3. Elevated CO2 in the atmosphere and oceans
4. Global nitrogen fertilization
5. Ozone depletion in Earth’s stratosphere
Increases in the scale and extent of human activity in the last two centuries have brought about environmental changes that affect most of the globe. These global changes include directional shifts in climate, greenhouse gas concentrations, nitrogen fixation, and stratospheric ozone depletion. They also include biotic changes such as land cover change, biological invasions, and global loss of biodiversity. In this chapter, we focus on the responses of ecological communities and ecosystems to directional changes in climate, atmosphere, and global biogeochemistry. Our understanding of these responses comes from observations of trends in nature, experiments manipulating global change factors at small scales over years to decades, and predictive models. We consider all of these sources, with an emphasis on the empirical knowledge derived from observations and experimentation.
biodiversity. The totality of the inherited variety of all forms of life across all levels of variation, from ecosystem to species to gene (E. O. Wilson).
biogeochemistry. The cycles of matter and energy that transport the Earth’s chemical components through time and space, and the chemical, physical, geological, and biological processes and reactions that govern the composition of the natural environment.
biomes. Generalized regional or global community types, such as tundra or tropical forest, characterized by dominant plant life forms and prevailing climate.
community. A group of interacting species living in a specified area. Communities are often defined by the dominant vegetation types, such as maple-oak or sagebrush. However, community composition is dynamic as species dominance and diversity shift in space and time.
ecosystem. An ecosystem is a complex system formed by the interactions of living (biotic) and nonliving (abiotic) components, which shape each other through exchange and material flows. An ecosystem can be bounded more or less arbitrarily and can range in scale from an ephemeral pond to the entire globe but most often refers to a landscape-scale system characterized by one or a specified range of community types (e.g., grassland ecosystems).
eutrophication. An increase in an ecosystem’s plant production resulting from nutrient inputs, often with undesirable effects such as excessive plant decay, oxygen deprivation, and water quality declines in aquatic systems.
nitrogen fixation. The conversion of inert atmospheric dinitrogen (N2) to nitrate and ammonia that can be taken up by organisms.
phenology. The timing of recurring biological phenomena, ranging from annual budburst and senescence in plants to the onset of animal migrations, egg laying, and metamorphosis.
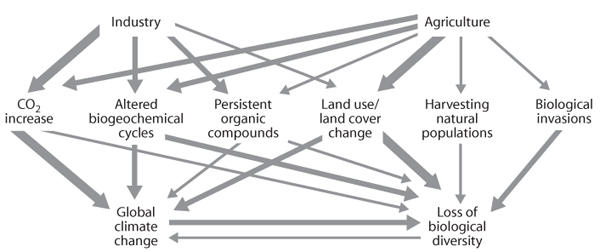
Humans, like all other organisms, modify their environment. Human modification, unlike that of other organisms, however, is drastically altering ecosystems over the entire globe through an explosion in human numbers and the scope of their activities. As anthropogenic impacts on the environment cause changes in biological systems, those changes in turn cause further changes in the environment, resulting in complex feedbacks and interactions. This process is described as global change. Many, although not all, global changes originate from transported chemical effects of human activity, such as carbon dioxide (CO2) emissions, aerosols, CFCs, and NOx emissions. Global changes also include direct effects on biological systems, such as habitat destruction, invasive species, and biodiversity loss. Finally, global changes interact extensively—for example, tropical forest destruction releases large amounts of greenhouse gases that contribute to global climate change (figure 1). In this chapter, we focus on climate change, CO2, nitrogen (N) deposition, and ozone depletion. Habitat loss and degradation, invasive species, and loss of biodiversity are discussed elsewhere in this volume, but we address them in the context of interactions with climate and atmospheric change.
Figure 1. The interactive nature of human activities and resulting global changes. (Adapted from Vitousek et al., 1997b)
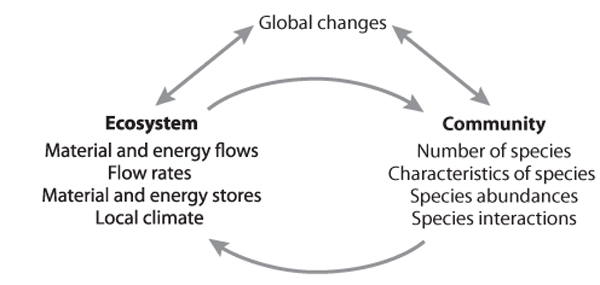
Communities and ecosystems respond to global changes interactively (figure 2). Global changes can affect ecosystem processes, such as through altered biogeochemical cycles, in ways that then influence the resident community. Conversely, global changes can drive community shifts directly, in ways that then affect ecosystem processes, such as when a shift from tropical forest to savanna vegetation drastically reduces local rainfall. The result is a feedback loop between community and ecosystem change, one that can buffer or amplify global change effects over time and result in long-term, ongoing changes even after a particular global change driver has stabilized. For example, an abrupt increase in temperature might reduce grassland soil moisture slightly for the first several years. The same warming over decades could transform that grassland to a woody shrubland, with much more extensive effects on water relations as the dense, shallow, seasonal grass roots are replaced by deep, well-spaced, perennial taproots.
Earth’s climate is strongly influenced by atmospheric concentrations of greenhouse gases and aerosols. Greenhouse gases warm the Earth’s surface by trapping heat in the lower troposphere, and aerosols contribute to cloud formation and cool Earth’s surface. Emissions of carbon dioxide (CO2), the most important human-generated greenhouse gas, come primarily from fossil fuel combustion and land use change. Two other important greenhouse gases, methane and nitrous oxides (NOx), arise mainly from agricultural production.
Human-caused increases in greenhouse gas concentrations now have accelerating effects on Earth’s climate. In the twentieth century, Earth warmed by an average of 0.75°C, and precipitation increased by 2%. Sea level rose 0.2 m as a result of thermal expansion of the world’s oceans. Rates of change were at least twice as fast during the later half of the century compared to the first and are continuing to increase with each year as greenhouse gas emissions increase. Changes in wind patterns, ocean salinity, and the periodicity and magnitude of extreme events have also occurred, including increased drought, tropical storms, heavy rainfall, and heat waves. If CO2 doubles by the end of this century as predicted, global mean temperatures will likely increase in the range of 2° to 4.5°C. Local changes will be more extreme: in the last century, at high latitudes, temperature increases have been nearly double that of the global mean, and precipitation has increased or decreased by as much as 40% in some locations.
Species populations respond to changes in climate through changes in abundance, distribution, and, in some cases, rapid evolutionary change. Species differ in their capacity to tolerate, adapt, and move, causing existing communities to disassemble and re-form anew. Many “no-analog” communities—those made up of combinations of species not currently found together— are expected to emerge. No-analog communities have existed in the past. For instance, during the last glaciation event in the United States 17,000 to 12,000 years ago, an ecosystem including spruce, sedge, oak, ash, and hophornbeam was abundant. These tree species do not occur together in the United States today. John Williams and colleagues estimate, based on climate simulation models, that 4% to 39% of the world’s land area will experience novel combinations of climate variables and almost certainly will develop no-analog communities. Such novelty means it will be exceedingly difficult to forecast future conditions.
Figure 2. Interactions among global changes and ecosystem and community responses.
Despite this challenge, predictive modeling studies have been an important avenue of climate change research to explore community and ecosystem change. Commonly, models are used to simulate changes in the distribution of biomes or plant functional types (PFTs) rather than individual species. Two prevalent types of vegetation modeling approaches are equilibrium biogeography models and dynamic global vegetation models (DGVMs). Equilibrium models provide “snapshot” views of what the distribution of biomes might look like given a particular CO2 level in the future. Dynamic models are gaining popularity because they help address how ecosystems might transition between states and can integrate vegetation dynamics with biogeochemical processes to explore changes in ecosystem function as well as structure.
Broadly, both equilibrium and dynamic vegetation models show major poleward shifts in cold-limited biomes as a result of climate change. For instance, tundra is predicted to decline in distribution as it will largely be “pushed off” the North American and Eurasian continents with the northward expansion of boreal forest. Boreal forest is also expected to decline as temperate forests expand northward. Dynamic models predict these vegetation changes will result in large productivity increases in northern latitudes. Biomes that are limited by water availability, rather than temperature, show more complex responses depending on the balance among precipitation change, hydrological response, and physiological adjustments by organisms.
Communities
Warming in the last century has been associated with shifts in species distributions upward in elevation and poleward. Camille Parmesan and Gary Yohe measured changes in the range boundary of 99 birds, butterflies, and alpine herbs. They found that range limits have moved on average 6.1 km per decade toward the poles or meters per decade upward in elevation. In another review, 80% of 434 species studied over time showed increased abundance in northern or high-elevation locations. For instance, J. P. Barry and colleagues compared surveys of intertidal marine invertebrates in the Monterey Bay, California, in the early 1930s to surveys in the early 1990s. These surveys show a clear pattern of increased abundance of southern species and decreased abundance of northern species.
Changing climate also affects the timing of climatecued events in organism life cycles and, in some cases, could desynchronize important trophic or mutualistic interactions among species. For instance, a bird species that uses photoperiod to cue migration may not shift its behavior although its caterpillar food may emerge earlier in the spring because of higher temperatures. Terry Root and colleagues report that for 694 species, spring events occurred an average of 5.1 days earlier per decade since 1951. On average, birds are shifting faster than invertebrates and amphibians, and trees are shifting much more slowly. Faster rates of change for some organisms compared to others means there will be strong selection pressure on species to shift food sources, nesting behavior, or other habits. Species that are generalists will likely be favored over specialists.
In the oceans, warming has marked impacts on coral reef communities, which support an estimated 25% of marine biodiversity. Corals themselves are colonies of small animals that feed by filtering plankton out of the water and secreting calcium carbonate skeletons. Inside of corals live symbiotic algae that provide food and give corals their bright colors. In response to various stresses including thermal warming, corals “bleach” because algae are lost from hosts, exposing the white calcium carbonate skeleton underneath. Bleaching events both reduce coral abundance and change community composition because coral species vary in sensitivity to warming. Corals also have high genetic diversity, and with warming, more resistant corals and symbionts could spread, buffering the survival of populations. This evolutionary response, however, might not keep pace with warming rates in combination with other stresses including overfishing, pollution, and ocean acidification.
Ecosystems
Warming at a global scale accelerates the water cycle, with net precipitation increases. However, on land, warming also increases water losses through evapotranspiration. Net drying has occurred and is expected in many parts of the world, with strong seasonal effects. Summer drying in the boreal forest and many montane regions, as a result of both earlier snowmelt and increased water losses, has produced forest diebacks in areas such as the southwestern United States and increased wildfire frequency, extent, and severity over large regions. Warming can also affect precipitation type: in California’s Sierra Nevada range, winter precipitation is shifting from snow to rain. The downstream implications are substantial, as flows shift toward winter and spring flood pulses rather than gradual snowpack melt over several months.
Warming effects on phenology influence productivity and other material and energy exchanges at local to global scales. In the northern hemisphere, the earlier onset of spring has led to an earlier and longer annual period of draw-down in atmospheric CO2 caused by plant photosynthesis and carbon uptake. When moisture is available, warming often accelerates and extends the annual duration of decomposition, speeding the return of carbon from soil and plants to the atmosphere. This effect, although also shaped by other global changes, could provide a positive feedback to climate change by shifting carbon stores from ecosystems to the atmosphere. In field experiments simulating global changes, warming increases productivity through the winter in Mediterranean grasslands but can also accelerate the end of the growing season. The net effect of warming on productivity depends on the degree to which production is limited by temperature versus by moisture or other resources. Ultimately, it depends most on how communities change in response to warming; for example, a shift from soft-bodied plants toward woody plants in meadow or tundra ecosystems strongly affects productivity and its distribution above and below ground.
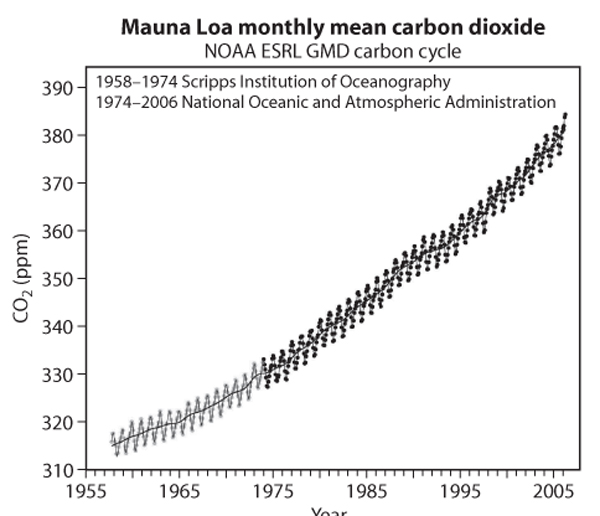
Atmospheric concentrations of CO2 have increased steadily since the advent of the industrial era and are accelerating each year (figure 3). The source of these rising levels is emissions of CO2 from human activity, mainly in the forms of fossil fuel combustion and land use change. Current atmospheric CO2 concentrations, revealed by ice core samples, are dramatically higher than at any time in the past 650,000 years.
Besides the climate-mediated effects explored earlier, rising atmospheric CO2 levels directly and profoundly influence communities and ecosystems because CO2 is a necessary input for photosynthesis. Most plants on Earth acquire CO2 for photosynthesis by opening stomatal pores in their leaves that let CO2 in and water vapor out through simple diffusion. Higher CO2 levels allow plants to acquire more CO2 per unit of time their stomata are open, permitting more rapid photosynthesis (58% faster on average across 60 studies) and growth. As a result, if other necessary resources are not limiting, elevated CO2 accelerates growth and can increase ecosystem productivity. This effect has been observed in growth chamber experiments, agricultural fields, and some natural ecosystems. In other natural ecosystems, however, other resources such as soil nitrogen limit plant growth, and increased CO2 shifts the competitive balance among species in the plant community rather than increasing overall productivity.
Figure 3. The longest record of Earth’s atmospheric carbon dioxide concentrations was begun by Dr. Charles Keeling and is maintained by the National Oceanic and Atmospheric Administration. Annual peaks and valleys occur because most of the world’s land area is in the Northern Hemisphere. CO2 levels drop in the summer when Northern Hemisphere plants are growing and rise in the winter when plants reduce activity.
By allowing plants to open stomata less, rising atmospheric CO2 also increases plant water use efficiency, on average, by a substantial 20%. This effect noticeably reduces ecosystem water losses to plant transpiration and increases soil moisture content in a range of herb-dominated ecosystems, including tall-grass prairie, salt marsh, crops, and several grasslands. Responses in woody ecosystems, especially conifer forests, are less clear because the behavior of woody plant stomata is less responsive to elevated CO2. Even relatively large gains in water storage, however, could be more than offset by increased evapotranspiration driven by CO2-induced climate warming. The net effects of rising atmospheric CO2 on ecosystem water balance, especially in arid ecosystems with more direct evaporation from soil, could be negative.
Plant-derived nitrogen is an essential source of protein to herbivore communities. Tissue quality in plants grown in CO2-enriched environments tends to be lower than in plants grown in ambient conditions because high CO2 concentrations dilute tissue nitrogen (N). The feeding intensity of herbivores depends on plants’ carbon-to-nitrogen ratio (C:N), leaf thickness, and concentration of defensive secondary compounds. Experiments consistently find that insects compensate for lower tissue quality by increasing consumption rates. Increased feeding may not compensate for decreased tissue quality in all cases however. Some studies show decreased growth rates of insects reared on CO2-enriched plant tissue, depending on the plant species involved, indicating variable plant response to CO2 enrichment. Herbivores may therefore shift feeding preferences to plants that maintain lower C:N in response to CO2 enrichment and produce a change in top-down control on plant community composition.
A troubling change resulting from increased CO2 emissions is acidification in marine ecosystems. Oceans currently absorb about a third of the CO2 released through fossil fuel combustion. As dissolved CO2 concentrations in oceans increase, so does acidity—first in the upper layers, then gradually in deeper waters. The effects of ocean acidification on marine communities are only beginning to be understood, but laboratory studies show that acidification directly threatens corals, crustaceans, and other marine organisms that use calcium carbonate to build shells and skeletons. Acidic waters dissolve calcium carbonate shells and make it increasingly difficult for organisms to precipitate calcium carbonate for shell building in the first place. Acidification rates to date vary across the globe and peak in the Atlantic; the consequences of continued acidification could include widespread losses of organisms, disrupting marine food webs and eliminating entire communities such as coral reefs.
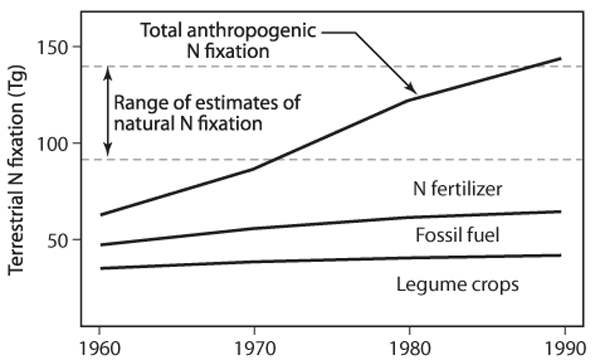
Nitrogen is abundant in the atmosphere in a biologically inert form called dinitrogen (N2). Human activity has vastly altered the global nitrogen cycle by fixing nitrogen deliberately for fertilizer and inadvertently during fossil fuel combustion (figure 4). Until the twentieth century, nitrogen fixation was accomplished primarily by bacteria in the root nodules of legume plants. In 1913, with the advent of the Haber-Bosch process, humans began to fix atmospheric nitrogen synthetically to produce ammonia fertilizer. The amount of industrially fixed nitrogen has since increased steadily. Humans now fix more nitrogen than all other natural sources combined. Synthetic fertilization use is widespread since the green revolution in the 1970s and is anticipated to increase by 70 in the next 20 years. Fertilizer runoff from agriculture affects aquatic communities and ecosystems around the world. In addition, fossil fuel burning, especially in automobiles, emits nitrous oxides (NOx), which are converted to nitrate and nitric acid in the atmosphere and deposited directly on vegetation and into bodies of water. Nitrogen deposition contributes to acid rain and ecosystem fertilization in large areas of the globe, with its most significant effects around and downwind from heavily industrialized and densely populated regions.
Much of the nitrogen applied to agricultural fields migrates into waterways. Nitrogen is a vital nutrient for organisms, but in excess amounts, it can restructure entire food webs and significantly decrease species diversity. In both freshwater and marine aquatic systems, excess nutrients cause algal blooms (“red tides”) that block light and deplete oxygen, creating “dead” zones. In the Black Sea, for example, heavy sewage, fertilizer inputs, and industrial waste from over 15 countries resulted in year-round severe eutrophication, frequent red tides, and fish kills. In concert with an accidentally introduced exotic jellyfish species that benefited from the high nutrient conditions, this fertilization drove 75% of commercial fish species to extinction in the 1980s.
Figure 4. Human fixation of nitrogen over time through fertilizer production, fossil fuel combustion, and the growth of legume crops, compared to all natural sources of N fixation. (From Vitousek et al., 1997a)
Nitrogen also reaches terrestrial systems from automobile and other industrial exhaust, with varying effects. In systems limited by scarce nitrogen, nitrogen deposition tends to increase productivity and carbon storage and can dramatically reduce biodiversity. In historically nutrient-poor ecosystems such as those with serpentine and calcareous soils, nitrogen deposition can dramatically increase productivity. In so doing, nitrogen deposition can reduce or eliminate endemic assemblages that are ill-adapted to compete against faster-growing invaders that are better able to exploit high soil fertility.
Ozone (O3) in the Earth’s stratosphere absorbs 95% of the ultraviolet radiation from the sun, which otherwise would destroy life. Chlorofluorocarbons (CFCs), used extensively for cooling in refrigerators and in aerosol products until an international ban in 1996, are mobile, long-lived, stable compounds that degrade in the stratosphere and destroy ozone. Ozone is a highly selective absorber of UV-B light, so the ratio of UV-B light to other wavelengths increases as ozone decreases. The effects of ozone depletion are unevenly distributed around the globe and pronounced at high latitudes during the winter and early spring, when the formation of stratospheric ice particles above the poles provides surfaces for rapid decomposition of CFCs and the breakdown of ozone.
Our knowledge of community and ecosystem changes that result from increased UV-B light comes largely from small-scale experiments in confined ecosystems called mesocosms. Investigators can manipulate UV-B levels directly in mescosms, increasing UV-B with radiation-emitting lamps or decreasing it with selective filters.
Organisms exhibit wide-ranging sensitivity to ultraviolet radiation, both within and across taxonomic groups such as higher plants and fungi. Species with greater pigmentation or body coverings such as hair are better protected from the harmful effects of UV-B light, whereas amphibians are poorly protected and highly sensitive. Microbes vary widely in sensitivity. Thus, increased UV-B light alters community composition toward less-sensitive species and affects species interactions. Max Bothwell and colleagues used an experimental river flume facility to test how algal and invertebrate communities exposed to different amounts of sunlight were affected. They found that the sensitivity to UV-B is higher in algal grazers than in algae, altering the balance between primary producers and consumers in shallow benthic communities.
UV-B–driven changes in trophic or community structure can alter carbon and nutrient cycling in ecosystems. In a subarctic heath system, David Johnson and colleagues showed that increased UV-B radiation alters soil microbial C:N through microbial community changes, with potentially far-reaching consequences for plant communities. Increased UV-B radiation can also directly affect ecosystem functioning. In semiarid systems, increased UV-B can accelerate decomposition and carbon cycling. Interactions between increased UV-B light and other global changes, although likely important, remain poorly understood.
Many anthropogenic global changes are occurring at once. Their interactions, rather than their individual effects, will have the biggest implications for ecological responses.
Some global changes are linked through positive feedbacks, so increases in one will cause increases in another. Habitat destruction, particularly in the world’s forests, contributes to greenhouse gas emissions and warming: climate changes are increasing wildfire and the extent of forest dieback in some regions, accelerating forest loss. Other global changes are linked through negative feedbacks. For example, aerosol emissions partially counteract the warming effects of greenhouse gases by reducing the amount of solar radiation reaching Earth’s surface.
In general, community and ecosystem responses to the suite of ongoing, interacting global changes cannot be predicted from studies of individual global changes. As study of global change interactions grows, it reveals a range of additive to synergistic or idiosyncratic effects. When nitrogen deposition and CO2 increases occur together, the added nitrogen can alleviate nutrient limitation of productivity responses to CO2. The result can be much greater productivity increases than under elevated CO2 or nitrogen fertilization alone. Elevated CO2 alone can lead to greater ecosystem uptake of atmospheric carbon, an effect some hope could slow the accumulation of greenhouse gases in the atmosphere. However, warming can offset this effect entirely by speeding up release of carbon into the atmosphere from decomposing matter.
Interactions among the global changes emphasized in this chapter are complicated further by their interaction with other human transformations of the environment. Many global changes—most notably nitrogen deposition, aspects of climate change such as wildfire and accelerated forest dieback, and habitat destruction—exacerbate the spread of invasive species by favoring weedy, fast-growing, disturbance-adapted species over slow-dispersing, long-lived, and specialized ones. Climate change and invasive grasses in regions from the Great Basin to Hawaii interact to transform wildfire regimes through a combination of hot, dry weather and expanding flammable fuel loads. In boreal lakes, emissions of nitrogen and sulfur (NOx and SOx) interact with warming to reduce dissolved organic matter (DOC). Lower DOC then allows more UV-B light (which has increased particularly at these high latitudes from stratospheric ozone depletion) to penetrate the lake surfaces, rendering large areas unable to support aquatic life. With less life in them, lake productivity declines, further reducing DOC.
A picture emerges of natural systems altered drastically by the accumulating effects of multiple global changes. Ecological science can provide some insight into the nature and magnitude of ongoing and potential future changes through experiments, historical study, and models. These insights can inform society’s work to flag the undesirable effects and, through a combination of adaptive stewardship and steps to slow or reverse global change rates, avert them. In a climate of complex global change interactions and uncertainty about the precise trajectories they will take, safeguarding the capacity of ecosystems to adapt to change is a minimum step to preserve options for the future.
Lovejoy, T. E., and L. J. Hannah. 2005. Climate Change and Biodiversity. New Haven, CT: Yale University Press.
Mooney, H. A., and R. J. Hobbs. 2000. Invasive Species in a Changing World. Washington, DC: Island Press.
Vitousek, P. 1994. Beyond global warming: Ecology and global change. The Robert H. MacArthur Award Lecture, Ecology 75: 1861–1876.
Vitousek, P. M., J. D. Aber, R. W. Howarth, G. E. Likens, P. A. Matson, D. W. Schindler, W. H. Schlesinger, and D. G. Tilman. 1997a. Human alteration of the global nitrogen cycle: Sources and consequences. Ecological Applications 7: 737–751.
Vitousek, P. M., C. M. D’Antonio, L. L. Loope, M. Rejmanek, and R. Westbrooks. 1997b. Introduced species: A significant component of human-caused global change. New Zealand Journal of Ecology 21: 1–16.
Vitousek, P. M., H. A. Mooney, J. Lubchenco, and J. A. Melillo 1997c. Human domination of Earth’s ecosystems. Science 277: 494–499.