1. What are “top-down” and “bottom-up” processes?
2. A history of converging views
5. Moving beyond vignettes: Empirical generality and tests of theory
In this chapter we briefly trace the historical debate and outline the theoretical and empirical evidence for factors controlling the biomass of predators, herbivores, and plants within and among ecosystems.
autotroph. Organisms that make their own food by synthesizing organic compounds from inorganic chemicals, usually via photosynthesis (e.g., algae, vascular plants).
biomass. The total mass of living biological material.
consumer. See heterotroph.
food web. Network of feeding relationships among organisms in a local community.
heterotroph. Organisms that must consume organic compounds as food for growth (e.g., animals, most bacteria, and fungi).
primary producer. See autotroph.
trophic. From Greek, “food,” this term refers to feeding of one species on another, as in “trophic interactions” or “trophic links.”
trophic level. Feeding position in a food chain: autotrophs form the basal trophic level, herbivores represent the second trophic level, and so on.
Humans are dramatically altering the global budgets of elemental nutrients that limit the growth and biomass of autotrophs, or primary producers. Through activities such as fossil fuel combustion and application of agricultural fertilizers, global pools of nitrogen and phosphorus have doubled and quintupled, respectively, relative to preindustrial levels. The impacts of these nutrient fertility bonanzas are most obvious in surface waters of lakes and coasts. Nutrient eutrophication often causes rapid and explosive blooms of algae and microorganisms and equally rapid death, decomposition, and ecosystem-wide oxygen starvation, or hypoxia. The Gulf of Mexico hypoxic “dead zone” at the mouth of the Mississippi River annually swells over areas exceeding 18,000 km2, larger than the U.S. state of Connecticut. Nutrient eutrophication is a jarring example of a bottom-up process, resource supply, that can dramatically alter autotrophs and the food webs that rely on them for energy and nutrition.
Concurrently, humans are changing the role and composition of consumers in food webs via species removals and additions. Habitat loss and degradation and selective hunting and fishing deplete consumers disproportionately from food webs; many top predators such as tigers, wild dogs, wolves, and sharks have been hunted to near ecological extinction. At the same time, humans are adding consumers to food webs for endpoints such as conservation, recreation, and agriculture as well as accidentally introducing invasive consumer species. In a dramatic example, the brown tree snake (Boiga irregularis), a nocturnal predator, was accidentally introduced to Guam after World War II. This single species has eaten its way through Guam’s native food web, causing direct reductions or complete extinctions of dozens of native birds, bats, and reptiles, and indirect negative impacts to native arthropods, forest tree seed dispersal, and recruitment. This example highlights an extreme change in top-down processes, or consumption of organic biomass, that can have dramatic effects throughout food webs.
Management of algal blooms, crop fertilization, agricultural insecticide use, and wildlife conservation are prime examples in which complex interactions between bottom-up processes (i.e., fertility) and top-down processes (i.e., consumption) challenge us to better understand the critical processes that bridge communities and ecosystems. Thus, understanding the ways in which altered resources and consumer community structure interact to control the biomass of predators, herbivores, and plants is not simply a problem for basic science but one that has an immediate impact on humans. Biological control of crop pests and control of lake clarity are two management realms that draw on knowledge about these interacting processes to bring about planned changes in whole ecosystems.
Thousands of scholarly studies report on the implications of fertility manipulations and biological weed or pest control introductions for applied endpoints such as agricultural yield. We focus here on the basic science underlying such bottom-up and top-down applications. Although such factors as genetics, disease, nutrition, dispersal, and spatial structure can be critically important in structuring communities, we focus primarily on fertility and consumer controls of communities, as even this more restricted literature is quite vast. We refer to an extremely simplified theoretical community, or “module,” describing one predator, one herbivore, one plant, and one nonbiological resource (e.g., nitrogen; figure 1). Most common mathematical descriptions of this module treat each level as a single species; however, these “species” often are conceptualized as unified “trophic levels,” each containing multiple interacting species. We will provide empirical examples of strong top-down and bottom-up control and examine evidence across the literature for whether these are special cases or represent general patterns in ecosystems. We will end by outlining a few of the most fruitful future directions for this vibrant and rapidly progressing field of community ecology.
First, let us not lose sight of the forest for the trees. Terrestrial biomes (e.g., tropical savannah, desert, arctic tundra) are defined by their dominant plants, which, in turn, grow in these regions primarily because of the regional combination of solar radiation, temperature, and precipitation. Aquatic systems have parallel, broadly defined regions (e.g., kelp forest, coral reef), with their location determined, in large part, by the regional combination of light availability and temperature. These broadly defined regions in both aquatic and terrestrial systems tend to have characteristic nutrient availability, but even the most nitrogen-rich oceans contain only about 1/10,000 the nitrogen of topsoil. At regional and local scales in all systems, nutrient supply and consumption may be virtually irrelevant for determining community biomass in habitats with extreme physical disturbance, such as those exposed to heavy surf or volcanic activity.
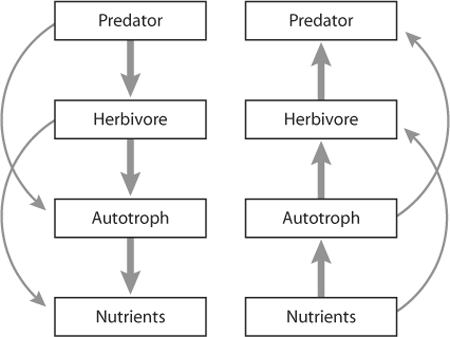
Figure 1. Community modules illustrate top-down and bottom-up direct and indirect interactions. In the first scenario, direct consumption (thick arrows) leads to indirect increases or reductions (thin arrows) in lower trophic levels and resource pools (e.g., soil nitrate). This scenario represents top-down consumer control of a community. In the second scenario, exemplifying bottom-up community regulation, fertilization directly and indirectly alters the abundance of autotrophs and consumers. These modules may define an entire community with only a single species at each level, may be composed of several species interacting within each level (e.g., several herbivore or plant species), or may be part of a larger, more complex food web.
In addition, although herbivores such as sea urchins, locusts, or rabbits can decimate plant biomass in some circumstances, as a group, herbivores consume less than an estimated 20% of annual terrestrial plant production, less than 40% of ocean production, and approximately 50% of freshwater production. Although herbivores such as crop pests can consume large proportions of crop biomass, herbivores substantially control the biomass of all producers in a region in only infrequent and notable cases. Thus, consumers can be important drivers of producer biomass patterns only if producers have sufficient light, precipitation, and nutrients, and physical disturbance is not extreme or frequent. Given this global context, in this chapter we step a bit closer to examine the roles of altered fertility and consumption within communities.
Our current understanding of the interactive effects of fertility and consumption in controlling the relative abundance of plants and consumers rests on the refinement of historical debates. As in most fields of research, this debate began with two opposing views. Both views were well supported by empirical evidence, and both had grounding in mathematical theory.
Charles Elton first proposed a “pyramid of numbers” in which primary producers dominate and consumer biomass decreases as trophic levels become more remote from the base of production. This generality seems to agree with our observations of terrestrial systems, but aquatic ecosystems often violate Elton’s rule with inverted biomass pyramids, or ratios of heterotroph-to-autotroph biomass (H:A) greater than 1. By observing successional transitions in producer quality from lakes to bogs to terrestrial communities, Raymond Lindeman reconciled this aquatic-terrestrial contrast by hypothesizing systematic and taxonomic differences in trophic conversion and assimilation efficiencies. This hypothesis explained both the increasing domination by plants in terrestrial habitats and, ultimately, the limitation of energy reaching the top consumers across all food webs.
This bottom-up view largely prevailed until Hairston, Smith, and Slobodkin (HSS) introduced the classic top-down alternative, that predators protect the “green world” from rabbits and sea urchins by regulating their densities below outbreak levels that could decimate producers. Robert Paine later coined the term trophic cascade, as an indirect effect of predators on plant biomass via consumption of herbivores, to describe this phenomenon. Numerous empirical examples, notably from aquatic or relatively simple terrestrial systems (e.g., monoculture crops), confirmed that experimental removal of predators could cause reductions in plants via increased herbivory.
Early criticism of HSS noted that primary producers are neither uniformly edible nor immediately available to consumers. Physical attributes, such as spines, trichomes, or tough and thickened leaf tissues, as well as a variety of constitutive or rapidly inducible chemical compounds, protect producers from herbivore consumption. A body of optimality theory predicted (1) that plants should develop adaptive defenses in proportion to the risk of herbivore consumption and (2) that available resources should constrain plant defenses, with trade-offs to other plant functions such as growth or reproduction. Some authors argued forcefully that consumers were irrelevant in tropical and temperate forests, deserts, and many other terrestrial habitats. Traits that confer strong competitive ability for limited resources on land (e.g., woody stems to grow for light) also reduce consumption and assimilation of plant biomass to herbivores and higher trophic levels.
With theoretical advances and increasing empirical data, the dialectic of top-down versus bottom-up forces yielded to a nuanced view acknowledging the dual role of these pressures within communities. Oksanen and colleagues used a simple mathematical model to propose that the total community biomass, number of trophic levels, and strength of top-down pressure in food webs should depend ultimately on the productivity of a system, thus melding top-down and bottom-up paradigms into a single hypothesis (see box 1). This hypothesis added the twist that food chain length can predict whether predators should have positive or negative indirect effects on producers. In three-level examples envisioned by HSS, predator presence should increase plant production relative to the same community lacking predators. In four-level food chains, the top predator releases herbivores indirectly by consuming third-level predators, thereby indirectly controlling producers (box 1). This theory predicts that the effects of productivity and predation should remain strong across all trophic links; however, Menge and Sutherland suggested an alternative model in which the effects of nutrients and predation attenuate as they travel through a food web. In this case, resource supply and species competition should most strongly control autotrophs, but at higher trophic levels, predation should increase in its controlling effect.
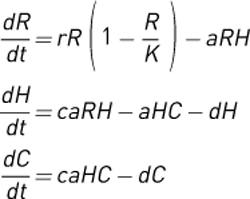
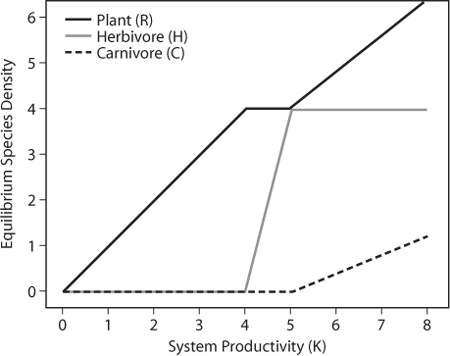
Box Figure 1. This simple mathematical model of a plant resource (R), an intermediate herbivore (H), and a top carnivore (C) produces a variety of predictions across a gradient of increasing productivity, including increased species richness, increased numbers of trophic levels, and changing relative abundances of each species.
These equations provide a dynamic mathematical description of an extremely simplified linear trophic web with a plant resource [R], an intermediate herbivore [H], and a top carnivore [C]. Here, the plant increases via logistic growth to a fixed carrying capacity [K]. Plants and herbivores are consumed only by the adjacent trophic level; there is no omnivory in this description. For simplicity, the attack [a], conversion efficiency [c], and death [d] rates are the same for both consumers. These equations allow us to make predictions for a community module with this structure at equilibrium with increasing system carrying capacity, K. The plot in box figure 1 shows the equilibrium predictions for this community module when r = 10, a = 0.5, c = 0.5, and d = 1.
System productivity controls total community biomass and the number of trophic levels, whereas consumption controls the distribution of biomass among trophic levels. Higher trophic levels can easily be added to this mathematical description; communities with an odd number of trophic levels (e.g., three: plant, herbivore, and predator) are predicted to release plants, and those with an even number of trophic levels (e.g., four: plant, herbivore, intermediate predator, and top predator) suppress plant growth.
The classic paradigm of a top-down trophic cascade is exemplified by lake systems with phytoplankton as the dominant primary producers. Phytoplankton are typically grazed by zooplankton (e.g., Daphnia spp.), which are consumed by small planktivorous fish (e.g., minnows). Whole-lake manipulations of piscivorous fish (e.g., bass) have shown that three-level chains lacking piscivorous fish result in green lakes because planktivorous fish (third trophic level) limit zooplankton (second level) and release phytoplankton (primary producers), whereas blue lakes result where piscivorous fish are added as the fourth trophic level. A great number of empirical examples show similar top-down scenarios from lake systems, and an entire class of restoration techniques focuses on the removal or addition of trophic levels (“biomanipulation”) to achieve desired states of primary productivity and water clarity in lakes.
A rare example of a terrestrial top-down trophic cascade comes from Isle Royale National Park, Michigan. Time series data revealed that the width of annual tree growth rings on balsam fir (Abies balsamea) correlated positively with the annual abundance of wolves (Canis lupus) and negatively with moose (Alces alces), the prey of wolves and the dominant browsing herbivore on the island. This example also demonstrates that pathogens or parasites may act as effective consumers in food webs. An outbreak of canine parvovirus decimated the wolf population in the 1990s, releasing the moose population from top-down control, again in the manner expected from theory for four-trophic-level chains (figure 2A). Controversy for years raged over whether terrestrial trophic cascades were empirically rare because they are less prevalent and powerful than those in aquatic systems or because they are simply more difficult to study at appropriate scales of time and space.
Top-down and bottom-up forces can shift in their relative importance over large spatial and temporal scales, as exemplified by studies from several marine communities. Along the Pacific coast of the Americas, kelp forests dominate hard substrates in the shallow subtidal zone, providing resources and habitat structure for a complex food web of microbes, algae, invertebrates, fish, and mammals. Seminal work from Alaska has shown that sea otters (Enhydra lutris) consume invertebrate grazers, such as sea urchins, which in the absence of otters can completely eliminate kelp forests and shift the community to an alternative state (“urchin barren”). Historical declines in large marine mammals recently have induced orca whales (Orcinus orca) to increase their predation intensity on otters, thus reducing the protective role of otters for kelp in a manner predicted by food chain theory (figure 2B; box 1). Geographic variation in seawater temperature may ultimately drive the relative importance of top-down and bottom-up forces in this kelp forest system. In the cold, nutrient-rich waters of Alaska, the nitrate needed for kelp growth is seldom limiting, and predators instead control food web dynamics. In the warmer waters along California’s southern coast, by contrast, otters apparently play a diminished role, and nitrate concentrations decline precipitously unless replenished by upwelling events that churn nutrients off the sea floor. Warm El Niño-Southern Oscillation (ENSO; coupled oceanic and atmospheric phenomena in the Pacific Ocean causing altered sea surface temperature, winds, and rainfall) events further intensify the nutrient limitation of kelps and can cause forest diebacks on broad scales. A similar pattern of geographic variation has been shown on the Northern Atlantic shelf, where food webs in colder waters were more sensitive to heavy exploitation of cod—and were more likely to be controlled by top-down trophic cascades—than the more resilient, species-rich food webs in warmer subtropical, nutrient-limited waters.
Figure 2. Data plots from two studies demonstrating the relative importance of top-down and bottom-up factors in regulating real communities. (A) In a three-level community, moose populations are controlled primarily by predation with some influence of resource competition, whereas after a viral outbreak created a four-level community and virtually removed the wolf population, moose were more strongly controlled by resource availability. (Reprinted from Wilmers, C. C., E. Post, R. 0. Peterson, and J. A. Vucetich. 2006. Predator disease out-break modulates top-down, bottom-up and climatic effects on herbivore population dynamics. Ecology Letters 9: 383–389) (B) Evidence for both three- and four-level top-down control. (Reprinted from Estes, J. A., M. T. Tinker, T. M. Williams, and D. F. Doak. 1998. Killer whale predation on sea otters linking oceanic and nearshore processes. Science 282: 473–476) In the 1980s, sea otter (a) and kelp (d) biomass was high, whereas sea urchin biomass (b) was low. In contrast, in the late 1990s, when otter abundance declined, urchins reached high biomass, and kelp was rare.
A relatively simple mathematical description of single-species interactions (box 1) incorporates both system fertility and consumptive interactions. This mathematical description predicts that, at equilibrium, predators should benefit plants indirectly via a trophic cascade. Thus, communities lacking predators should have less plant biomass and more herbivore biomass than a similar community with predators present. But this same mathematical description also predicts that alterations of system fertility or productivity should affect all trophic levels of the community such that more enriched communities should contain relatively more predator biomass than comparable, but less enriched, communities. This mathematical description allows us to develop logical, if quite general, hypotheses about the resulting biomass of plants and herbivores when we examine or manipulate fertility or consumers in real communities.
As born empiricists observing our world, we know that most communities are composed of many species of interacting plants, herbivores, and predators, so this description of a whole community using a three-species module (box 1), with a single predator species specializing on a single herbivore species, which in turn specializes on a single plant with a single limiting resource, offends our empirical sensibilities. Obviously, this mathematical description is an extreme simplification of any natural community. Given its simplicity, it is surprising how often this exceedingly simplified mathematical description matches our observations of communities, as in the lake, Isle Royale, and kelp forest communities described earlier in this chapter (figure 2).
There are many examples of communities in which fertilization and consumer addition or removal do not support the predictions of our simple model. In these cases, complexities of the community appear to either enhance (rarely) or reduce (commonly) the predicted direct and indirect effects of fertility and consumption. Most community ecologists have a pet hypothesis about when and where top-down or bottom-up effects should be most apparent (box 2), and the simple theory presented here (box 1) has been extended in many ways to produce testable predictions. In many cases, a slight addition of complexity can aid our logic and increase our predictive power. For example, modules of single-species consumptive interactions (figure 1) can exist within more complex food webs, but even species-poor communities commonly have several species in each trophic level. In fact, compensation within the plant community caused by a trade-off between abilities to resist herbivory and compete for limited resources tends to reduce both direct and indirect top-down effects. Top-down and bottom-up effects can still interact, even within complex communities; in communities featuring both competition within a trophic level and consumption among levels, top-down effects are predicted to be strongest at low to moderate productivity levels. Similarly, herbivore avoidance of predators or preference among plant species, for example, can cause a variety of community responses to enrichment, including altered composition and biomass of both plants and herbivores.
When is consumption most likely to indirectly affect plant biomass? When should we expect fertility to control community composition? A plethora of hypotheses exist in the literature to address these questions, some based in models, others based in logical arguments. These hypotheses range from biological to methodological, falling into broad categories that in many cases are interlinked:
Spatial heterogeneity. Predators will be ineffective if herbivores can hide. Overall, predators will control the biomass of herbivores and plants in less complex habitats with few herbivore refuges.
Food webs that deviate from a linear food chain. Real communities are often better described as a food web than a food chain. Trophic interactions such as omnivory, competition, or intraguild predation, or behavioral interactions such as avoidance or territoriality, tend to increase interference and reduce the overall impact of each consumer level on the next. Predators should have little control over the biomass of subsequent trophic levels in complex communities, leading to stronger bottom-up control.
Predation risk, nonconsumptive effects, and flexible foraging. At intermediate levels in food chains, herbivores must balance the trade-off between eating and being eaten. Predator cues or the threat of predation may shift the quality and quantity of herbivore dietary intake—effects that can cascade to change plant community composition and ecosystem processes without consumptive changes in herbivore densities.
Species turnover. Changes in nutrient and consumer regimes can precipitate turnover in community species composition. Herbivore pressure can transform plant communities to favor less edible, more defended plants or may select for abundance of tolerant plants that rapidly recover to replace less-tolerant species. These changes may depend on nutrient resource supply.
Communities at equilibrium. When they experimentally examine the effects of predation on herbivore and plant biomass, studies measuring treatment effects before plants have had time to regenerate are likely to measure only the effects of consumption, not the effects of long-term alterations in equilibrium biomass. In addition, because herbivory can lead to a change in the composition of the plant community, short-term studies may produce a biased estimate of the true long-term effects of predator removal on plant and herbivore biomass. Finally, communities subject to frequent or intense disturbance may never reach equilibrium, reducing the importance of top-down or bottom-up controls.
Resource availability and quality. Where plant resources are nutritious and easy to consume, consumption rates by herbivores should also be higher. Thus, herbivores should strongly control the biomass of nutritious plants, and reduction of herbivores by predation should cause greater increases in plant biomass in a community with nutritious plants than in a similar community with less nutritious plants.
Predator or herbivore efficiency. Consumers with low metabolic costs (e.g., many invertebrates) can convert a higher percentage of what they consume into reproduction than can those with higher metabolic costs (e.g., mammals). In general, consumers that are extremely effective at converting food into progeny should effectively control the biomass of their resource, leading to greater control by predators over the biomass of herbivores and plants.
Historically, empiricists have approached ecology with a system-specific mindset, tending to think and work within a single ecosystem type. Ecological generalities, such as the factors determining the relative importance of consumers and fertility, are indeed interesting and important within systems but are not constrained by system boundaries. In fact, examining and manipulating consumers and fertility both within and among ecosystems is a promising avenue for confirming, refuting, or suggesting new hypotheses about when fertility or consumers should control community biomass distribution (see box 2). Clearly an important step in discovering generalities about communities is to quantify the relative importance of consumers and fertility across gradients in fertility and disturbance, species composition, and community complexity as well as among ecological systems. But a single, replicated, manipulative experimental study to successfully accomplish this goal is virtually impossible to envision.
The literature examining single cases of consumer addition or removal under a variety of fertility regimes has grown in the decades since the publication of HSS and spans a broad range of systems, taxa, spatial and temporal scales, and food web complexity. Drawing from this diverse literature, recent quantitative syntheses have examined the emergent evidence for the relative influence of top-down and bottom-up control of communities across numerous empirical case studies. Most of these reviews and meta-analyses have focused on within-system comparisons, but several also have examined the evidence for fertility and consumer controls among systems.
These quantitative analyses of the existing empirical data show a great deal of variation in the relative importance of consumers and fertility in regulating H:A ratios and community composition. However, some consistent patterns emerge from these analyses. Both top-down and bottom-up forces can have substantial food web effects; the direct effects of nutrients and herbivores on plants and predators on herbivores are often quite strong. However, predators can impart strong top-down effects across entire food chains, whereas nutrients primarily affect plant productivity. When responses are compared among systems, plant biomass tends to increase with fertilization in all systems, whereas herbivory consistently suppresses plants in oceans and lakes but has variable effects on land. The indirect effects of predators on plants tend to be strongest in some marine and lake communities and weakest in streams and on land. Overall, the herbivore-plant link is weak; in contrast to the predictions of simple community theory (box 1), herbivores in predator-free communities appear unable to take advantage of increases in ecosystem productivity. Regulation of community biomass by consumers appears to be associated with consumer metabolism, an intriguing finding that deserves further experimental examination across a variety of taxa (see Underrepresented Taxa and Systems in the next section).
After decades of exhaustive theoretical, empirical, and synthetic investigation into the roles of top-down and bottom-up forces in diverse ecosystems, our understanding of the relative strengths of these processes has come a long way since Elton and Lindeman. Even so, there remain many rapidly progressing areas in which major questions remain. Here, we outline a few exceptionally promising avenues for future research.
We thank all our past and current mentors, colleagues, and collaborators who have shaped our thinking on this topic. We also thank Angela Brandt, Matt Bracken, and Michel Loreau, who read and commented on drafts, and Eric Seabloom, who assisted with R coding for box 1.
Borer, E. T., B. S. Halpern, and E. W. Seabloom. 2006. Asymmetry in community regulation: Effects of predators and productivity. Ecology 87: 2813–2820. This quantitative meta-analytic review examines predator removal experiments across productivity gradients and in factorial fertilizations in marine, freshwater, and terrestrial systems.
Gruner, D. S., J. E. Smith, E. W. Seabloom, S. A. Sandin, J. T. Ngai, H. Hillebrand, W. S. Harpole, J. J. Elser, E. E. Cleland, M.E.S. Bracken, E. T. Borer, and B. M. Bolker. 2008. A cross-system synthesis of herbivore and nutrient resource control on producer biomass. Ecology Letters 11: 740–755. This synthesis uses meta-analyses to test predictions from simple mechanistic models and demonstrates strong and independent effects of herbivores and nutrient resources on plant community biomass that are broadly similar in relative magnitude across ecosystem types.
Hairston, N. G., F. E. Smith, and L. B. Slobodkin. 1960. Community structure, population control, and competition. American Naturalist 94: 421–425. This foundational paper argues via a verbal model for top-down control maintaining high plant biomass; often referred to as the Green World Hypothesis.
Hillebrand, H., D. S. Gruner, E. T. Borer, M. E. Bracken, E. E. Cleland, J. J. Elser, W. S. Harpole, J. T. Ngai, E. W. Seabloom, J. B. Shurin, and J. E. Smith. 2007. Community structure and ecosystem productivity mediate the intrinsic control of producer diversity across major ecosystem types. Proceedings of the National Academy of Sciences, U.S.A. 104: 10904–10909. This meta-analysis examines results from hundreds of published experiments and demonstrates how top-down and bottom-up forces can interact to change producer composition even when biomass is not affected.
Leibold, M. A., J. M. Chase, J. B. Shurin, and A. L. Downing. 1997. Species turnover and the regulation of trophic structure. Annual Review of Ecology and Systematics 28: 467–494. This is a review of models and evidence for fertility-driven shifts in species composition that alter the strength of trophic interactions.
Lindeman, R. L. 1942. The trophic-dynamic aspect of ecology. Ecology 23: 399–418. This article presents a classic synthesis, published posthumously, that anchored the bottom-up perspective as the dominant paradigm for a generation of ecologists. This paper merged the ecosystem-level ideas of nutrient and energy flux with community-level feeding relationships to explain long-term dynamics of plant community succession.
Matson, P. A., and M. D. Hunter, eds. 1992. Special feature: The relative contributions to top-down and bottom-up forces in population and community ecology. Ecology 73: 723–765. This special feature included four seminal contributions (authored by Hunter and Price, Power, Strong, and Menge) that surged this topic to the forefront of community ecological thinking. All contributions argued for a pluralistic view that embraces the complexity and heterogeneity of real ecosystems and communities.
Menge, B. A., and J. P. Sutherland. 1976. Species diversity gradients: Synthesis of the roles of predation, competition, and temporal heterogeneity. American Naturalist 110: 351–369. This foundational verbal model argues that the relative importance of nutrients and consumption should vary with environmental disturbance: consumption should structure communities that experience low physical disturbance but should be relatively unimportant in physically disturbed habitats.
Oksanen, L., S. D. Fretwell, J. Arruda, and P. Niemela. 1981. Exploitation ecosystems in gradients of primary productivity. American Naturalist 118: 240–261. This article melded the bottom-up and top-down paradigms into a single mathematical framework with predictions for food chain length and strength of trophic cascades.
Polis, G. A. 1999. Why are parts of the world green? Multiple factors control productivity and the distribution of biomass. Oikos 86: 3–15. This article presents an engaging narrative review of broad-scale patterns and case studies showing the multiplicity of potential abiotic and biotic limiting factors that alter the relative strengths of top-down and bottom-up control.
Polis, G. A., and D. R. Strong. 1996. Food web complexity and community dynamics. American Naturalist 147: 813–846. This article presents a critical appraisal of simple food chain models of trophic dynamics that argues that strong indirect trophic interactions in complex food webs are the exception, not the rule.