6. Reconciling historical and mechanistic interpretations
Competition has long been thought to play a foremost role in the organization of ecological communities, and this has been a core concept in the building of niche theory. However, many observed patterns in nature are difficult to reconcile with the predictions of niche theory, and they reflect the historical nature of community assembly. The neutral theory of biodiversity has recently been developed to provide an alternative interpretation of patterns in community organization in the absence of competitive difference among coexisting species.
Hutchinsonian ratio. Body size ratio of the larger species over the smaller species in a pair of species; niche theory predicts that co-occurring species should have larger body size ratio than expected by chance
neutrality. Assumption of equivalence in individuals’ prospects of reproduction or of death, irrespective of the species they belong to
phylogenetic overdispersion and clustering. The tendency of species to be on average more or less (respectively) evolutionarily related in a sample than in the larger species pool
species abundance distribution Φ(n). Number of species with exactly n individuals in a sample
Ecological communities are complex assemblages of organisms shaped by environmental constraints and interacting through a variety of ecological processes but also reflecting historical contingencies. Understanding these processes has been a central goal of animal and plant ecology for almost a century. Early on, researchers put forward the idea that communities were tightly organized associations of species, with sharp boundaries, and that they were amenable to classification, just as species can be classified taxonomically. This idea may be related to the famous work of the Russian experimentalist Georgyi Gause. He and others developed research projects on simple species assemblages easily amenable to experiments, such as yeast and paramecia (Gause, 1934). Because these systems included only a few species, typically two, and controlled environmental conditions, it was possible to find the theoretical conditions under which species may coexist stably in association. Nicely, these experiments could be reframed into the mathematical theory developed by the Italian mathematician Vito Volterra (1931). The major finding of both theory and experiments was that, of two species with identical ecological requirements and competing for limited resources in a stable environment, one will eventually exclude the other. This result later developed into a fundamental principle, called the principle of competitive exclusion.
The view that ecological communities should be the result of tight species association did not remain un-criticized over these years. In 1926, the plant ecologist and taxonomist Henry Gleason, for instance, suggested that species do not, as a rule, live their lives in tight associations, a view that was echoed in 1935 by one of the most prominent British ecologists of the first half of the twentieth century, Sir Arthur Tansley. Some authors even suggested the extreme view that species interactions play no role in community organization (Andrewartha and Birch, 1954). This longstanding debate in ecology was admirably synthesized by Robert MacArthur (1972). Although he acknowledged the role of history in shaping species assemblages, he noted “unravelling the history of a phenomenon has always appealed to some people and describing the machinery of the phenomenon to others.” He further suggested that the ecologist tends to be “machinery oriented,” whereas the biogeographer tends to be “history oriented.” To MacArthur, useful patterns of species diversity are shaped by repeatable phenomena, not by chance events.
An important issue here is: How do experimental findings generalize to natural communities and to more than a couple of species? This question was most clearly addressed in 1959 by the American limnologist G. Evelyn Hutchinson, one of the most influential ecologists of the twentieth century. Central to the understanding of species coexistence is the concept of the ecological niche of a species, the biological equipment of a species in relation to competition and to environmental conditions. For animals, niches may be defined in terms of the food of a species, as developed early on by Charles Elton. For plants, an operational definition of the niche is more problematical, even though much research has been devoted to quantifying plant life-history strategies. Although the concept of the niche was developed historically earlier, it was Hutchinson and his students, including MacArthur, who proposed an operational definition of this concept to most ecologists (for a historical overview, see Sharon Kingsland, 1995).
Following Gause’s principle, one would expect that competition plays a role in species coexistence if interspecific competition is more intense than intraspecific competition. This implies that one could assess the role and nature of competition experimentally. Studies on this topic are almost innumerable. In 1983, Thomas Schoener and Joseph Connell both reviewed this huge literature (over 200 published studies at that time and many more since then), and they found that well over half of the published studies convincingly demonstrated some effect of competitive exclusion. However, they also found that studies with more species tended to have fewer species evidencing competition. This suggests that interspecific competition may be more diffuse in species-rich communities. Also important is the fact that although numerous researchers have claimed to quantify competition, as had been reviewed by Connell and Schoener, few have actually addressed the importance of competition in shaping ecological communities. Deborah Goldberg and Andrew Barton (1992) found that, in the plant community, no more than 89 studies had addressed patterns of competition, 63% of which actually focused on a single species in a background of competing species. Fewer than half a dozen experiments had suitably addressed the consequences of plant competition on communitywide patterns by the early 1990s.
After almost a half-century of research on the role of competition on community organization, one would think that a clear theory had emerged and that quantitative and predictive theories are by now available. Not quite so. Predictions of niche theory have proven difficult to test. As a result, the original quest for a universal role of competition in community assembly rules has turned into a more ramified research program, including a search for the relative importance of processes aside from competition.
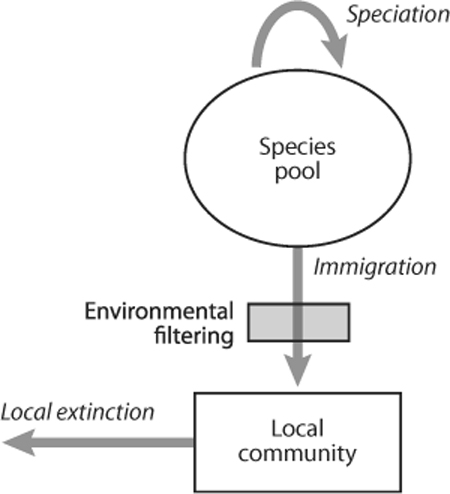
Dispersal limitation has long been known to limit the distribution of species and hence to contribute to the makeup of communities. Just by chance, a species may be absent from a site where it would have thrived. This simple remark has resulted in an elegant model by John G. Skellam in a famous paper (1951). Likewise, demographic processes may cause a rare species to be present in a community by chance rather than because it is a superior competitor (see figure 1). These two processes have long been considered a nuisance by ecologists but are in fact essential to explain many patterns of community organization. In 1987, Robert Ricklefs called this bias against nondeterministic interpretations of community assembly the “eclipse of history” in ecology.
This view has recently been reinforced by the neutral theory of biodiversity. The concept of neutrality is over three decades old in ecology: it was developed in 1976 by Hal Caswell, who used it as a null model for testing the importance of competition. Caswell constructed his model by assuming that co-occurring species were not interacting. This approach remained largely unnoticed, with the notable exception of work by Stephen Hubbell (1979, 1986, 1997), who tested theories of tree species coexistence in tropical rain forests. Hubbell’s thinking eventually matured into an important book (Hubbell, 2001), which offered a novel interpretation of the neutral theory. In his book, Hubbell assumes that individuals in a community all have the same prospects of birth and death and that species interaction is fully symmetrical. In this sense, Hubbell’s theory is neutral. As Hubbell remarked, many people are confused by the word “neutral,” which, to many, is equivalent to “nothing is going on.” However, the neutral assumption does not imply that individuals or species do not compete with each other, only that competition should be symmetrical: an individual of species A has the same influence on an individual of species B as an individual of species B has on an individual of species A. Most importantly, Hubbell constructed a theory in which a local community is seen as a dispersal-limited sample of a regional species pool. In the regional species pool, species arise through speciation, a process that is summarized by a single parameter θ, equal to the product of the per capita speciation rate and the number of individuals in the region. At the local community level, the intensity of dispersal limitation is quantified by the immigration rate m, the probability that a new individual is an immigrant.
Figure 1. Local community assembly from a regional species pool. At the regional scale, the evolutionary processes dominate, in particular speciation. A local community is a filtered sample of this regional species pool because not all species can reach the site (dispersal limitation) and because not all species are able to settle in the local environment (environmental filtering). The immigration of new species is balanced by the local extinction of species through competitive displacement, demographic drift, or catastrophic events.
Beyond Caswell’s seminal work, Hubbell’s theory finds its mathematical inspiration in the neutral theory of molecular evolution, proposed in 1964 by Motoo Kimura, itself rooted in population genetics theory of the first half of the twentieth century. Kimura, impressed by the variability of patterns in the genetic data that were just becoming available at that time, suggested that most of these patterns could be caused by random changes in the course of molecular evolution rather than by deterministic processes. That most of the genetic variability was a result of chance rather than selection was utterly counterintuitive, and Kimura’s ideas were harshly criticized by the selectionist school. A direct consequence of this theory, however, was that if molecular evolution is neutral, then the cumulative amount of molecular divergence between groups could be a useful measure of the time since the split occurred.
The molecular clock concept, although much debated, has become a standard tool in population genetics and in phylogenetics. Moreover, powerful statistical methods have been developed from the neutral theory, and these were used to test a selective departure from neutrality (see Warren Ewens’ book on mathematical population genetics, republished in 2004). As in molecular evolution, Hubbell’s theory was bluntly criticized, which led to a series of tests of the hypotheses or the predictions of this theory. Brian McGill et al. (2006) offered an interesting overview of these critiques and concluded that there is an “overwhelming weight of evidence against neutral theory.” Despite this view, these authors and many others still believe that this approach provides an interesting framework for testing ecological hypotheses.
The next section summarizes evidence for the role of competitive and neutral processes in the organization of communities.
One of the most classic ecological patterns in community ecology is the species abundance distribution, the number of species of a given abundance in a biodiversity survey. Let us call Φ(n) the number of species with exactly n individuals in such a survey. With this definition, the total number of individuals in the survey is 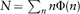 , and the total number of species is
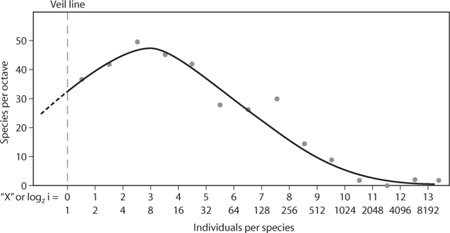
, and the total number of species is  . In a classic work, the geneticist Ronald Fisher (1943) provided a simple model for the species abundance distribution. The Fisher species abundance distribution was predicted to be a so-called log-series distribution, Φ(n) = αxn/n, where α and x are simple functions of S and N defined above (see for instance Evelyn C. Pielou’s 1975 book for a derivation of this result). Interestingly, the following relation: S = α ln (1 + N/α) holds in Fisher’s model. This suggests that α, also known as Fisher’s α in the ecological literature, is an unbiased index of diversity, if the model’s assumptions are met. Fisher used his model to explain data on Lepidoptera accumulated over years by two colleagues of his, Steven Corbet and C. B. Williams. The data suggested that most of the species in a survey should be represented by a few individuals. Later, Preston (1948) revisited this question, suggesting that empirical samples of a community would be better explained by the so-called log-normal distribution Φ(n) ∼ A exp (– B[ln (n/n0)]2), truncated at n = 1 because only species with at least one individual can be observed in a sample (figure 2). Hence, the left part of the species abundance distribution is “veiled” because of a limited sampling, a phenomenon sometimes referred to as Preston’s “veil line.”
. In a classic work, the geneticist Ronald Fisher (1943) provided a simple model for the species abundance distribution. The Fisher species abundance distribution was predicted to be a so-called log-series distribution, Φ(n) = αxn/n, where α and x are simple functions of S and N defined above (see for instance Evelyn C. Pielou’s 1975 book for a derivation of this result). Interestingly, the following relation: S = α ln (1 + N/α) holds in Fisher’s model. This suggests that α, also known as Fisher’s α in the ecological literature, is an unbiased index of diversity, if the model’s assumptions are met. Fisher used his model to explain data on Lepidoptera accumulated over years by two colleagues of his, Steven Corbet and C. B. Williams. The data suggested that most of the species in a survey should be represented by a few individuals. Later, Preston (1948) revisited this question, suggesting that empirical samples of a community would be better explained by the so-called log-normal distribution Φ(n) ∼ A exp (– B[ln (n/n0)]2), truncated at n = 1 because only species with at least one individual can be observed in a sample (figure 2). Hence, the left part of the species abundance distribution is “veiled” because of a limited sampling, a phenomenon sometimes referred to as Preston’s “veil line.”
Figure 2. Species abundance distribution plotted in logarithmic classes of abundance. The data correspond to a moth survey performed by C. O. Dirks in 1931–1934 at Orono, Maine (56, 131 collections for 344 species). (Reprinted with permission from Preston, F. W. 1948. The commonness, and rarity, of species. Ecology 29: 254–283)
How can these patterns of species abundance be explained by the processes of community organization? In 1957, Robert MacArthur proposed a connection between niche theory and the species abundance distribution. He reasoned that if the total niche space in a community could be graphically represented by a segment of unit length, then the niche of each species would be represented by a fraction of this segment, further assuming nonoverlapping niches. This is sometimes called a “niche preemption model” in the literature. He then suggested that the niche segment was partitioned at random into S subsegments (“broken stick”), each representing the niche of a species. Finally, assuming that the size of the niche space for a species was proportional to the abundance of this species, he was able to derive a mathematical formula for the species abundance distribution under this model. Although MacArthur’s “broken stick” model generally failed to reproduce empirical species abundance distributions, as he himself acknowledged, it eventually became a great source of inspiration for his colleagues, as it was a notable attempt to relate empirical patterns to the processes of niche partitioning at play in the organization of ecological communities.
It was subsequently discovered by Joel E. Cohen (1968) that the same result as MacArthur’s could be obtained simply by drawing S random variables from an exponential distribution and normalizing these numbers such that their sum is equal to 1. Hence, exactly the same mathematical form of the species abundance distribution can be obtained based on a niche-based argument, or based on a purely probabilistic argument. This line of reasoning also offered a useful connection with Fisher’s species abundance model. Let us assume that the species abundances are represented by S random variables x with the same two-parameter distribution,
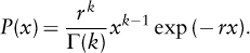
This is called the Γ distribution in probability theory. Then Cohen’s exponential model holds if k = 1, and Fisher’s model is recovered in the limit when k tends to zero. More biologically relevant models were proposed later on, notably in 1996 by Steinar Engen and Russell Lande (see Lande et al., 2003) and by Hubbell (2001).
There is no shortage of competing models predicting species abundance distributions. However, a proper statistical theory able to assess how well a model fits data is more difficult to obtain. Because the species abundance data are far from normal, a more general method is based on likelihood theory. The likelihood function—a term coined by Fisher—quantifies how likely a model is for a set of model parameters and given the sample at hand. The model parameters may be estimated by the value that maximizes the likelihood function. It is a remarkable fact that for Hubbell’s dispersal-limited neutral theory, a likelihood function can be computed exactly based on the species abundance data of a local sample, as was shown by Rampal Etienne and Han Olff, and the parameters θ and m can therefore be estimated directly (Alonso et al., 2006). They constructed an alternative niche preemption model, for which they also constructed a likelihood theory. Using tropical forest tree species abundance data, they showed that the neutral model was more likely than the niche preemption model. Although much has been made of the fact that nonneutral models provide a better fit to empirical species abundance distributions, very few have actually based their results on Etienne and Olff’s exact statistical theory (McGill et al., 2006). For instance, Igor Volkov and his collaborators (2005) suggested that a community model including density-dependent regulation always outperformed the neutral model’s fits of empirical tropical tree species abundance distributions. With David Alonso and Rampal Etienne (2006), we reassessed this result based on an exact likelihood-based comparison, and we showed that their conclusion was inaccurate. These studies, as well as others, suggest either that nonrandom ecological processes are indeed much less important for community organization than chance events or that the species abundance distribution is an uninformative pattern to detect nonneutrality in ecological communities.
Species abundance distributions are useful patterns in a single ecological community. However, a most powerful test of the role of competition in community organization makes use of the variation in the species composition across local communities. Charles Elton, in his famous paper “Competition and the Structure of Ecological Communities” published in 1946, sought to provide a simple pattern to assess whether island bird community assemblages were organized by competition. He reasoned that if competition is prevalent in islands, then closely allied species that share many niche traits should not be observed together. He used this remark to devise an original test of the competition theory: if competition is at work, then congeneric species should more rarely co-occur than expected by chance because they usually tend to share the same niche. Hence, the species to genus ratio (or S/G ratio) should decrease as competition intensity increases. He measured many S/G ratios in islands of the United Kingdom archipelago and showed that they were indeed smaller than observed in the corresponding source pools of species (the main islands). One problem with Elton’s test, however, was that many genera typically tend to have a single species, although a few have many species. In other words, the S/G statistic is difficult to measure with a great reliability. Daniel Simberloff (1970) pointed out another deficiency in Elton’s reasoning. He showed that if larger areas are sampled, the number of both species and genera are expected to increase, but the S/G ratio should also increase. The larger S/G ratio in mainland areas compared with islands was found by Simberloff to be almost solely a result of this bias.
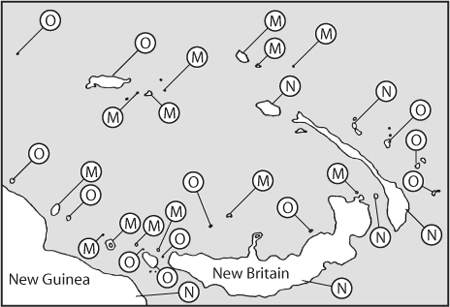
Figure 3. Checkerboard pattern of distribution for two related small cuckoo-doves in the Bismark archipelago. Symbol M stands for islands where only Macropygia mackinlayi occurs, N for islands where only Macropygia nigrirostris occurs, and O for islands where neither occurs. (Redrawn from Diamond, J. M. 1975. Assembly of species communities. In M. L. Cody and J. M. Diamond, eds. Ecology and Evolution of Communities. Cambridge, MA: Harvard University Press, 342–344.)
The study of species co-occurrence on archipelagoes was to become a very active field of research. In 1975, Jared Diamond published a large analysis of bird communities in archipelagoes east of the island of New Guinea (figure 3). Of special interest to him was to test the existence of assembly rules for these bird communities. One of these assembly rules stipulated that “some pairs of species never coexist, either by themselves or as part of a larger combination”; that is, one species excludes competitively the other if they were occurring on the same island. Although Diamond found evidence for this pattern in his data, it generated a critical evaluation by Connor and Simberloff (1979), who pointed out that Diamond had not tested them against a proper null assumption. As in the S/G ratio case, the suitable way of testing that some species combinations never exist in nature is to provide a suitable “space of the possible” resulting solely from random migration events into the island and to compare these possible configurations to the observed one. One way of testing Diamond’s rule, Connor and Simberloff proposed, would be to construct a matrix of species cooccurrences, such that an entry i, j is nonzero if species i and j co-occur, and zero otherwise. The next step would be to construct a “randomized” community by permuting the matrix of species co-occurrences according to different protocols, which has been made easy by computer programs. Connor and Simberloff (1979) then concluded, based on the graphic match of the null model compared with observed data, that allopatric speciation and limited dispersal would be equally valid explanations of the observed patterns. However, there are several ways of randomizing the data. One of the most fundamental critiques of Connor and Simberloff’s (1979) solution is that, as Jonathan Roughgarden (1983) puts it, “Islands do not reach into urns and draw out their species. There are real processes that bring species to islands.” Thus, the choice of a proper null model is a difficult one, and, following Roughgarden’s prescription, one should seek null models with a better biological motivation than random assortment models. A historical review of the alternative solutions developed to test Diamond’s fifth assembly rule was nicely summarized in 1996 by Nick Gotelli and Gary Graves (see their chapter 7), and a definitive test of the hypothesis was provided by Gotelli and McCabe (2002).
One puzzling problem with niche theory is that it can lead to contradicting predictions about the spatial distribution of species. The classic prediction is that related species should exclude each other competitively, as Elton suggested. A newer prediction, however, is that related species have similar abilities to withstand a given environment and hence are more likely to be found in the same habitat. This latter prediction suggests implicitly the existence of species “associations.” Similar species may compete, but they are found in the same environments simply because they have overlapping ecological requirements. Many studies have sought evidence for such habitat associations in plants by relating spatial patterns of species distribution to abiotic environmental factors. Another prediction, derived from the neutral theory, is that changes in species composition across samples should not depend on the environment or on species identities but only on pairwise geographic distance between samples. Richard Condit et al. (2002) tested this hypothesis using networks of tropical forest tree plots in Panama, Ecuador, and Peru. They found that the prediction of the neutral theory that species similarity should decline logarithmically with geographic distance was consistent with field observations. However, working on plants abundant in the understory of tropical forests (ferns and Melastomataceae), Hanna Tuomisto and her colleagues (2003) found that the predictions of the neutral theory were not met. Instead, they suggested that environmental variation best explains variation in diversity in their data. Ordination techniques have also been used to quantify the relative role of geographic distance and environmental variation in explaining patterns of species similarity. Karl Cottenie (2005) published a meta-analysis of distributional data on 158 species, and he found that most of the variation in biodiversity was explained by a combination of both factors rather than a single factor. This body of evidence suggests again that even though the neutral theory cannot alone predict patterns of species co-occurrence, neither can pure niche-based theories.
Niche theory makes predictions not only for the spatial distribution and co-occurrence of species but also for the appearance and behavior of the species that do co-occur. For instance, in 1945 David Lack reported that beak size for Darwin finches in the Galápagos islands depended on the whether the species were or were not co-occurring with other finch species. This fact had also been remarked upon by G. Evelyn Hutchinson (1959) in two “water boatman” species, aquatic insects in genus Corixa, found in sympatry throughout Europe (C. affinis and C. punctata). Although these two species are difficult to separate on the basis of their morphology or ecology, a striking difference between them is that one species was much smaller than the other. He reasoned that these differences in size were not a fact of chance but that they enable the species to partition the food web structure although their ecological requirements are the same. Because Hutchinson provided as a clear niche-theoretical prediction that the body size of co-occurring species should differ, the ratio of larger body size over smaller body size was later referred to as the “Hutchinsonian ratio.”
This study of body size ratios in co-occurring species was to become one of the most classic tests of competition theory in ecological communities. Two remarkable examples are here singled out. Thomas Schoener (1970) in a study of the Anolis lizards of the Lesser Antilles archipelago, a classic model in island biogeography, found that of the 27 Lesser Antillean islands, nine had two Anolis species, and these varied in size by a factor greater than 1.5. In contrast, in the single species islands, species tend to be intermediate in size. Abbott et al. (1977), in a monograph on the ecology of Darwin finches, also confirmed this pattern, showing that in islands where two species are found together, their beak size ratios tend to be large. This prediction was also tested in continental environments, for instance by Jim Brown (1973), who explored body size ratios in desert rodents in the United States.
As for previous ecological patterns, the relation between size ratios and competition was critically reassessed by Daniel Simberloff and William Boecklen (1981). To properly test whether size ratios in co-occurring species may be a signal of competition, they reasoned, a rigorous statistic of size overdispersion should be defined. When they ran this improved statistical test on published studies that had claimed to establish Hutchinsonian ratios in empirical data sets, they found that only a third of these claims were valid, the remaining two-thirds being not statistically different from a random assortment of species. For instance, even though both Schoener and Brown claimed their patterns of species co-occurrence should be explained by competition, Simberloff and Boecklen showed that chance would equally well explain their results.
Where do we stand today on Hutchinson’s body size ratios? Simberloff and his collaborators have paved the way for more rigorous tests, and they offered the alternative hypothesis that chance, rather than deterministic factors, might be responsible for at least some of the body size patterns observed in nature. Since then, however, tests of Hutchinson’s hypothesis have been mostly carried out by evolutionary ecologists, with hypotheses and methods that were largely inaccessible to community ecology during the 1970s and the 1980s. The next section summarizes these results.
That two different species should have a different size to partition the biotic environment was a remarkable enough hypothesis because it was amenable to testing. W. L. Brown and E. O. Wilson (1956) made an even more surprising remark: in the same species, the size may vary depending on whether or not they co-occur with a related species. As an illustration, they provided the striking example of two Old World bird species (genus Sitta). The species overlap only in Iran, and where they co-occur, one species tends to have much smaller beak and body sizes than when it is found alone, whereas the other species tends to have much larger beak and body sizes. This phenomenon was referred to by Brown and Wilson as “character displacement.” This topic has attracted a number of ecologists and evolutionary biologists. Tamar Dayan and Daniel Simberloff (2004) recently reviewed published empirical evidence, and they concluded that research over the past two decades has found convincing evidence in favor of character displacement. If one were to extirpate one of the two bird species in Brown and Wilson’s example, the other would likely return to its normal size.
In Hutchinson’s example, however, body sizes are realized differences, and they are maintained irrespective of whether species co-occur or not. What is the difference? The evolutionary time scales involved in the two processes differ; Hutchinson’s body size differences refer to realized evolutionary divergences. On the other hand, Brown and Wilson’s intraspecific differences demonstrate spectacularly their potential for evolutionary divergence. In 1983, Ted Case offered a new terminology to classify possible causes of large body size ratios: character adjustment is the adaptive change in species caused by character displacement, whereas character assortment is the mechanism of ecological competitive exclusion in species that have already evolved divergent characters.
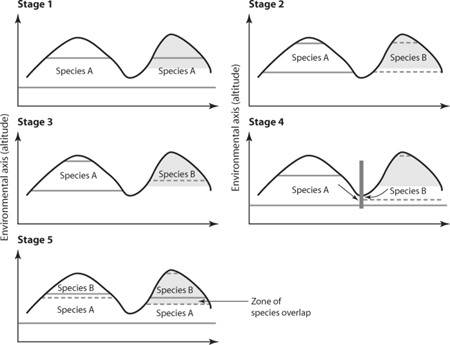
Is there a relation between character adjustment and character assortment, or, in other words, do plastic differences trigger events of speciation? To employ a wording more consistent with evolutionary theory, the issue is to know whether character displacement evolved before or after speciation was completed. If the former, then it is to be expected that character displacement may have played a role in the early stages of population divergence. It has long been thought that adaptive changes alone are in general insufficient to give rise to new species when the types are in sympatry. Instead, the primary cause of speciation is thought to be geographic isolation, followed by character divergence, as illustrated in figure 4. This question has been a central focus of evolutionary biology for several decades, and it remains a fertile point of contact between ecology and evolution, as demonstrated in Dolph Schluter’s (2001) review.
In a landmark study of Schoener’s Anolis lizards, Jonathan Losos (1990) proposed a new look at the causes of morphological divergence in species. Use should be made of the facts that co-occurring species have a shared evolutionary history and that this evolutionary history is reflected in the species phylogeny. If competitive exclusion is the prime mechanism for large body size ratios in two species, then it should be expected that the ancestral species should have an intermediate body size. Losos mapped events of body size changes onto the best phylogenetic hypothesis for genus Anolis available at the time, and he found that body size change did account for some of the observed body size ratios in two-species islands, but these were entirely caused by a single evolutionary event. He then suggested that most of the remaining differences in body size should be ascribed to character assortment rather than to character adjustment.
Such phylogeny-based analyses in community ecology have been scarce even after the heated debate over community organization processes in the 1980s. One explanation is simply that reliable phylogenetic hypotheses for groups of co-occurring taxa are difficult to construct. It is only with the rise of modern molecular phylogenetics and robust phylogeny reconstruction methods, based either on molecular sequences or on supertree methods, that these questions came to the fore. Ancestral state reconstructions and comparative methods became standard in the ecologist’s toolbox as a result of the book by Paul Harvey and Mark Pagel (1991). Then, ecologists began to include the historical information contained in phylogenies in tests of community organization. The underlying idea is to factor out lineage effects to assess the role of environment or of competition without potentially confounding historical factors. Losos (1996) and Webb et al. (2002) provided a comprehensive overview of this new line of thinking.
Figure 4. Stages of species formation. In this hypothetical example, species A is initially widespread (stage 1), but it becomes isolated as a result of a change in climatic conditions leading to its isolation in populations in two mountaintops (stage 2). After isolation, the two populations are exposed to different environmental conditions (gray zone on the right mountaintop versus white zone on the left one), leading to allopatric differentiation in the incipient species (stage 3). When the climatic conditions return to normal, the species come into contact again (stage 4), and they compete for space, potentially leading to character displacement and eventually to niche shift (stage 5). (Adapted from MacArthur, 1972; and from Wiens, J. J. 2004. Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evolution 58: 193–197)
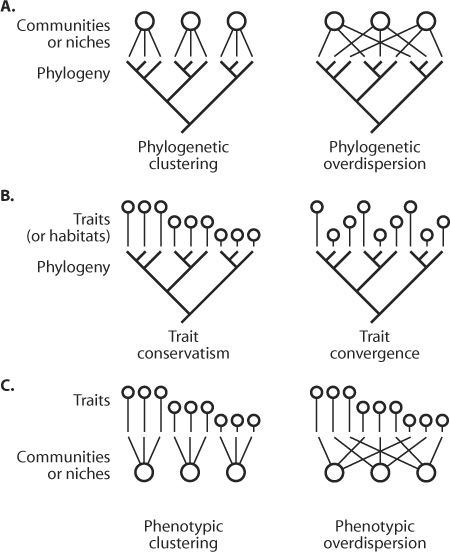
A fruitful use of these ideas consists in using the structure of the tree subtended by a local community and comparing it to that of the whole species pool. In the same way as in Elton’s (1946) test, the idea is that if competition is really shaping ecological communities, then species should be more overdispersed phylogenetically than any subtree of the same size drawn at random in the species pool. Suppose that we are working with trees of the Borneo rainforest, as in Cam Webb’s (2000) study, and that we have at hand a number of permanent 1-ha plots in which all trees have been censused. Assuming that the species in the same lineage share part of the same evolutionary history, they should have many similar niche features. Hence, they should be more prone to competitive exclusion. This implies that in any local community organized through competition, co-occurring species should be more phylogenetically dispersed than expected by chance in the regional pool’s phylogeny (phylogenetic overdispersion, figure 5). If, on the other hand, whole lineages have evolved some sort of habitat specialization, e.g., through adaptive speciation, then we should observe that species tend to be more phylogenetically clustered. This alternative is similar to Ted Case’s concepts of adjustment versus assortment, but for niches rather than for single characters.
Figure 5. This graphic representation illustrates different possible outcomes of tests based on species distribution data paired to a phylogenetic hypothesis (A), species traits paired to a phylogenetic hypothesis (B), or species traits paired to species distribution (C). The left panels show a case of conservatism in co-occurring species (A), in traits (B), or of phenotypic clustering in communities (or niches). The right panels show the opposite case of overdispersion. The idea that phylogenetically related species have similar niche traits and hence are more likely to compete is represented here by panel B, left. If this hypothesis holds, then phylogenetically related species should not co-occur, which is depicted here by panel A, right. (C) Classic tests of community ecology, such as Elton’s island models, that mostly ignore the phylogenetic relatedness among species. (Modified from Cavender-Bares, J., D. D. Ackerly, D. A. Baum, and F. A. Bazzaz. 2004. Phylogenetic overdispersion in Floridian oak communities. American Naturalist 163: 823–843)
It was possible for Webb to rely on a molecular phylogeny of flowering plants put together by the Angiosperm Phylogeny Group 2 years earlier. He then constructed several statistics that measured whether the co-occurring tree species tended to be more clumped in the phylogeny or more overdispersed. He computed these statistics for the plot samples and compared his results with a null model of a community randomly assembled from the species pool. He found that the samples were phylogenetically clustered, suggesting that trees species assemblages are organized through habitat specialization and that interspecific competition plays a minor role during community assembly. This approach has been influential in community ecology over the past few years (see Webb et al., 2002). The neutral theory makes the prediction of neither undernor overdispersion in the co-occurring species. However, because species assemblages are dispersal limited, their topology differs from random trees.
Returning to MacArthur’s original remark that it is difficult to see all at once the “machinery” and the “history” of a phenomenon, ecologists now agree that both viewpoints are valuable, depending on the question being addressed. Quite likely, the recent development of the neutral theory of biodiversity has helped reconcile these viewpoints. In some cases, deterministic interpretations of a phenomenon hold valuable knowledge, and the hope of the ecologist is that, having unveiled this causal link, she will be able to make robust predictions. In other cases, the interpretation of a phenomenon cannot ignore historical contingencies. Indeed, for most complex systems, we know that it is illusory to search for a fully mechanistic interpretation of a phenomenon that will also give rise to robust and long-term predictions. Although this has been known by meteorologists for almost five decades, it has not discouraged them from constructing increasingly sophisticated methods of weather forecasting.
During decades of debate over the processes that control the organization of ecological communities, the prevailing view has matured. The original dogma that competition was the only mechanism able to explain patterns of community organization has drifted to tests including stochastic processes. As a result, recent studies tend to be considerably more mathematically sophisticated than those in the past. This reflects the fact that more ecological data, and also more computer power, are available today. Researchers have also opened the door to new approaches that not only are inspired by evolutionary theory but make full use of this body of knowledge. Evolution questions are historical in nature, and they call for a proper historical framework, for instance, a robust phylogenetic hypothesis. Although it has long been remarked that ecological questions should be properly interpretable in light of evolution, it is only with the advent of more easily accessible methods in molecular biology that this idea has swept through the field of ecology. Meanwhile, classic studies in community ecology also benefited from more interaction with physiologists. They gained from serious attempts to develop synthetic analyses involving field studies and theory as well. On the modeling side, much progress remains ahead of us, as increasingly complex objects (phylogenies, large-scale trait databases, regional species distribution) are being assembled for testing ecological hypotheses.
Alonso, David, Rampal S. Etienne, and Alan J. McKane. 2006. The merits of neutral theory. Trends in Ecology and Evolution 21: 351–356. This article summarizes the mathematical foundations of the neutral theory of biodiversity.
Dayan, Tamar, and Daniel Simberloff. 2005. Ecological and community-wide character displacement: The next generation. Ecology Letters 8: 875–894. This article provides a comprehensive overview of 50 years of research on the consequences of competition in closely related species.
Gause, Georgyi F. 1934. The Struggle for Existence. Baltimore: Williams & Wilkins. Available online at http://www.ggause.com/Contgau.htm. This is a foundation of model testing and experimental methods in ecology. This book paved the way for much theoretical ecology until today.
Gotelli, Nicholas J., and Gary R. Graves. 1996. Null Models in Ecology. Washington, DC: Smithsonian Institution Press. This book reviews many of the important patterns of community ecology as well as tests of ecological theories using these patterns.
Hubbell, S.P. 2001. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, NJ: Princeton University Press. In only a few years, this book has become a classic in ecology. It provides a new perspective on patterns of community organization and offers a theoretical interpretation of these patterns that is an alternative to classic niche theory.
Hutchinson, G. Evelyn. 1959. Homage to Santa Rosalia or why are there so many kinds of animals? American Naturalist 93: 145–159. This is a classic in community ecology that formulates an interpretation for body size differences among closely related species.
Kingsland, Sharon E. 1995. Modeling Nature: Episodes in the History of Population Ecology, 2nd ed. Chicago: University of Chicago Press. This wonderful book provides a concise history of key concepts in ecology and of the lives of those who forged them.
Losos, Jonathan A. 1990. A phylogenetic analysis of character displacement in Caribbean Anolis lizards. Evolution 44: 558–569. This presents the first phylogenetically controlled test of Hutchinson body size ratio. This paper provided a much needed new light on an old heated debate in ecology.
MacArthur, Robert H. 1972. Geographical Ecology: Patterns in the Distribution of Species. Princeton, NJ: Princeton University Press. This is one of the tipping points in the history of ecology. It is a deep and easily accessible text for general readers.
McGill, Brian J., Brian A. Maurer, and Michael D. Weiser. 2006. Empirical evaluation of neutral theory. Ecology 87: 1411–1423. In this article, the authors provide an overview of several tests of the neutral theory of biodiversity. This is an excellent introduction to the recent literature on this topic, although some of their conclusions are still a matter of debate and should therefore be taken with a grain of salt.
Schluter, Dolph. 2001. Ecology and the origin of species. Trends in Ecology and Evolution 16: 372–380. This easily accessible overview of speciation theory summarizes the different modes of speciation and evidence for them. See also Jerry Coyne and Allen E. Orr’s book on speciation.
Webb, Campbell O., David D. Ackerly, Mark A. McPeek, and Michael J. Donoghue. 2002. Phylogenies and community ecology. Annual Review of Ecology and Systematics 33: 475–505. This is the best overview to date of how the dialogue between phylogeny and ecology may lead to progress in both disciplines.