its own length, fly upside down, loop, roll, and land on the ceiling, all in less than a second. As an article entitled “The Mechanical Design of Insect Wings” notes, “Subtle details of engineering and design, which no man-made airfoil can match, reveal how insect wings are remarkably adapted to the acrobatics of flight.” The evolution of insect wings is an argument for natural selection, not against it. A change in selection pressure is not the same as no selection pressure.
Complex design lies at the heart of all these arguments, and that offers a final excuse to dismiss Darwin. Isn’t the whole idea a bit squishy? Since no one knows the number of kinds of possible organisms, how can anyone say that an infinitesimal fraction of them have eyes? Perhaps the idea is circular: the things one calls “adaptively complex” are just the things that one believes couldn’t have evolved any other way than by natural selection. As Noam Chomsky wrote,
So the thesis is that natural selection is the only physical explanation of design that fulfills a function. Taken literally, that cannot be true. Take my physical design, including the property that I have positive mass. That fulfills some function—namely, it keeps me from drifting into outer space. Plainly, it has a physical explanation which has nothing to do with natural selection. The same is true of less trivial properties, which you can construct at will. So you can’t mean what you say literally. I find it hard to impose an interpretation that doesn’t turn it into the tautology that where systems have been selected to satisfy some function, then the process is selection.
Claims about functional design, because they cannot be stated in exact numbers, do leave an opening for a skeptic, but a little thought about the magnitudes involved closes it. Selection is not invoked to explain mere usefulness; it’s invoked to explain improbable usefulness. The mass that keeps Chomsky from floating into outer space is not an improbable condition, no matter how you measure the probabilities. “Less trivial properties”—to pick an example at random, the vertebrate eye—are improbable conditions, no matter how you measure the probabilities. Take a dip net and scoop up objects from the solar system; go back to life on the planet a billion years ago and sample the organisms; take a collection of molecules and calculate all their physically possible configurations; divide the human body into a grid of one-inch cubes. Calculate the proportion of samples that have positive mass. Now calculate the proportion of samples that can form an optical image. There will be a statistically significant difference in the proportions, and it needs to be explained.
At this point the critic can say that the criterion—seeing versus not seeing—is set a posteriori, after we know what animals can do, so the probability estimates are meaningless. They are like the infinitesimal probability that I would have been dealt whatever poker hand I happened to have been dealt. Most hunks of matter cannot see, but then most hunks of matter cannot flern either, where I hereby define flern as the ability to have the exact size and shape and composition of the rock I just picked up.
Recently I visited an exhibition on spiders at the Smithsonian. As I marveled at the Swiss-watch precision of the joints, the sewing-machine motions by which it drew silk from its spinnerets, the beauty and cunning of the web, I thought to myself, “How could anyone see this and not believe in natural selection!” At that moment a woman standing next to me exclaimed, “How could anyone see this and not believe in God!” We agreed a priori on the facts that need to be explained, though we disagreed about how to explain them. Well before Darwin, theologians such as William Paley pointed to the engineering marvels of nature as proof of the existence of God. Darwin did not invent the facts to be explained, only the explanation.
But what, exactly, are we all so impressed by? Everyone might agree that the Orion constellation looks like a big guy with a belt, but that does not mean we need a special explanation of why stars align themselves into guys with belts. But the intuition that eyes and spiders show “design” and that rocks and Orion don’t can be unpacked into explicit criteria. There has to be a heterogeneous structure: the parts or aspects of an object are unpredictably different from one another. And there has to be a unity of function: the different parts are organized to cause the system to achieve some special effect—special because it is improbable for objects lacking that structure, and special because it benefits someone or something. If you can’t state the function more economically than you can describe the structure, you don’t have design. A lens is different from a diaphragm, which in turn is different from a photopigment, and no unguided physical process would deposit the three in the same object, let alone align them perfectly. But they do have something in common—all are needed for high-fidelity image formation—and that makes sense of why they are found together in an eye. For the flerning rock, in contrast, describing the structure and stating the function are one and the same. The notion of function adds nothing.
And most important, attributing adaptive complexity to natural selection is not just a recognition of design excellence, like the expensive appliances in the Museum of Modern Art. Natural selection is a falsifiable hypothesis about the origin of design and imposes onerous empirical requirements. Remember how it works: from competition among replicators. Anything that showed signs of design but did not come from a long line of replicators could not be explained by—in fact, would refute—the theory of natural selection: natural species that lacked reproductive organs, insects growing like crystals out of rocks, television sets on the moon, eyes spewing out of vents on the ocean floor, caves shaped like hotel rooms down to the details of hangers and ice buckets. Moreover, the beneficial functions all have to be in the ultimate service of reproduction. An organ can be designed for seeing or eating or mating or nursing, but it had better not be designed for the beauty of nature, the harmony of the ecosystem, or instant self-destruction. Finally, the beneficiary of the function has to be the replicator. Darwin pointed out that if horses had evolved saddles, his theory would immediately be falsified.
Rumors and folklore notwithstanding, natural selection remains the heart of explanation in biology. Organisms can be understood only as interactions among adaptations, by-products of adaptations, and noise. The by-products and noise don’t rule out the adaptations, nor do they leave us staring blankly, unable to tell them apart. It is exactly what makes organisms so fascinating—their improbable adaptive design—that calls for reverse-engineering them in the light of natural selection. The by-products and noise, because they are defined negatively as un-adaptations, also can be discovered only via reverse-engineering.
This is no less true for human intelligence. The major faculties of the mind, with their feats no robot can duplicate, show the handiwork of selection. That does not mean that every aspect of the mind is adaptive. From low-level features like the sluggishness and noisiness of neurons, to momentous activities like art, music, religion, and dreams, we should expect to find activities of the mind that are not adaptations in the biologists’ sense. But it does mean that our understanding of how the mind works will be woefully incomplete or downright wrong unless it meshes with our understanding of how the mind evolved. That is the topic of the rest of the chapter.
Why did brains evolve to start with? The answer lies in the value of information, which brains have been designed to process.
Every time you buy a newspaper, you are paying for information. Economic theorists have explained why you should: information confers a benefit that is worth paying for. Life is a choice among gambles. One turns left or right at the fork in the road, stays with Rick or leaves with Victor, knowing that neither choice guarantees fortune or happiness; the best one can do is play the odds. Stripped to its essentials, every decision in life amounts to choosing which lottery ticket to buy. Say a ticket costs $1.00 and offers a one-in-four chance of winning $10.00. On average, you will net $1.50 per play ($10.00 divided by 4 equals $2.50, minus $1.00 for the ticket). The other ticket costs $1.00 and offers a one-in-five chance of winning $12.00. On average, you will net $1.40 per play. The two kinds of tickets come in equal numbers, and neither has the odds or winnings marked on it. How much should you pay for someone to tell you which is which? You should pay up to four cents. With no information, you would have to choose at random, and you could expect to make $1.45 on average ($1.50 half the time, $1.40 half the time). If you knew which had the better average payoff, you would make an average of $1.50 each play, so even if you paid four cents you would be ahead by one cent each play.
Most organisms don’t buy lottery tickets, but they all choose between gambles every time their bodies can move in more than one way. They should be willing to “pay” for information—in tissue, energy, and time—if the cost is lower than the expected payoff in food, safety, mating opportunities, and other resources, all ultimately valuated in the expected number of surviving offspring. In multicellular animals the information is gathered and translated into profitable decisions by the nervous system.
Often, more information brings a greater reward and earns back its extra cost. If a treasure chest has been buried somewhere in your neighborhood, the single bit of information that locates it in the north or the south half is helpful, because it cuts your digging time in half. A second bit that told you which quadrant it was in would be even more useful, and so on. The more digits there are in the coordinates, the less time you will waste digging fruitlessly, so you should be willing to pay for more bits, up to the point where you are so close that further subdivision would not be worth the cost. Similarly, if you were trying to crack a combination lock, every number you bought would cut down the number of possibilities to try, and could be worth its cost in the time saved. So very often more information is better, up to a point of diminishing returns, and that is why some lineages of animals have evolved more and more complex nervous systems.
Natural selection cannot directly endow an organism with information about its environment, or with the computational networks, demons, modules, faculties, representations, or mental organs that process the information. It can only select among genes. But genes build brains, and different genes build brains that process information in different ways. The evolution of information processing has to be accomplished at the nuts-and-bolts level by selection of genes that affect the brain-assembly process.
Many kinds of genes could be the targets of selection for better information processing. Altered genes could lead to different numbers of proliferative units along the walls of the ventricles (the cavities in the center of the brain), which beget the cortical neurons making up the gray matter. Other genes could allow the proliferative units to divide for different numbers of cycles, creating different numbers and kinds of cortical areas. Axons connecting the neurons can be re-routed by shifting the chemical trails and molecular guideposts that coax the axons in particular directions. Genes can change the molecular locks and keys that encourage neurons to connect with other ones. As in the old joke about how to carve a statue of an elephant (remove all the bits that don’t look like an elephant), neural circuits can be sculpted by programming certain cells and synapses to commit suicide on cue. Neurons can become active at different points in embryogenesis, and their firing patterns, both spontaneous and programmed, can be interpreted downstream as information about how to wire together. Many of these processes interact in cascades. For example, increasing the size of one area allows it to compete better for real estate downstream. Natural selection does not care how baroque the brain-assembly process is, or how ugly the resulting brain. Modifications are evaluated strictly on how well the brain’s algorithms work in guiding the perception, thought, and action of the whole animal. By these processes, natural selection can build a better and better functioning brain.
But could the selection of random variants really improve the design of a nervous system? Or would the variants crash it, like a corrupted byte in a computer program, and the selection merely preserve the systems that do not crash? A new field of computer science called genetic algorithms has shown that Darwinian selection can create increasingly intelligent software. Genetic algorithms are programs that are duplicated to make multiple copies, though with random mutations that make each one a tiny bit different. All the copies have a go at solving a problem, and the ones that do best are allowed to reproduce to furnish the copies for the next round. But first, parts of each program are randomly mutated again, and pairs of programs have sex: each is split in two, and the halves are exchanged. After many cycles of computation, selection, mutation, and reproduction, the surviving programs are often better than anything a human programmer could have designed.
More apropos of how a mind can evolve, genetic algorithms have been applied to neural networks. A network might be given inputs from simulated sense organs and outputs to simulated legs and placed in a virtual environment with scattered “food” and many other networks competing for it. The ones that get the most food leave the most copies before the next round of mutation and selection. The mutations are random changes in the connection weights, sometimes followed by sexual recombination between networks (swapping some of their connection weights). During the early iterations, the “animals”—or, as they are sometimes called, “animats”—wander randomly over the terrain, occasionally bumping into a food source. But as they evolve they come to zip directly from food source to food source. Indeed, a population of networks that is allowed to evolve innate connection weights often does better than a single neural network that is allowed to learn them. That is especially true for networks with multiple hidden layers, which complex animals, especially humans, surely have. If a network can only learn, not evolve, the environmental teaching signal gets diluted as it is propagated backward to the hidden layers and can only nudge the connection weights up and down by minuscule amounts. But if a population of networks can evolve, even if they cannot learn, mutations and recombinations can reprogram the hidden layers directly, and can catapult the network into a combination of innate connections that is much closer to the optimum. Innate structure is selected for.
Evolution and learning can also go on simultaneously, with innate structure evolving in an animal that also learns. A population of networks can be equipped with a generic learning algorithm and can be allowed to evolve the innate parts, which the network designer would ordinarily have built in by guesswork, tradition, or trial and error. The innate specs include how many units there are, how they are connected, what the initial connection weights are, and how much the weights should be nudged up and down on each learning episode. Simulated evolution gives the networks a big head start in their learning careers.
So evolution can guide learning in neural networks. Surprisingly, learning can guide evolution as well. Remember Darwin’s discussion of “the incipient stages of useful structures”—the what-good-is-half-an-eye problem. The neural-network theorists Geoffrey Hinton and Steven Nowlan invented a fiendish example. Imagine an animal controlled by a neural network with twenty connections, each either excitatory (on) or neutral (off). But the network is utterly useless unless all twenty connections are correctly set. Not only is it no good to have half a network; it is no good to have ninety-five percent of one. In a population of animals whose connections are determined by random mutation, a fitter mutant, with all the right connections, arises only about once every million (220) genetically distinct organisms. Worse, the advantage is immediately lost if the animal reproduces sexually, because after having finally found the magic combination of weights, it swaps half of them away. In simulations of this scenario, no adapted network ever evolved.
But now consider a population of animals whose connections can come in three forms: innately on, innately off, or settable to on or off by learning. Mutations determine which of the three possibilities (on, off, learnable) a given connection has at the animal’s birth. In an average animal in these simulations, about half the connections are learnable, the other half on or off. Learning works like this. Each animal, as it lives its life, tries out settings for the learnable connections at random until it hits upon the magic combination. In real life this might be figuring out how to catch prey or crack a nut; whatever it is, the animal senses its good fortune and retains those settings, ceasing the trial and error. From then on it enjoys a higher rate of reproduction. The earlier in life the animal acquires the right settings, the longer it will have to reproduce at the higher rate.
Now with these evolving learners, or learning evolvers, there is an advantage to having less than one hundred percent of the correct network. Take all the animals with ten innate connections. About one in a thousand (210) will have all ten correct. (Remember that only one in a million nonlearning animals had all twenty of its innate connections correct.) That well-endowed animal will have some probability of attaining the completely correct network by learning the other ten connections; if it has a thousand occasions to learn, success is fairly likely. The successful animal will reproduce earlier, hence more often. And among its descendants, there are advantages to mutations that make more and more of the connections innately correct, because with more good connections to begin with, it takes less time to learn the rest, and the chances of going through life without having learned them get smaller. In Hinton and Nowlan’s simulations, the networks thus evolved more and more innate connections. The connections never became completely innate, however. As more and more of the connections were fixed, the selection pressure to fix the remaining ones tapered off, because with only a few connections to learn, every organism was guaranteed to learn them quickly. Learning leads to the evolution of innateness, but not complete innateness.
Hinton and Nowlan submitted the results of their computer simulations to a journal and were told that they had been scooped by a hundred years. The psychologist James Mark Baldwin had proposed that learning could guide evolution in precisely this way, creating an illusion of Lamarckian evolution without there really being Lamarckian evolution. But no one had shown that the idea, known as the Baldwin effect, would really work. Hinton and Nowlan showed why it can. The ability to learn alters the evolutionary problem from looking for a needle in a haystack to looking for the needle with someone telling you when you are getting close.
The Baldwin effect probably played a large role in the evolution of brains. Contrary to standard social science assumptions, learning is not some pinnacle of evolution attained only recently by humans. All but the simplest animals learn. That is why mentally uncomplicated creatures like fruit flies and sea slugs have been convenient subjects for neuroscientists searching for the neural incarnation of learning. If the ability to learn was in place in an early ancestor of the multicellular animals, it could have guided the evolution of nervous systems toward their specialized circuits even when the circuits are so intricate that natural selection could not have found them on its own.
Complex neural circuitry has evolved in many animals, but the common image of animals climbing up some intelligence ladder is wrong. The common view is that lower animals have a few fixed reflexes, and that in higher ones the reflexes can be associated with new stimuli (as in Pavlov’s experiments) and the responses can be associated with rewards (as in Skinner’s). On this view, the ability to associate gets better in still higher organisms, and eventually it is freed from bodily drives and physical stimuli and responses and can associate ideas directly to each other, reaching an apex in man. But the distribution of intelligence in real animals is nothing like this.
The Tunisian desert ant leaves its nest, travels some distance, and then wanders over the burning sands looking for the carcass of an insect that has keeled over from the heat. When it finds one, it bites off a chunk, turns, and makes a beeline for the nest, a hole one millimeter in diameter as much as fifty meters away. How does it find its way back? The navigation depends on information gathered during the outward journey, not on sensing the nest like a beacon. If someone lifts the ant as it emerges from the nest and plunks it down some distance away, the ant wanders in random circles. If someone moves the ant after it finds food, it runs in a line within a degree or two of the direction of its nest with respect to the abduction site, slightly overshoots the point where the nest should be, does a quick U-turn, and searches for the nonexistent nest. This shows that the ant has somehow measured and stored the direction and distance back to the nest, a form of navigation called path integration or dead reckoning.
This example of information processing in animals, discovered by the biologist Rudiger Wehner, is one of many that the psychologist Randy Gallistel has used to try to get people to stop thinking about learning as the formation of associations. He explains the principle:
Path integration is the integration of the velocity vector with respect to time to obtain the position vector, or some discrete equivalent of this computation. The discrete equivalent in traditional marine navigation is to record the direction and speed of travel (the velocity) at intervals, multiply each recorded velocity by the interval since the previous recording to get interval-by-interval displacements (e.g., making 5 knots on a northeast course for half an hour puts the ship 2.5 nautical miles northeast of where it was), and sum the successive displacements (changes in position) to get the net change in position. These running sums of the longitudinal and latitudinal displacements are the deduced reckoning of the ship’s position.
Audiences are incredulous. All that computation inside the little bitty pinhead of an ant? Actually, as computation goes, this is pretty simple stuff; you could build a device to do it for a few dollars out of little parts hanging on the pegboard at Radio Shack. But intuitions about the nervous system have been so impoverished by associationism that a psychologist would be accused of wild, profligate speculation if she were to attribute this machinery to a human brain, let alone an ant brain. Could an ant really do calculus, or even arithmetic? Not overtly, of course, but then neither do we when we exercise our own faculty of dead reckoning, our “sense of direction.” The path integration calculations are done unconsciously, and their output pokes into our awareness—and the ant’s, if it has any—as an abstract feeling that home is thataway, yea far.
Other animals execute even more complicated sequences of arithmetic, logic, and data storage and retrieval. Many migratory birds fly thousands of miles at night, maintaining their compass direction by looking at the constellations. As a Cub Scout I was taught how to find the North Star: locate the tip of the handle of the Little Dipper, or extrapolate from the front lip of the Big Dipper a distance seven times its depth. Birds are not born with this knowledge, not because it is unthinkable that it could be innate, but because if it were innate it would soon be obsolete. The earth’s axis of rotation, and hence the celestial pole (the point in the sky corresponding to north), wobbles in a 27,000-year cycle called the precession of the equinoxes. The cycle is rapid in an evolutionary timetable, and the birds have responded by evolving a special algorithm for learning where the celestial pole is in the night sky. It all happens while they are still in the nest and cannot fly. The nestlings gaze up at the night sky for hours, watching the slow rotation of the constellations. They find the point around which the stars appear to move, and record its position with respect to several nearby constellations, acquiring the information imparted to me by the Cub Scout manual. Months later they can use any of these constellations to maintain a constant heading—say, keeping north behind them while flying south, or flying into the celestial pole the next spring to return north.
Honeybees perform a dance that tells their hivemates the direction and distance of a food source with respect to the sun. As if that weren’t impressive enough, the bees have evolved a variety of calibrations and backup systems to deal with the engineering complexities of solar navigation. The dancer uses an internal clock to compensate for the movement of the sun between the time she discovered the source and the time she passes on the information. If it’s cloudy, the other bees estimate the direction using the polarization of light in the sky. These feats are the tip of an iceberg of honeybee ingenuity, documented by Karl Von Frisch, James Gould, and others. A psychologist colleague of mine once thought that bees offered a good pedagogical opportunity to convey the sophistication of neural computation to our undergraduates. He devoted the first week of his entry-level course in cognitive science to some of the ingenious experiments. The next year the lectures spilled over to the second week, then the third, and so on, until the students complained that the course had become an Introduction to Bee Cognition.
There are dozens of comparable examples. Many species compute how much time to forage at each patch so as to optimize their rate of return of calories per energy expended in foraging. Some birds learn the emphemeris function, the path of the sun above the horizon over the course of the day and the year, necessary for navigating by the sun. The barn owl uses sub-millisecond discrepancies between the arrival times of a sound at its two ears to swoop down on a rustling mouse in pitch blackness. Cacheing species place nuts and seeds in unpredictable hiding places to foil thieves, but months later must recall them all. I mentioned in the preceding chapter that the Clark’s Nutcracker can remember ten thousand hiding places. Even Pavlovian and operant conditioning, the textbook cases of learning by association, turn out to be not a general stickiness of coinciding stimuli and responses in the brain, but complex algorithms for multivariate, nonstationary time series analysis (predicting when events will occur, based on their history of occurrences).
The moral of this animal show is that animals’ brains are just as specialized and well engineered as their bodies. A brain is a precision instrument that allows a creature to use information to solve the problems presented by its lifestyle. Since organisms’ lifestyles differ, and since they are related to one another in a great bush, not a great chain, species cannot be ranked in IQ or by the percentage of human intelligence they have achieved. Whatever is special about the human mind cannot be just more, or better, or more flexible animal intelligence, because there is no such thing as generic animal intelligence. Each animal has evolved information-processing machinery to solve its problems, and we evolved machinery to solve ours. The sophisticated algorithms found in even the tiniest dabs of nervous tissue serve as yet another eye-opener—joining the difficulty of building a robot, the circumscribed effects of brain damage, and the similarities between twins reared apart—for the hidden complexity we should expect to find in the human mind.
The brains of mammals, like the bodies of mammals, follow a common general plan. Many of the same cell types, chemicals, tissues, sub-organs, way-stations, and pathways are found throughout the class, and the major visible differences come from inflating or shrinking the parts. But under the microscope, differences appear. The number of cortical areas differs widely, from twenty or fewer in rats to fifty or more in humans. Primates differ from other mammals in the number of visual areas, their interconnections, and their hookup to the motor and decision regions of the frontal lobes. When a species has a noteworthy talent, it is reflected in the gross anatomy of its brain, sometimes in ways visible to the naked eye. The takeover of monkeys’ brains by visual areas (about one-half the territory) reflects—more accurately, allows—their aptitude for depth, color, motion, and visually guided grasping. Bats that rely on sonar have additional brain areas dedicated to their ultrasonic hearing, and desert mice that cache seeds are born with a bigger hippocampus— a seat of the cognitive map—than closely related species that don’t cache.
The human brain, too, tells an evolutionary story. Even a quick side-by-side comparison shows that the primate brain must have been considerably re-engineered to end up as a human brain. Our brains are about three times too big for a generic monkey or ape of our body size. The inflation is accomplished by prolonging fetal brain growth for a year after birth. If our bodies grew proportionally during that period, we would be ten feet tall and weigh half a ton.
The major lobes and patches of the brain have been revamped as well. The olfactory bulbs, which underlie the sense of smell, have shriveled to a third of the expected primate size (already puny by mammalian standards), and the main cortical areas for vision and movement have shrunk proportionally as well. Within the visual system, the first stop for information, the primary visual cortex, takes up a smaller proportion of the whole brain, while the later areas for complex-form processing expand, as do the temporo-parietal areas that shunt visual information to the language and conceptual regions. The areas for hearing, especially for understanding speech, have grown, and the prefrontal lobes, the seat of deliberate thought and planning, have ballooned to twice what a primate our size should have. While the brains of monkeys and apes are subtly asymmetrical, the human brain, especially in the areas devoted to language, is so lopsided that the two hemispheres can be distinguished by shape in the jar. And there have been takeovers of primate brain areas for new functions. Broca’s area, involved in speech, has a homologue (evolutionary counterpart) in monkeys, but they obviously don’t use it for speech, and they don’t even seem to use it to produce shrieks, barks, and other calls.
It’s interesting to find these differences, but the human brain could be radically different from an ape’s brain even if one looked like a perfect scale model of the other. The real action is in the patterns of connections among neurons, just as the differences in content among different computer programs, microchips, books, or videocassettes lie not in their gross shapes but in the combinatorial arrangements of their tiny constituents. Virtually nothing is known about the functioning microcir-cuitry of the human brain, because there is a shortage of volunteers willing to give up their brains to science before they are dead. If we could somehow read the code in the neural circuitry of growing humans and apes, we would surely find substantial differences.
Are the marvelous algorithms of animals mere “instincts” that we have lost or risen above? Humans are often said to have no instincts beyond the vegetative functions; we are said to reason and behave flexibly, freed from specialized machinery. The featherless biped surely understands astronomy in a sense that the feathered biped does not! True enough, but it is not because we have fewer instincts than other animals; it is because we have more. Our vaunted flexibility comes from scores of instincts assembled into programs and pitted in competitions. Darwin called human language, the epitome of flexible behavior, “an instinct to acquire an art” (giving me the title for The Language Instinct), and his follower William James pressed the point:
Now, why do the various animals do what seem to us such strange things, in the presence of such outlandish stimuli? Why does the hen, for example, submit herself to the tedium of incubating such a fearfully uninteresting set of objects as a nestful of eggs, unless she have some sort of a prophetic inkling of the result? The only answer is ad hominem. We can only interpret the instincts of brutes by what we know of instincts in ourselves. Why do men always lie down, when they can, on soft beds rather than on hard floors? Why do they sit round the stove on a cold day? Why, in a room, do they place themselves, ninety-nine times out of a hundred, with their faces towards its middle rather than to the wall? Why do they prefer saddle of mutton and champagne to hard-tack and pond-water? Why does the maiden interest the youth so that everything about her seems more important and significant than anything else in the world? Nothing more can be said than that these are human ways, and that every creature likes its own ways, and takes to the following them as a matter of course. Science may come and consider these ways, and find that most of them are useful. But it is not for the sake of their utility that they are followed, but because at the moment of following them we feel that that is the only appropriate and natural thing to do. Not one man in a billion, when taking his dinner, ever thinks of utility. He eats because the food tastes good and makes him want more. If you ask him why he should want to eat more of what tastes like that, instead of revering you as a philosopher we will probably laugh at you for a fool.…
And so, probably, does each animal feel about the particular things it tends to do in presence of particular objects. To the broody hen the notion would probably seem monstrous that there should be a creature in the world to whom a nestful of eggs was not the utterly fascinating and precious and never-to-be-too-much sat-upon object which it is to her.
The human reactions described in the passage still may strike you as versions of animal instincts. What about our rational, flexible thought? Can it be explained as a set of instincts? In the preceding chapter I showed how our precision intelligence can be broken down into smaller and smaller agents or networks of information processing. At the lowest levels, the steps have to be as automatic and unanalyzed as the reactions of the most brutish animal. Remember what the tortoise said to Achilles. No rational creature can consult rules all the way down; that way infinite regress lies. At some point a thinker must execute a rule, because he just can’t help it: it’s the human way, a matter of course, the only appropriate and natural thing to do—in short, an instinct. When all goes well, our reasoning instincts link up into complex programs for rational analysis, but that is not because we somehow commune with a realm of truth and reason. The same instincts can be seduced by sophistry, bump up against paradoxes like Zeno’s beguiling demonstrations that motion is impossible, or make us dizzy as they ponder mysteries like sentience and free will. Just as an ethologist unmasks an animal’s instincts with clever manipulations of its world, such as slipping a mechanical bee into a hive or rearing a chick in a planetarium, psychologists can unmask human reasoning instincts by couching problems in devilish ways, as we shall see in Chapter 5.
Ambrose Bierce’s Devil’s Dictionary defines our species as follows:
Man, n. An animal so lost in rapturous contemplation of what he thinks he is as to overlook what he indubitably ought to be. His chief occupation is extermination of other animals and his own species, which, however, multiplies with such insistent rapidity as to infest the whole habitable earth and Canada.
Homo sapiens sapiens is indeed an unprecedented animal, with many zoologically unique or extreme traits. Humans achieve their goals by complex chains of behavior, assembled on the spot and tailored to the situation. They plan the behavior using cognitive models of the causal structure of the world. They learn these models in their lifetimes and communicate them through language, which allows the knowledge to accumulate within a group and over generations. They manufacture and depend upon many kinds of tools. They exchange goods and favors over long periods of time. Food is transported long distances, processed extensively, stored, and shared. Labor is divided between the sexes. Humans form large, structured coalitions, especially among males, and coalitions wage war against each other. Humans use fire. Kinship systems are complex and vary with other aspects of their lifestyles. Mating relations are negotiated by kin, often by groups exchanging daughters. Ovulation is concealed, and females may choose to have sex at any time rather than at certain points in a reproductive cycle.
A few of these traits are found among some of the great apes, but to a much lesser degree, and most are not found at all. And humans have rediscovered traits that are rare among primates but found in other animals. They are bipedal. They live longer than other apes, and bear helpless offspring who stay children (that is, sexually immature) for a substantial part of their lives. Hunting is important, and meat a large part of the diet. Males invest in their offspring: they tote children around, protect them against animals and other humans, and give them food. And as The Devil’s Dictionary points out, humans occupy every ecozone on earth.
Aside from the retooling of the skeleton that gives us upright posture and precision manipulation, what makes us unusual is not our body but our behavior and the mental programs that organize it. In the comic strip Calvin and Hobbes, Calvin asks his tiger companion why people are never content with what they have. Hobbes replies, “Are you kidding? Your fingernails are a joke, you’ve got no fangs, you can’t see at night, your pink hides are ridiculous, your reflexes are nil, and you don’t even have tails! Of course people aren’t content!” But despite these handicaps, humans control the fate of tigers, rather than vice versa. Human evolution is the original revenge of the nerds.
Perhaps recoiling from the image of the pasty-faced, pocket-protected, polyester-clad misfits, theorists on human evolution have looked far and wide for alternative theories. Human ingenuity has been explained away as a by-product of blood vessels in the skull that radiate heat, as a runaway courtship device like the peacock’s tail, as a stretching of chimpanzee childhood, and as an escape hatch that saved the species from the evolutionary dead end of bearing fewer and fewer offspring. Even in theories that acknowledge that intelligence itself was selected for, the causes are badly underpowered in comparison with the effects. In various stories the full human mind sprang into existence to solve narrow problems like chipping tools out of stone, cracking open nuts and bones, throwing rocks at animals, keeping track of toddlers, following herds to scavenge their dead, and maintaining social bonds in a large group.
There are grains of truth in these accounts, but they lack the leverage of good reverse-engineering. Natural selection for success in solving a particular problem tends to fashion an idiot savant like the dead-reckoning ants and stargazing birds. We need to know what the more general kinds of intelligence found in our species are good for. That requires a good description of the improbable feats the human mind accomplishes, not just one-word compliments like “flexibility” or “intelligence.” That description must come from the study of the modern mind, cognitive science. And because selection is driven by the fate of the whole individual, it is not enough to explain the evolution of a brain in a vat. A good theory has to connect all the parts of the human lifestyle—all ages, both sexes, anatomy, diet, habitat, and social life. That is, it has to characterize the ecological niche that humans entered.
The only theory that has risen to this challenge comes from John Tooby and the anthropologist Irven DeVore. Tooby and DeVore begin by noting that species evolve at one another’s expense. We fantasize about the land of milk and honey, the big rock candy mountain, and tangerine trees with marmalade skies, but real ecosystems are different. Except for fruits (which trick hungry animals into dispersing seeds), virtually every food is the body part of some other organism, which would just as soon keep that part for itself. Organisms evolve defenses against being eaten, and would-be diners evolve weapons to overcome these defenses, prodding the would-be meals to evolve better defenses, and so on, in an evolutionary arms race. These weapons and defenses are genetically based and relatively fixed within the lifetime of the individual; therefore they change slowly. The balance between eater and eaten develops only over evolutionary time.
Humans, Tooby and DeVore suggest, entered the “cognitive niche.” Remember the definition of intelligence from Chapter 2: using knowledge of how things work to attain goals in the face of obstacles. By learning which manipulations achieve which goals, humans have mastered the art of the surprise attack. They use novel, goal-oriented courses of action to overcome the Maginot Line defenses of other organisms, which can respond only over evolutionary time. The manipulations can be novel because human knowledge is not just couched in concrete instructions like “how to catch a rabbit.” Humans analyze the world using intuitive theories of objects, forces, paths, places, manners, states, substances, hidden biochemical essences, and, for other animals and people, beliefs and desires. (These intuitive theories are the topic of Chapter 5.) People compose new knowledge and plans by mentally playing out combinatorial interactions among these laws in their mind’s eye.
Many theorists have wondered what illiterate foragers do with their capacity for abstract intelligence. The foragers would have better grounds for asking the question about modern couch potatoes. Life for foragers (including our ancestors) is a camping trip that never ends, but without the space blankets, Swiss Army knives, and freeze-dried pasta al pesto. Living by their wits, human groups develop sophisticated technologies and bodies of folk science. All human cultures ever documented have words for the elements of space, time, motion, speed, mental states, tools, flora, fauna, and weather, and logical connectives (not, and, same, opposite, part-whole, and general-particular). They combine the words into grammatical sentences and use the underlying propositions to reason about invisible entities like diseases, meteorological forces, and absent animals. Mental maps represent the locations of thousands of noteworthy sites, and mental calendars represent nested cycles of weather, animal migrations, and the life histories of plants. The anthropologist Louis Liebenberg recounts a typical experience with the !Xõ of the central Kalahari Desert:
While tracking down a solitary wildebeest spoor [tracks] of the previous evening !Xõ trackers pointed out evidence of trampling which indicated that the animal had slept at that spot. They explained consequently that the spoor leaving the sleeping place had been made early that morning and was therefore relatively fresh. The spoor then followed a straight course, indicating that the animal was on its way to a specific destination. After a while, one tracker started to investigate several sets of footprints in a particular area. He pointed out that these footprints all belonged to the same animal, but were made during the previous days. He explained that the particular area was the feeding ground of that particular wildebeest. Since it was, by that time, about mid-day, it could be expected that the wildebeest may be resting in the shade in the near vicinity.
All foraging peoples manufacture cutters, pounders, containers, cordage, nets, baskets, levers, and spears and other weapons. They use fire, shelters, and medicinal drugs. Their engineering is often ingenious, exploiting poisons, smokeouts, glue traps, gill nets, baited lines, snares, corrals, weirs, camouflaged pits and clifftops, blowguns, bows and arrows, and kites trailing sticky fishing lines made out of spider silk.
The reward is an ability to crack the safes of many other living things: burrowing animals, plants’ underground storage organs, nuts, seeds, bone marrow, tough-skinned animals and plants, birds, fish, shellfish, turtles, poisonous plants (detoxified by peeling, cooking, soaking, parboiling, fermenting, leaching, and other tricks of the kitchen magician), quick animals (which can be ambushed), and large animals (which cooperating groups can drive, exhaust, surround, and dispatch with weapons). Ogden Nash wrote:
The hunter crouches in his blind
’Neath camouflage of every kind,
And conjures up a quacking noise
To lend allure to his decoys.
This grown-up man, with pluck and luck
Is hoping to outwit a duck.
And outwit it he does. Humans have the unfair advantage of attacking in this lifetime organisms that can beef up their defenses only in subsequent ones. Many species cannot evolve defenses rapidly enough, even over evolutionary time, to defend themselves against humans. That is why species drop like flies whenever humans first enter an ecosystem. And it’s not just the snail darters and snowy owls recently threatened by dams and loggers. The reason you have never seen a living mastodon, saber-tooth, giant woolly rhinoceros, or other fantastic Ice Age animal is that humans apparently extinguished them thousands of years ago.
The cognitive niche embraces many of the zoologically unusual features of our species. Tool manufacture and use is the application of knowledge about causes and effects among objects in the effort to bring about goals. Language is a means of exchanging knowledge. It multiplies the benefit of knowledge, which can not only be used but exchanged for other resources, and lowers its cost, because knowledge can be acquired from the hard-won wisdom, strokes of genius, and trial and error of others rather than only from risky exploration and experimentation. Information can be shared at a negligible cost: if I give you a fish, I no longer possess the fish, but if I give you information on how to fish, I still possess the information myself. So an information-exploiting lifestyle goes well with living in groups and pooling expertise—that is, with culture. Cultures differ from one another because they pool bodies of expertise fashioned in different times and places. A prolonged childhood is an apprenticeship for knowledge and skills. That shifts the balance of payoffs for males toward investing time and resources in their offspring and away from competing over sexual access to females (see Chapter 7). And that in turn makes kinship a concern of both sexes and all ages. Human lives are long to repay the investment of a long apprenticeship. New habitats can be colonized because even if their local conditions differ, they obey the laws of physics and biology that are already within humans’ ken, and can be exploited and outsmarted in their turn.
Why did some miocene ape first enter the cognitive niche? Why not a groundhog, or a catfish, or a tapeworm? It only happened once, so no one knows. But I would guess that our ancestors had four traits that made it especially easy and worth their while to evolve better powers of causal reasoning.
First, primates are visual animals. In monkeys such as the rhesus macaque, half the brain is dedicated to sight. Stereoscopic vision, the use of differences in the vantage points of the two eyes to give a sense of depth, developed early in the primate lineage, allowing early nocturnal primates to move among treacherous fine branches and to grab insects with their hands. Color vision accompanied the switch of the ancestors of monkeys and apes to the day shift and their new taste for fruits, which advertise their ripeness with gaudy hues.
Why would the vision thing make such a difference? Depth perception defines a three-dimensional space filled with movable solid objects. Color makes objects pop out from their backgrounds, and gives us a sensation that corresponds to the stuff an object is made of, distinct from our perception of the shape of the stuff. Together they have pushed the primate brain into splitting the flow of visual information into two streams: a “what” system, for objects and their shapes and compositions, and a “where” system, for their locations and motions. It can’t be a coincidence that the human mind grasps the world—even the most abstract, ethereal concepts—as a space filled with movable things and stuff (see Chapters 4 and 5). We say that John went from being sick to being well, even if he didn’t move an inch; he could have been in bed the whole time. Mary can give him many pieces of advice, even if they merely talked on the phone and nothing changed hands. Even scientists, when they try to grasp abstract mathematical relationships, plot them in graphs that show them as two- and three-dimensional shapes. Our capacity for abstract thought has co-opted the coordinate system and inventory of objects made available by a well-developed visual system.
It is harder to see how a standard mammal could have moved in that direction. Most mammals hug the ground sniffing the rich chemical tracks and trails left behind by other living things. Anyone who has walked a frisky cocker spaniel as it explores the invisible phantasmagoria on a sidewalk knows that it lives in an olfactory world beyond our understanding. Here is an exaggerated way of stating the difference. Rather than living in a three-dimensional coordinate space hung with movable objects, standard mammals live in a two-dimensional flatland which they explore through a zero-dimensional peephole. Edwin Abbott’s Flatland, a mathematical novel about the denizens of a plane, showed that a two-dimensional world differs from our own in ways other than just lacking one third of the usual dimensions. Many geometric arrangements are simply impossible. A full-faced human figure has no way of getting food into his mouth, and a profiled one would be divided into two pieces by his digestive tract. Simple devices like tubes, knots, and wheels with axles are unbuildable. If most mammals think in a cognitive flatland, they would lack the mental models of movable solid objects in 3-D spatial and mechanical relationships that became so essential to our mental life.
A second possible prerequisite, this one found in the common ancestor of humans, chimpanzees, and gorillas, is group living. Most apes and monkeys are gregarious, though most mammals are not. Living together has advantages. A cluster of animals is not much more detectable to a predator than a single animal, and if it is detected, the likelihood that any individual will be picked off is diluted. (Drivers feel less vulnerable speeding when they are in a group of speeders, because chances are the traffic cop will stop someone else.) There are more eyes, ears, and noses to detect a predator, and the attacker can sometimes be mobbed. A second advantage is in foraging efficiency. The advantage is most obvious in cooperative hunting of large animals, such as in wolves and lions, but it also helps in sharing and defending other ephemeral food resources too big to be consumed by the individual who found it, such as a tree laden with ripe fruit. Primates that depend on fruit, and primates that spend time on the ground (where they are more vulnerable to predators), tend to hang out in groups.
Group living could have set the stage for the evolution of humanlike intelligence in two ways. With a group already in place, the value of having better information is multiplied, because information is the one commodity that can be given away and kept at the same time. Therefore a smarter animal living in a group enjoys a double advantage: the benefit of the knowledge and the benefit of whatever it can get in trade for the knowledge.
The other way in which a group can be a crucible of intelligence is that group living itself poses new cognitive challenges. There are also disadvantages to the madding crowd. Neighbors compete over food, water, mates, and nest sites. And there is the risk of exploitation. Hell is other people, said Jean-Paul Sartre, and if baboons were philosophers no doubt they would say that hell is other baboons. Social animals risk theft, cannibalism, cuckoldry, infanticide, extortion, and other treachery.
Every social creature is poised between milking the benefits and suffering the costs of group living. That creates a pressure to stay on the right side of the ledger by becoming smarter. In many kinds of animals, the largest-brained and smartest-behaving species are social: bees, parrots, dolphins, elephants, wolves, sea lions, and, of course, monkeys, gorillas, and chimpanzees. (The orangutan, smart but almost solitary, is a puzzling exception.) Social animals send and receive signals to coordinate predation, defense, foraging, and collective sexual access. They exchange favors, repay and enforce debts, punish cheaters, and join coalitions.
The collective expression for hominoids, “a shrewdness of apes,” tells a story. Primates are sneaky baldfaced liars. They hide from rivals’ eyes to flirt, cry wolf to attract or divert attention, even manipulate their lips into a poker face. Chimpanzees monitor one another’s goals, at least crudely, and sometimes appear to use them in pedagogy and deception. One chimp, shown a set of boxes with food and one with a snake, led his companions to the snake, and after they fled screaming, feasted in peace. Vervet monkeys are yentas who keep close track of everyone’s comings and goings and friends and enemies. But they are so dense about the nonsocial world that they ignore the tracks of a python and the ominous sight of a carcass in a tree, the unique handiwork of a leopard.
Several theorists have proposed that the human brain is the outcome of a cognitive arms race set in motion by the Machiavellian intelligence of our primate forebears. There’s only so much brain power you need to subdue a plant or a rock, the argument goes, but the other guy is about as smart as you are and may use that intelligence against your interests. You had better think about what he is thinking about what you are thinking he is thinking. As far as brain power goes, there’s no end to keeping up with the Joneses.
My own guess is that a cognitive arms race by itself was not enough to launch human intelligence. Any social species can begin a never-ending escalation of brain power, but none except ours has, probably because without some other change in lifestyle, the costs of intelligence (brain size, extended childhood, and so on) would damp the positive feedback loop. Humans are exceptional in mechanical and biological, not just social, intelligence. In a species that runs on information, each faculty multiplies the value of the others. (Incidentally, the expansion of the human brain is no evolutionary freak crying out for a runaway positive feedback loop. The brain tripled in size in five million years, but that is leisurely by evolutionary timekeeping. There was enough time in hominid evolution for the brain to shoot up to human size, shrink back down, and shoot up again several times over.)
A third pilot of intelligence, alongside good vision and big groups, is the hand. Primates evolved in trees and have hands to grasp the branches. Monkeys use all four limbs to run along the tops of branches, but apes hang from them, mainly by their arms. They have put their well-developed hands to use in manipulating objects. Gorillas meticulously dissect tough or thorny plants to pick out the edible matter, and chimpanzees use simple tools such as stems to fish out termites, rocks to bash open nuts, and mashed leaves to sponge up water. As Samuel Johnson said about dogs walking on their hind legs, while it is not done well, you are surprised to find it done at all. Hands are levers of influence on the world that make intelligence worth having. Precision hands and precision intelligence co-evolved in the human lineage, and the fossil record shows that hands led the way.
Finely tooled hands are useless if you have to walk on them all the time, and they could not have evolved by themselves. Every bone in our bodies has been reshaped to give us our upright posture, which frees the hands for carrying and manipulating. Once again we have our ape ancestors to thank. Hanging from trees calls for a body plan that is different from the horizontal four-wheel-drive design of most mammals. Apes’ bodies are already tilted upward with arms that differ from their legs, and chimpanzees (and even monkeys) walk upright for short distances to carry food and objects.
Fully upright posture may have evolved under several selection pressures. Bipedal walking is a biomechanically efficient way to retool a tree-hanging body to cover distance on the flat ground of the newly entered savanna. Upright posture also allows one to peer over grass like a marmot. Hominids go out in the midday sun; this zoologically unusual work shift brought in several human adaptations for keeping cool, such as hairlessness and profuse sweating. Upright posture might be another; it is the opposite of lying down to get a tan. But carrying and manipulation must have been crucial inducements. With the hands free, tools could be assembled out of materials from different locations and brought to where they were most useful, and food and children could be carried to safe or productive areas.
A final usher of intelligence was hunting. Hunting, tool use, and bipedalism were for Darwin the special trinity that powered human evolution. “Man the Hunter” was the major archetype in both serious and pop accounts through the 1960s. But the macho image that resonated with the decade of John Glenn and James Bond lost its appeal in the feminist-influenced small planet of the 1970s. A major problem for Man the Hunter was that it credited the growth of intelligence to the teamwork and foresight needed by men in groups to fell large game. But natural selection sums over the lives of both sexes. Women did not wait in the kitchen to cook the mastodon that Dad brought home, nor did they forgo the expansion of intelligence enjoyed by evolving men. The ecology of modern foraging peoples suggests that Woman the Gatherer provided a substantial portion of the calories in the form of highly processed plant foods, and that requires mechanical and biological acumen. And, of course, in a group-living species, social intelligence is as important a weapon as spears and clubs.
But Tooby and DeVore have argued that hunting was nonetheless a major force in human evolution. The key is to ask not what the mind can do for hunting, but what hunting can do for the mind. Hunting provides sporadic packages of concentrated nutrients. We did not always have tofu, and the best natural material for building animal flesh is animal flesh. Though plant foods supply calories and other nutrients, meat is a complete protein containing all twenty amino acids, and provides energy-rich fat and indispensable fatty acids. Across the mammals, carnivores have larger brains for their body size than herbivores, partly because of the greater skill it takes to subdue a rabbit than to subdue grass, and partly because meat can better feed ravenous brain tissue. Even in the most conservative estimates, meat makes up a far greater proportion of foraging humans’ diet than of any other primate’s. That may have been one of the reasons we could afford our expensive brains.
Chimpanzees collectively hunt small animals like monkeys and bush pigs, so our common ancestor probably hunted as well. The move to the savanna must have made hunting more appealing. Notwithstanding the teeming wildlife in the Save-the-Rainforest posters, real forests have few large animals. Only so much solar energy falls on a patch of ground, and if the biomass it supports is locked up in wood it is not available to make animals. But grass is like the legendary self-replenishing goblet, growing back as soon as it is grazed. Grasslands can feed vast herds of herbivores, who in turn feed carnivores. Evidence of butchery appears in the fossil record almost two million years ago, the time of Homo habilis. Hunting must be even older, since we know that chimpanzees do it, and their activities would not leave evidence in the fossil record. Once our ancestors increased their hunting, the world opened up. Plant foods are scarce during the winter at higher altitudes and latitudes, but hunters can survive there. There are no vegetarian Eskimos.
Our ancestors have sometimes been characterized as meek scavengers rather than brave hunters, in keeping with today’s machismo-puncturing ethos. But while hominids may occasionally have scavenged, they probably could not have made a living from it, and if they did, they were no wimps. Vultures get away with scavenging because they can scan large territories for carcasses and flee on short notice when more formidable competitors show up. Otherwise, scavenging is not for the faint of heart. A carcass is jealously guarded by its hunter or an animal fierce enough to have stolen it. It is attractive to microorganisms, who quickly poison the meat to repel other would-be scavengers. So when modern primates or hunter-gatherers come across a carcass, they usually leave it alone. In a poster widely available in head shops in the early 1970s, one vulture says to the other, “Patience, my ass! I’m going to kill something.” The poster got it right, except for the vulture part: mammals that do scavenge, such as hyenas, also hunt.
Meat is also a major currency of our social life. Imagine a cow who tries to win the favors of a neighbor by dropping a clump of grass at its feet. One could forgive the second cow for thinking, “Thanks, but I can get my own grass.” The nutritional jackpot of a felled animal is another matter. Miss Piggy once advised, “Never eat anything bigger than you can lift.” A hunter with a dead animal larger than he can eat and about to become a putrefying mass is faced with a unique opportunity. Hunting is largely a matter of luck. In the absence of refrigeration, a good place to store meat for leaner times is in the bodies of other hunters who will return the favor when fortunes reverse. This eases the way for the male coalitions and the extensive reciprocity that are ubiquitous in foraging societies.
And there are other markets for a hunter’s surplus. Having concentrated food to offer one’s offspring changes the relative payoffs for males between investing in their young and competing with other males for access to females. The robin bringing a worm to the nestlings reminds us that most animals that provision their young do so with prey, the only food that repays the effort to obtain it and transport it.
Meat also figures into sexual politics. In all foraging societies, presumably including our ancestors’, hunting is overwhelmingly a male activity. Women are encumbered with children, which makes hunting inconvenient, and men are bigger and more adept at killing because of their evolutionary history of killing each other. As a result, males can invest surplus meat in their children by provisioning the children’s pregnant or nursing mothers. They also can trade meat with females for plant foods or for sex. Brazen bartering of the carnal for the carnal has been observed in baboons and chimpanzees and is common in foraging peoples. Though people in modern societies are ever-so-more discreet, an exchange of resources for sexual access is still an important part of the interactions between men and women all over the world. (Chapter 7 explores these dynamics and how they originated in differences in reproductive anatomy, though of course anatomy is not destiny in modern ways of life.) In any case, we have not lost the association completely. Miss Manners’ Guide to Excruciatingly Correct Behavior advises:
There are three possible parts to a date, of which at least two must be offered: entertainment, food, and affection. It is customary to begin a series of dates with a great deal of entertainment, a moderate amount of food, and the merest suggestion of affection. As the amount of affection increases, the entertainment can be reduced proportionately. When the affection is the entertainment, we no longer call it dating. Under no circumstances can the food be omitted.
Of course no one really knows whether these four habits formed the base camp for the ascent of human intelligence. And no one knows whether there are other, untried gradients to intelligence in biological design space. But if these traits do explain why our ancestors were the only species out of fifty million to follow that route, it would have sobering implications for the search for extraterrestrial intelligence. A planet with life may not be enough of a launching pad. Its history might have to include a nocturnal predator (to get stereo vision), with descendants that switched to a diurnal lifestyle (for color) in which they depended on fruit and were vulnerable to predators (for group living), which then changed their means of locomotion to swinging beneath branches (for hands and for precursors to upright posture), before a climate shift sent them from the forest into grasslands (for upright posture and hunting). What is the probability that a given planet, even a planet with life, has such a history?
Species |
Date |
Height |
Physique |
Brain |
Chimp-hominid ancestor (if similar to modern chimps) |
8–6 million years ago |
1–1.7 meters |
long arms, short thumbs, curved fingers and toes; adapted for knuckle-walking and tree-climbing |
450 cc |
Ardipithecus ramidus |
4.4 million years ago |
? |
probably bipedal |
? |
Australopithecus anamensis |
4.2–3.9 million years ago |
? |
bipedal |
? |
Australopithecus afarensis (Lucy) |
4–2.5 million years ago |
1–1.2 meters |
fully bipedal with modified hands but ape-like features: thorax, long arms, curved fingers and toes |
400–500 cc |
Homo habilis (Handyman) |
2.3–1.6 million years ago |
1–1.5 meters |
some specimens: small with long arms; others: robust but human |
500–800 cc |
Homo erectus |
1.9 million–300,000 (maybe 27,000) years ago |
1.3–1.5 meters |
robust but human |
750–1250 cc |
Archaic Homo sapiens |
400,000–100,000 years ago |
? |
robust but modern |
1100–1400 cc |
Early Homo sapiens |
130,000–60,000 years ago |
1.6–1.85 meters |
robust but modern |
1200–1700 cc |
Homo sapiens (Cro-Magnon) |
45,000–12,000 years ago |
1.6–1.8 meters |
modern |
1300–1600 (cf. today: 1000–2000, average 1350) |
The dry bones of the fossil record tell of a gradual entry into the cognitive niche. A summary of the current evidence on the species thought to be our direct ancestors is shown in the table below.
Skull |
Teeth |
Tools |
Distribution |
very low forehead; projecting face; huge brow ridges |
large canines |
stone hammers, leaf sponges, stem probes, branch levers |
West Africa |
? |
chimplike molars but not canines |
? |
East Africa |
apelike fragments |
chimplike size and placement; humanlike enamel |
? |
East Africa |
low flat forehead; projecting face; big brow ridges |
large canines and molars |
none? flakes? |
East Africa (maybe also west) |
smaller face; rounder skull |
smaller molars |
flakes, choppers, scrapers |
East and South Africa |
thick; large brow ridges (Asia); smaller, protruding face |
smaller teeth |
symmetrical hand axes |
Africa (may be separate species), Asia, Europe |
higher skull; smaller, protruding face; large brow ridges |
smaller teeth |
better hand axes, retouched flakes |
Africa, Asia, Europe |
high skull; medium brow ridges; slightly protruding face; chin |
smaller teeth |
retouched flakes; flake-blades; points |
Africa, Western Asia |
modern |
modern |
blades; drills; spear throwers; needles; engravers; bone |
worldwide |
Millions of years before our brains billowed out, some descendants of the common ancestor of chimpanzees and humans walked upright. In the 1920s that discovery came as a shock to human chauvinists who imagined that our glorious brains led us up the ladder, perhaps as our ancestors decided at each rung what use to make of their newfound smarts. But natural selection could not have worked that way. Why bulk up your brain if you can’t put it to use? The history of paleoanthropology is the discovery of earlier and earlier birthdays for upright posture. The most recent discoveries put it at four or even four and a half million years ago. With hands freed, subsequent species ratchet upward, click by click, in the features that distinguish us: the dexterity of hands, the sophistication of tools, the dependence on hunting, the size of brains, the range of habitats. The teeth and jaw become smaller. The face that opposes it becomes less muzzle-like. The brow ridges that anchor the muscles that close the jaw shrink and disappear. Our delicate faces differ from the brutes’ because tools and technology have taken over from teeth. We slaughter and skin animals with blades, and soften plants and meats with fire. That eases the mechanical demands on the jaw and skull, allowing us to shave bone from our already heavy heads. The sexes come to differ less in size, suggesting that males spent less of their resources beating each other up and perhaps more on their children and the children’s mothers.
The stepwise growth of the brain, propelled by hands and feet and manifested in tools, butchered bones, and increased range, is good evidence, if evidence were needed, that intelligence is a product of natural selection for exploitation of the cognitive niche. The package was not an inexorable unfolding of hominid potential. Other species, omitted from the table, spun off in every epoch to occupy slightly different niches: nutcracking and root-gnawing australopithecines, perhaps one of the two habiline subtypes, quite possibly the Asian branches of erectus and archaic sapiens, and probably the Ice Age-adapted Neanderthals. Each species might have been outcompeted when a neighboring, more sapiens-like population had entered far enough into the cognitive niche to duplicate the species’ more specialized feats and do much else besides. The package was also not the gift of a macromutation or random drift—for how could such luck have held up in one lineage for millions of years, over hundreds of thousands of generations, in species after bigger-brained species? Moreover, the bigger brains were no mere ornaments but allowed their owners to make finer tools and infest more of the planet.
According to the standard timetable in paleoanthropology, the human brain evolved to its modern form in a window that began with the appearance of Homo habilis two million years ago and ended with the appearance of “anatomically modern humans,” Homo sapiens sapiens, between 200,000 and 100,000 years ago. I suspect that our ancestors were penetrating the cognitive niche for far longer than that. Both ends of the R&D process might have to be stretched beyond the textbook dates, providing even more time for our fantastic mental adaptations to have evolved.
At one end of the timetable is the four-million-year-old australop-ithecines-like afarensis (the species of the charismatic fossil called Lucy). They are often described as chimpanzees with upright posture because their brain size was in the chimpanzee ballpark and they left no clear evidence of tool use. That implies that cognitive evolution did not begin till two million years later, when larger-brained habilines earned their “handyman” name by chipping choppers.
But that can’t be right. First, it is ecologically improbable that a tree-dweller could have moved onto open ground and retooled its anatomy for upright walking without repercussions on every other aspect of its lifestyle and behavior. Modern chimps use tools and transport objects, and would have had much more incentive and success if they could carry them around freely. Second, though australopithecines’ hands retain some apelike curvature of the fingers (and may have been used at times to run up trees for safety), the hands visibly evolved for manipulation. Compared to chimps’ hands, their thumbs are longer and more opposable to the other fingers, and their index and middle fingers are angled to allow cupping the palm to grasp a hammerstone or a ball. Third, it’s not so clear that they had a chimp-sized brain, or that they lacked tools. The paleoanthropologist Yves Coppens argues that their brains are thirty to forty percent bigger than expected for a chimpanzee of their body size, and that they left behind modified quartz flakes and other tools. Fourth, skeletons of the tool-using habilines (handymen) have now been found, and they do not look so different from the australopithecines’.
Most important, hominids did not arrange their lives around the convenience of anthropologists. We are lucky that a rock can be carved into a cutter and that it lasts for millions of years, so some of our ancestors inadvertently left us time capsules. But it’s much harder to carve a rock into a basket, a baby sling, a boomerang, or a bow and arrow. Contemporary hunter-gatherers use many self-composting implements for every lasting one, and that must have been true of hominids at every stage. The archeological record is bound to underestimate tool use.
So the standard timetable for human brain evolution begins the story too late; I think it also ends the story too early. Modern humans (us) are said to have first arisen between 200,000 and 100,000 years ago in Africa. One kind of evidence is that the mitochondrial DNA (mDNA) of everyone on the planet (which is inherited only from one’s mother) can be traced back to an African woman living sometime in that period. (The claim is controversial, but the evidence is growing.) Another is that anatomically modern fossils first appear in Africa more than 100,000 years ago and in the Middle East shortly afterward, around 90,000 years ago. The assumption is that human biological evolution had pretty much stopped then. This leaves an anomaly in the timeline. The anatomically modern early humans had the same toolkit and lifestyle as their doomed Neanderthal neighbors. The most dramatic change in the archeological record, the Upper Paleolithic transition—also called the Great Leap Forward and the Human Revolution—had to wait another 50,000 years. Therefore, it is said, the human revolution must have been a cultural change.
Calling it a revolution is no exaggeration. All other hominids come out of the comic strip B.C., but the Upper Paleolithic people were the Flint-stones. More than 45,000 years ago they somehow crossed sixty miles of open ocean to reach Australia, where they left behind hearths, cave paintings, the world’s first polished tools, and today’s aborigines. Europe (home of the Cro-Magnons) and the Middle East also saw unprecedented arts and technologies, which used new materials like antler, ivory, and bone as well as stone, sometimes transported hundreds of miles. The toolkit included fine blades, needles, awls, many kinds of axes and scrapers, spear points, spear throwers, bows and arrows, fishhooks, engravers, flutes, maybe even calendars. They built shelters, and they slaughtered large animals by the thousands. They decorated everything in sight—tools, cave walls, their bodies—and carved knick-knacks in the shapes of animals and naked women, which archeologists euphemistically call “fertility symbols.” They were us.
Ways of life certainly can shoot off without any biological change, as in the more recent agricultural, industrial, and information revolutions. That is especially true when populations grow to a point where the insights of thousands of inventors can be pooled. But the first human revolution was not a cascade of changes set off by a few key inventions. Ingenuity itself was the invention, manifested in hundreds of innovations tens of thousands of miles and years apart. I find it hard to believe that the people of 100,000 years ago had the same minds as those of the Upper Paleolithic revolutionaries to come—indeed, the same minds as ours—and sat around for 50,000 years without it dawning on a single one of them that you could carve a tool out of bone, or without a single one feeling the urge to make anything look pretty.
And there is no need to believe it—the 50,000-year gap is an illusion. First, the so-called anatomically modern humans of 100,000 years ago may have been more modern than their Neanderthal contemporaries, but no one would mistake them for contemporary humans. They had brow ridges, protruding faces, and heavily built skeletons outside the contemporary range. Their bodies had to evolve to become us, and their brains surely did as well. The myth that they are completely modern grew out of the habit of treating species labels as if they were real entities. When applied to evolving organisms, they are no more than a convenience. No one wants to invent a new species every time a tooth is found, so intermediate forms tend to get shoehorned into the nearest available category. The reality is that hominids must always have come in dozens or hundreds of variants, scattered across a large network of occasionally interacting subpopulations. The tiny fraction of individuals immortalized as fossils at any point were not necessarily our direct ancestors. The “anatomically modern” fossils are closer to us than to anyone else, but either they had more evolving to do or they were away from the hotbed of change.
Second, the revolution probably began well before the commonly cited watershed of 40,000 years ago. That’s when fancy artifacts begin to appear in European caves, but Europe has always attracted more attention than it deserves, because it has lots of caves and lots of archeologists. France alone has three hundred well-excavated paleolithic sites, including one whose cave paintings were scrubbed off by an overenthusiastic boy scout troop that mistook them for graffiti. The entire continent of Africa has only two dozen. But one, in Zaire, contains finely crafted bone implements including daggers, shafts, and barbed points, together with grindstones brought from miles away and the remains of thousands of catfish, presumably the victims of these instruments. The collection looks postrevolutionary but is dated at 75,000 years ago. One commentator said it was like finding a Pontiac in Leonardo da Vinci’s attic. But as archeologists are starting to explore this continental attic and date its contents, they are finding more and more Pontiacs: fine stone blades, decorated tools, useless but colorful minerals transported hundreds of miles.
Third, the mitochondrial Eve of 200,000 to 100,000 years ago was not a party to any evolutionary event. Contrary to some fantastic misunderstandings, she did not undergo some mutation that left her descendants smarter or more talkative or less brutish. Nor did she mark the end of human evolution. She is merely a mathematical necessity: the most recent common ancestor of all living people along the female-female-female line of great-great-…-great-grandmothers. For all the definition says, Eve could have been a fish.
Eve, of course, turned out to be not a fish but an African hominid. Why would anyone assume that she was a special hominid, or even that she lived in special times? One reason is that she made many other times and places non-special. If twentieth-century Europeans’ and Asians’ mDNA is a variant of 200,000-year-old African mDNA, they must be descendants of an African population at the time. Eve’s contemporary Europeans and Asians left no mDNA in today’s Europeans and Asians, and thus presumably were not their ancestors (at least—and this is a big proviso—not their all-maternal-line ancestors).
But that says nothing about evolution’s having stopped with Eve. We can assume that most evolution was done with by the time the ancestors of the modern races separated and stopped exchanging genes, since today we are birds of a feather. But that did not happen as soon as Eve breathed her last. The diaspora of the races, and the end of significant human evolution, must have occurred much later. Eve is not our most recent common ancestor, only our most recent common ancestor in the all-maternal line. The most recent common ancestor along a mixed-sex line of descendants lived much later. You and a first cousin share an ancestor of just two generations ago, your common grandmother or grandfather. But in looking for a shared all-female-line ancestor (your mother’s mother’s mother, and so on), then except for one kind of cousin (the child of your mother’s sister), there’s almost no limit to how far back you might have to go. So if someone were to guess the degree of relatedness between you and your cousin based on your most recent ancestor, he would say you were closely related. But if he could check only the most recent all-female-line ancestor, he might guess that you are not related at all! Similarly, the birthday of humanity’s most recent common all-female-line ancestor, mitochondrial Eve, overestimates how long ago all of humanity was still interbreeding.
Well after Eve’s day, some geneticists think, our ancestors passed through a population bottleneck. According to their scenario, which is based on the remarkable sameness of genes across modern human populations, around 65,000 years ago our ancestors dwindled to a mere ten thousand people, perhaps because of a global cooling triggered by a volcano in Sumatra. The human race was as endangered as mountain gorillas are today. The population then exploded in Africa and spun off small bands that moved to other corners of the world, possibly mating now and again with other early humans in their path. Many geneticists believe that evolution is especially rapid when scattered populations exchange occasional migrants. Natural selection can quickly adapt each group to local conditions, so one or more can cope with any new challenge that arises, and their handy genes will then be imported by the neighbors. Perhaps this period saw a final flowering in the evolution of the human mind.0
All reconstructions of our evolutionary history are controversial, and the conventional wisdom changes monthly. But I predict that the closing date of our biological evolution will creep later, and the opening date of the archeological revolution will creep earlier, until they coincide. Our minds and our way of life evolved together.
Are we still evolving? Biologically, probably not much. Evolution has no momentum, so we will not turn into the creepy bloat-heads of science fiction. The modern human condition is not conducive to real evolution either. We infest the whole habitable and not-so-habitable earth, migrate at will, and zigzag from lifestyle to lifestyle. This makes us a nebulous, moving target for natural selection. If the species is evolving at all, it is happening too slowly and unpredictably for us to know the direction.
But Victorian hopes spring eternal. If genuine natural selection cannot improve us, maybe a human-made substitute can. The social sciences are filled with claims that new kinds of adaptation and selection have extended the biological kind. But the claims, I think, are misleading.
The first claim is that the world contains a wonderful process called “adaptation” that causes organisms to solve problems. Now, in Darwin’s strict sense, adaptation in the present is caused by selection in the past. Remember how natural selection gives an illusion of teleology: selection may look like it is adapting each organism to its needs in the present, but really it is just favoring the descendants of organisms that were adapted to their own needs in the past. The genes that built the most adaptive bodies and minds among our ancestors got passed down to build the innate bodies and minds of today (including innate abilities to track certain kinds of environmental variation, as in tanning, callusing, and learning).
But for some, that does not go far enough; adaptation happens daily. “Darwinian social scientists” such as Paul Turke and Laura Betzig believe that “modern Darwinian theory predicts that human behavior will be adaptive, that is, designed to promote maximum reproductive success … through available descendent and nondescendent relatives.” “Functionalists” such as the psychologists Elizabeth Bates and Brian MacWhinney say that they “view the selectional processes operating during evolution and the selectional processes operating during [learning] as part of one seamless natural fabric.” The implication is that there is no need for specialized mental machinery: if adaptation simply makes organisms do the right thing, who could ask for anything more? The optimal solution to a problem—eating with one’s hands, finding the right mate, inventing tools, using grammatical language—is simply inevitable.
The problem with functionalism is that it is Lamarckian. Not in the sense of Lamarck’s second principle, the inheritance of acquired characteristics—the giraffes who stretched their necks and begat baby giraffes with necks pre-stretched. Everyone knows to stay away from that. (Well, almost everyone: Freud and Piaget stuck to it long after it was abandoned by biologists.) It is Lamarckian in the sense of his first principle, “felt need”—the giraffes growing their necks when they hungrily eyed the leaves just out of reach. As Lamarck put it, “New needs which establish a necessity for some part really bring about the existence of that part as a result of efforts.” If only it were so! As the saying goes, if wishes were horses, beggars would ride. There are no guardian angels seeing to it that every need is met. They are met only when mutations appear that are capable of building an organ that meets the need, when the organism finds itself in an environment in which meeting the need translates into more surviving babies, and in which that selection pressure persists over thousands of generations. Otherwise, the need goes unmet. Swimmers do not grow webbed fingers; Eskimos do not grow fur. I have studied three-dimensional mirror-images for twenty years, and though I know mathematically that you can convert a left shoe into a right shoe by turning it around in the fourth dimension, I have been unable to grow a 4-D mental space in which to visualize the flip.
Felt need is an alluring idea. Needs really do feel like they bring forth their own solutions. You’re hungry, you have hands, the food’s in front of you, you eat with your hands; how else could it be? Ah, but you’re the last one we should ask. Your brain was fashioned by natural selection so that it would find such problems obvious. Change the mind (to a robot’s, or to another animal’s, or to a neurological patient’s), or change the problem, and it’s no longer so obvious what’s obvious. Rats can’t learn to drop a piece of food for a larger reward. When chimpanzees try to imitate someone raking in an inaccessible snack, they don’t notice that the rake has to be held business-end down, even if the role model makes a conspicuous show of aligning it properly. Lest you feel smug, the chapters to come will show how the design of our own minds gives rise to paradoxes, brain-teasers, myopias, illusions, irrationalities, and self-defeating strategies that prevent, rather than guarantee, the meeting of our everyday needs.
But what about the Darwinian imperative to survive and reproduce? As far as day-to-day behavior is concerned, there is no such imperative. People watch pornography when they could be seeking a mate, forgo food to buy heroin, sell their blood to buy movie tickets (in India), postpone childbearing to climb the corporate ladder, and eat themselves into an early grave. Human vice is proof that biological adaptation is, speaking literally, a thing of the past. Our minds are adapted to the small foraging bands in which our family spent ninety-nine percent of its existence, not to the topsy-turvy contingencies we have created since the agricultural and industrial revolutions. Before there was photography, it was adaptive to receive visual images of attractive members of the opposite sex, because those images arose only from light reflecting off fertile bodies. Before opiates came in syringes, they were synthesized in the brain as natural analgesics. Before there were movies, it was adaptive to witness people’s emotional struggles, because the only struggles you could witness were among people you had to psych out every day. Before there was contraception, children were unpostponable, and status and wealth could be converted into more children and healthier ones. Before there was a sugar bowl, salt shaker, and butter dish on every table, and when lean years were never far away, one could never get too much sweet, salty, and fatty food. People do not divine what is adaptive for them or their genes; their genes give them thoughts and feelings that were adaptive in the environment in which the genes were selected.
The other extension of adaptation is the seemingly innocuous cliché that “cultural evolution has taken over from biological evolution.” For millions of years, genes were transmitted from body to body and were selected to confer adaptations on organisms. But after humans emerged, units of culture were transmitted from mind to mind and were selected to confer adaptations on cultures. The torch of progress has been passed to a swifter runner. In 2001: A Space Odyssey, a hairy arm hurls a bone into the air, and it fades into a space station.
The premise of cultural evolution is that there is a single phenomenon—the march of progress, the ascent of man, apes to Armageddon— that Darwin explained only up to a point. My own view is that human brains evolved by one set of laws, those of natural selection and genetics, and now interact with one another according to other sets of laws, those of cognitive and social psychology, human ecology, and history. The reshaping of the skull and the rise and fall of empires may have little in common.
Richard Dawkins has drawn the clearest analogy between the selection of genes and the selection of bits of culture, which he dubbed memes. Memes such as tunes, ideas, and stories spread from brain to brain and sometimes mutate in the transmission. New features of a meme that make its recipients more likely to retain and disseminate it, such as being catchy, seductive, funny, or irrefutable, will lead to the meme’s becoming more common in the meme pool. In subsequent rounds of retelling, the most spreadworthy memes will spread the most and will eventually take over the population. Ideas will therefore evolve to become better adapted to spreading themselves. Note that we are talking about ideas evolving to become more spreadable, not people evolving to become more knowledgeable.
Dawkins himself used the analogy to illustrate how natural selection pertains to anything that can replicate, not just DNA. Others treat it as a genuine theory of cultural evolution. Taken literally, it predicts that cultural evolution works like this. A meme impels its bearer to broadcast it, and it mutates in some recipient: a sound, a word, or a phrase is randomly altered. Perhaps, as in Monty Python’s Life of Brian, the audience of the Sermon on the Mount mishears “Blessed are the peacemakers” as “Blessed are the cheesemakers.” The new version is more memorable and comes to predominate in the majority of minds. It too is mangled by typos and speakos and hearos, and the most spreadable ones accumulate, gradually transforming the sequence of sounds. Eventually they spell out, “That’s one small step for a man, one giant leap for mankind.”
I think you’ll agree that this is not how cultural change works. A complex meme does not arise from the retention of copying errors. It arises because some person knuckles down, racks his brain, musters his ingenuity, and composes or writes or paints or invents something. Granted, the fabricator is influenced by ideas in the air, and may polish draft after draft, but neither of these progressions is like natural selection. Just compare the input and the output—draft five and draft six, or an artist’s inspiration and her oeuvre. They do not differ by a few random substitutions. The value added with each iteration comes from focusing brainpower on improving the product, not from retelling or recopying it hundreds of thousands of times in the hope that some of the malaprops or typos will be useful.
Stop being so literal-minded! respond the fans of cultural evolution. Of course cultural evolution is not an exact replica of the Darwinian version. In cultural evolution, the mutations are directed and the acquired characteristics are inherited. Lamarck, while being wrong about biological evolution, turned out to be right about cultural evolution.
But this won’t do. Lamarck, recall, was not just unlucky in his guess about life on this planet. As far as explaining complex design goes, his theory was, and is, a non-starter. It is mute about the beneficent force in the universe or all-knowing voice in the organism that bestows the useful mutations. And it’s that force or voice that’s doing all the creative work. To say that cultural evolution is Lamarckian is to confess that one has no idea how it works. The striking features of cultural products, namely their ingenuity, beauty, and truth (analogous to organisms’ complex adaptive design), come from the mental computations that “direct”—that is, invent—the “mutations,” and that “acquire”—that is, understand—the “characteristics.”
Models of cultural transmission do offer insight on other features of cultural change, particularly their demographics—how memes can become popular or unpopular. But the analogy is more from epidemiology than from evolution: ideas as contagious diseases that cause epidemics, rather than as advantageous genes that cause adaptations. They explain how ideas become popular, but not where ideas come from.
Many people unfamiliar with cognitive science see cultural evolution as the only hope for grounding wispy notions like ideas and culture in rigorous evolutionary biology. To bring culture into biology, they reason, one shows how it evolved by its own version of natural selection. But that is a non sequitur; the products of evolution don’t have to look like evolution. The stomach is firmly grounded in biology, but it does not randomly secrete variants of acids and enzymes, retain the ones that break down food a bit, let them sexually recombine and reproduce, and so on for hundreds of thousands of meals. Natural selection already went through such trial and error in designing the stomach, and now the stomach is an efficient chemical processor, releasing the right acids and enzymes on cue. Likewise, a group of minds does not have to recapitulate the process of natural selection to come up with a good idea. Natural selection designed the mind to be an information processor, and now it perceives, imagines, simulates, and plans. When ideas are passed around, they aren’t merely copied with occasional typographical errors; they are evaluated, discussed, improved on, or rejected. Indeed, a mind that passively accepted ambient memes would be a sitting duck for exploitation by others and would have quickly been selected against.
The geneticist Theodosius Dobzhansky famously wrote that nothing in biology makes sense except in the light of evolution. We can add that nothing in culture makes sense except in the light of psychology. Evolution created psychology, and that is how it explains culture. The most important relic of early humans is the modern mind.