1. A view of landscapes as functioning systems
2. Spatial heterogeneity as an organizing principle in landscape dynamics
3. Function and dysfunction in landscape dynamics
5. Assessing landscape dynamics: Thresholds and climate change
A terrestrial landscape can be viewed as a system of biological elements (organisms, populations, communities) forming a pattern across a topographic geomorphic unit. The dynamics of these landscape systems are driven by topography and climate and by interacting geochemical and biophysical processes. Although we recognize that important conceptual advances in landscape dynamics have been developed, such as how landscapes behave as complex adaptive and self-organizing systems, in this chapter we particularly focus on the development of the notion of landscape function, that is, how a landscape works as a geochemical-biophysical system to regulate vital resources over space and time. In highly functional landscapes, a major rainfall event will trigger runoff, but, overall, little loss of water, topsoil, and organic matter occurs from the system because these resources are dynamically captured by patterned structures within the landscape such as vegetation patches, which function as reserves or resource “banks.” Vegetation patches then utilize retained resources (water, nutrients) in growth pulses to produce biomass such as seeds, most of which are cycled back into the system (soil seed banks). Biomass can also function to maintain the retentive capacity (structure) of the patch and can provide shelter and food for fauna or for consumption by livestock, which when harvested represent offtake from the system. Damaged landscapes become dysfunctional by losing their capacity to effectively regulate resources.
landscape dynamics. How a landscape, as a system of interacting components, structures, and processes, varies in space and time
landscape function. How a landscape works as a tightly coupled geochemical-biophysical system to regulate the spatial availability and dynamics of resources
landscape heterogeneity. The mix of different components, structures, and processes occurring in a landscape, such as how different organisms disperse among different vegetation patches
landscape restoration/rehabilitation. The actions and processes taken to help damaged landscapes recover toward a specified goal (landform, land use)
landscapes as self-organizing systems. How components, structures, and processes in a landscape dynamically organize to form complex, adaptive, and stable systems
landscape system threshold. A point in the dynamics of a landscape where the system changes to a different state, as, for example, a damaged landscape becomes dysfunctional to the point where available resources no longer support a species
We view landscapes and marvel at the patterning of their interconnected ecosystems and wonder about what dynamic processes have caused these patterns. What have we learned about the dynamics of landscape patterns and processes in recent times? In this chapter we explore new developments in landscape dynamics by building on the work of Turner, Gardner, and O’Neill (2001) and others and by adding our Australian perspective. Disturbance-induced effects on landscapes are described in later chapters in part IV.
The importance of spatial heterogeneity and self-organization for explaining landscape dynamics has become increasingly recognized over the last 25 years as seen in the writings of Kolosa and Pickett (1991) and Rietkerk and others (2002). This recognition has led to studies on landscape function, that is, how a landscape works as a tightly coupled geochemical-biophysical system that regulates the sources and dynamics of energy, water, and nutrient resources (Tongway and Ludwig, 1997). Landscape function and its dynamics in space and time can be resolved into the availability of vital resources, which strongly affect the responses of biota, especially if stressed or disturbed.
Initially, distinctions were made between measured system complexity and functional heterogeneity, which described how ecological entities such as species perceived, related to, and responded to each other. In the last 25 years, fine-scaled ecological processes have been increasingly integrated with broad-scale geographic-geomorphologic studies to better understand overall landscape function. The underlying processes that determine how landscapes function, the need for understanding heterogeneity, and how function is affected by stress and disturbance have been studied in more detail by integrating disciplines (Lovett et al., 2005); this has benefited both the science and the management of landscapes.
A metaphor for how landscapes function as an integration of processes is a gear train (Lavelle et al., 2006). A landscape may be visualized as system of intermeshing gears, with each gear being a distinct ecosystem with its unique structures (composition) and processes (size, speed) but tightly interconnected (meshed) with other ecosystems. Those ecosystems remote from each other (distant gears) only interact through systems and processes connecting them (the meshing of gears between them), and importantly, the overall landscape is not complete as a functional system until the entire gear train is in place. In a sense, the assembling of the gear train reflects the synthesis of landscape ecology at a range of scales over the last two decades.
This gear-train metaphor is intended to highlight the interconnectivity of processes in terms of the structure and functioning of well-functioning landscapes and the importance of maintaining all of the functional links from the very finest to the coarsest scales (from the smallest, fast-spinning gear to the largest, slow-turning gear). Typically, there is a logical progression in examining landscape dynamics, commencing with detecting and characterizing the pattern at an intermediate scale, accounting for pattern in terms of geochemical-biophysical processes and then extending the analysis to both coarser and finer scales to understand the underlying interconnectivity of ecosystem processes to, for example, conserve resources.
Many disciplines have mechanistically studied ecosystems in detail at fine scales (individual cogs on gears) according to the rules of reductionist science. However, broad-scale studies looking at landscape patterns and processes have been largely descriptive and discussed retrospectively because experiments on broader scales are very difficult and expensive to conduct. Recent progress has recognized landscapes as having fundamental and crucial interactions between ecosystems (connectivity between gears) (Shugart, 1998), which are central to progressing knowledge about landscape function and dysfunction. In particular, it is critical to understand how changes in landscape dynamics are explained by the underlying processes at fine scales when landscapes are subject to stress and disturbance (see later chapters in part IV).
Understanding landscape dynamics requires studies on how pattern and process change through time in terms of how geochemical-biophysical processes link organisms to other organisms and their environment. Linking processes at all scales from the microscopic to the regional are crucial to achieving a predictive understanding of landscape dynamics. All useful models of landscape dynamics must deal with interactions across multiple scales yet make use of the basic principles most strongly expressed at each scale. Returning to the gear-train metaphor, no cog, however small, is irrelevant to understanding landscape dynamics. A cog on a gear of any size (scale) damaged by stress or disturbance can be taken as a limit to the capacity of the landscape to function efficiently. Repair may be possible, but only when our understanding of how the gear train works (how the landscape functions as a system) is adequate.
Interpreting changes in landscapes requires an understanding of how geochemical-biophysical processes affect the dynamic availability of vital resources such as water, nutrients, and organic matter (Ludwig and Tongway, 2000), not just changes in species populations or vegetation patterns. Switches to new alternate stability domains (basins of attraction) can be mechanistically explained in terms of resource availability in space and time (Gunderson and Holling, 2001). These include oversupply, supply cutoff, and unexpected synergisms between different resources that result in a critical threshold being crossed.
Noy-Meir, in 1981, was one of the first ecologists to emphasize the importance of the availability of vital resources in three-dimensional space over time. His dynamic models integrated structure, spatial arrangement, and persistence of life forms as being dependent on differential water availability from runoff-runon processes and recognized that rainfall itself was a poor predictor of biological outcomes. Integrating the availability of soil nutrients with water in runoff-runon redistribution processes (Tongway and Ludwig, 1997) helps to explain the landscape patterns generated by Noy-Meir’s three-dimensional dynamic modeling, which also produced sigmoidal or S-shaped curves to reflect how plant production responds to water availability in arid ecosystems.
There is a continuum of scale in processes from the microscopic to regional landscapes. At a micro scale, the functional role of organisms can be characterized into four groups: microflora, micropredators, litter transformers, and ecosystem engineers (Lavelle and Spain, 2001). The activities of these groups affect and control processes such as soil gas exchange, water percolation, and soil aggregate stability, whose influence is easily recognized at larger scales. The aggregate effect of these processes and properties at landscape scale provides a mechanistic interpretation of macroscopic processes such as surface hydrology, nutrient cycling, plant nutrition, and soil erosion, which are properties readily recognized by land managers.
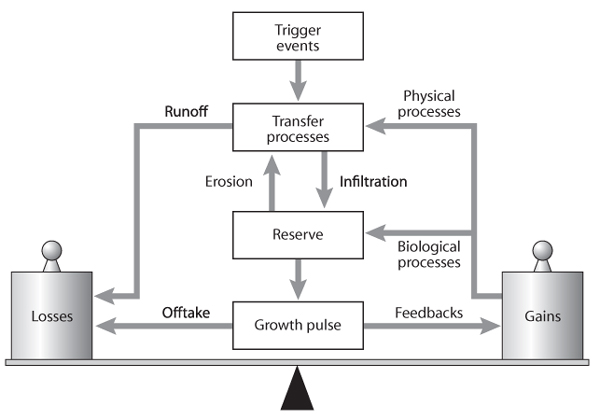
The concepts outlined above are very useful in proposing explanatory frameworks for understanding landscape dynamics. A conceptual framework has been developed that depicts landscape function as a sequence of processes, commencing with rainfall as a trigger (figure 1), which initiates transfer processes such as runoff and erosion that spatially relocate resources such as water, topsoil, organic matter, and seeds across the landscape. Some of these resources may exit the landscape as outflow, and some may be stored in the soil reserve. The reserve may be considered metaphorically as a “bank” dealing in many diverse but interacting ecological “currencies”: water, nutrients, seeds, and soil fauna. Some locations in the landscape, such as runon areas with patches of vegetation on deeper soils, absorb or capture more resources than other parts (interpatch runoff areas). Within vegetation patches, a pulse of plant growth may ensue, the magnitude of which depends on the status of soil moisture and mineralized nutrients (stored reserves). Some of the materials produced by the growth pulse may be lost from the system by fire or herbivory (offtake), but much is cycled back to the reserve by biological feedback where plant litter is reincorporated into the soil by a range of soil biota, and the seed pool is replenished. A physical feedback loop affects the extent to which changes in plant density at ground level regulate the amount and rate at which water and other resources are transported across the landscape. The trigger-transfer-reserve-pulse framework is depicted here as being on a fulcrum, implying that there is a crucial balance between resource losses (outflow plus offtake) and gains (feedback loops) in the landscape system; this balance dynamically fluctuates over time and can be “knocked” out of balance if disturbances damage the system.
Figure 1. The trigger-transfer-reserve-pulse conceptual framework for how geochemical-biophysical processes function in landscapes to dynamically balance resource gains and losses. (After Tongway and Ludwig, 1997)
Landscape function can be represented as a continuum from the least-damaged landscape, called highly functional, to the most-damaged, called highly dysfunctional, landscapes (Ludwig and Tongway, 2000). This continuum is evident in different landscapes such as grasslands and shrublands (plate 6). In highly functional grasslands, a high density of grass plants facilitates the capture, retention, and use of wind- and water-borne resources, whereas in dysfunctional grasslands, these grasses can be replaced by shrubs that fail to trap resources because of low ground contact cover, so that they blow and wash out of the landscape (plate 6, top). High rates of wind and water erosion and low soil nutrient concentrations characterize these dysfunctional landscapes. In contrast, highly functional shrublands effectively trap resources, but dysfunctional shrublands have bare crusted soils that allow water to run off and wind to blow materials away (plate 6, bottom).
In both of these landscapes, landscape function can be defined in terms of the dynamics of water and nutrients at scales varying from the rhizosphere to the local catchment. The grassland versus the shrubland illustrates the different scales at which these landscapes are organized (fine-grained grass tussocks versus coarsegrained shrub clumps), yet both can effectively capture and use vital resources such as water, nutrients, and organic matter. Both grasses and shrubs function above and below ground and provide goods and services (food, shelter) to other biota present.
Reductions in effective retention of vital resources by arid and semiarid landscapes have been characterized as degrees of landscape dysfunction or desertification (Tongway and Ludwig, 2002), and we noted earlier how landscape dynamics can be viewed as changes in the balance between gains and losses of resources in space and time. This concept of resource gains versus losses is very useful when working with highly disturbed sites such as lands affected by mining. If successful, the total development of rehabilitation on a mine site follows an S-shaped curve, which can be partitioned into two components: biological and physical (figure 2). Rehabilitation starts at a low level of natural landscape development (figure 2, point A). Initially this development is largely of the physical component (point B), typically caused by reshaping of landforms (e.g., overburden or spoil heaps; plate 7, photo A) into smoother surfaces that are “ripped” along contours (plate 7, photo B). The aim is to engineer a surface that will capture rainwater and minimize soil erosion. Initially the biological input is very small (figure 2, point C), typically microbial activity in any topsoil applied to surfaces and the initial vegetation establishment (plate 7, photo C). Over time, the vegetation will markedly develop so that biological processes dominate, and the rehabilitated landscape becomes highly developed and functional (figure 2, point D; plate 7, photo D). Although the physical component remains important, it typically declines over time because, for example, riplines flatten.
Figure 2. An example of how the rehabilitation of a landscape develops over time as an S-shaped curve as the combination of two general components: biological and physical processes.
As landscape function increases on rehabilitated mines, biological processes and the accumulation of resources (“natural capital”; Aronson et al., 2007) are improved, as is the capacity of the system to withstand stress and disturbance. This capacity is commonly referred to as the “buffering capacity” or “resilience” of the system (Gunderson and Holling, 2001). Typically, this capacity is described by changes in the biota of a system, but fundamentally system resilience is perhaps better explained by the robustness of interacting processes (viz., the gear-train metaphor).
Mine site rehabilitation provides useful evidence of the initiation and development of feedbacks by establishing plant and soil biota communities, where, after a period, newly acquired resources can be observed to contribute to landscape stability. In particular, plant litter decomposition improves soil aggregate stability by binding soil particles with organic residues: water entry and storage improve, thus augmenting whatever properties were initially provided by engineering. Later, microclimatic conditions created by growing foliage ameliorate the immediate effects of weather and permit other biota to become established. The strengthening of these feedback mechanisms over time can be monitored, as can the “biodiversity” improvement.
As the new landscape develops, its natural capital increases (Aronson et al., 2007), as do the complexity of geochemical and biophysical processes and their interactions. A great deal of attention has been given to the “end game” of the “capital and complexity” response (Gunderson and Holling, 2001), but in heavily impacted landscapes, early information about success in establishing competent resource use is also important to understand, as excessive outflow of resources may prevent vegetation from becoming established. In the gear-train metaphor, a start needs to be made at the scale of the most readily studied scale of pattern and process, extending to both coarser- and finer-scale linked processes.
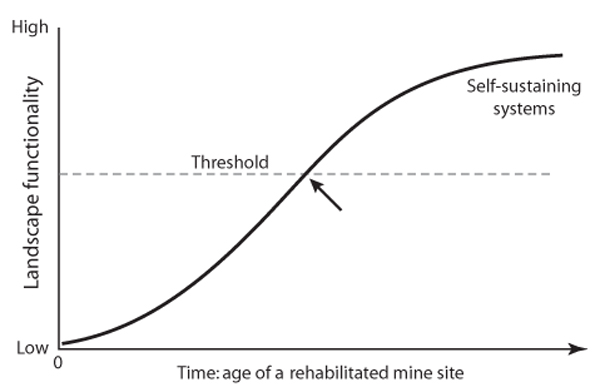
A useful practical concept is that of a critical threshold that marks the point where an ecosystem or landscape is self-sustaining or not. As illustrated in figure 3, as mine site rehabilitation develops over time to be highly functional, it follows an S-shaped curve. At some point along this curve, landscape functionality conceptually crosses a threshold above which the landscape system becomes increasingly self-organizing and self-sustaining because of highly functional geochemical-biophysical processes.
Process-based explanations of crossing such thresholds are more useful than biota-based explanations because of delays in some biotic responses. A critical threshold is crossed when a small change in a landscape driver results in a marked system response. In practical terms, operating a system close to the critical threshold raises the potential for a substantive change. Hence, it is important to understand the factors contributing to the system’s “buffering capacity” in the vicinity of the threshold and to their mutual interactions. For example, allowing a landscape to continue to be grazed by livestock as drought conditions persist or worsen may result in surface conditions that massively erode the landscape when drought-breaking rains fall. Factors such as soil organic matter loss and exposure of unstable subsoils predispose the landscape to damage.
Figure 3. An S-shaped curve describing how mine site restoration typically progresses over time. At some point in time (arrow) a threshold is crossed (dashed line) when landscape functionality exceeds the level required for the site to become self-sustaining.
Because the proposed framework for landscape dynamics has climate and weather as major driver-triggering responses (see figure 1), landscape responses to modeled scenarios of climate can be addressed directly in process terms. Crucial questions proposed by Pounds and Puschendorf (2004) include: How quickly can ecosystems respond to changing climate, especially amount and seasonality of rainfall? Will there be a critical period of adjustment and uncertainty, or will there be a smooth transition to new scenarios?
We have described landscape dynamics using a process-based approach in which geochemical-biophysical processes interact to regulate the economy (gains minus losses) or availability of vital resources. By understanding how landscapes function to conserve resources in terms of spatial patterns and dynamic processes, a diverse range of landscape problems and land management issues can be addressed. Here we used a mine site rehabilitation example, but the principles of landscape function can be applied to a wide range of biomes and land uses such as grasslands and savannas grazed by livestock.
Aronson, James, Suzanne J. Milton, and James N. Blignaut, eds. 2007. Restoring Natural Capital: Science, Business and Practice. Washington DC: Island Press.
Gunderson, Lance H., and C. S. Holling, eds. 2001. Panarchy: Understanding Transformations in Human and Natural Systems. Washington DC: Island Press.
Kolasa, J., and S.T.A. Pickett. 1991. Ecological Heterogeneity. New York: Springer-Verlag.
Lavelle, Patrick, T. Decaens, M. Aubert, S. Barot, M. Blouin, F. Bureau, P. Margerie, P. Mora, and J.-P. Rossi. 2006. Soil invertebrates and ecosystem services. European Journal of Soil Biology 42: S3-S15.
Lavelle, Patrick, and Alister V. Spain. 2001. Soil Ecology. Dordrecht, The Netherlands: Kluwer Academic Publishers.
Lovett, Gary M., Monica G. Turner, Clive G. Jones, and Kathleen C. Weathers, eds. 2005. Ecosystem Function in Heterogeneous Landscapes. New York: Springer-Verlag.
Ludwig, John A., and David J. Tongway. 2000. Viewing rangelands as landscape systems. In O. Arnalds and S. Archer, eds., Rangeland Desertification. Dordrecht, The Netherlands: Kluwer Academic Publishers, 39–52.
Noy-Meir, I. 1981. Spatial effects in modelling of arid ecosystems. In David W. Goodall and Ray A. Perry, eds., Arid Land Ecosystems: Structure, Functioning and Management, vol. 2. Cambridge, UK: Cambridge University Press, 411–432.
Pounds, J. A., and R. Puschendorf. 2004. Ecology: Clouded futures: Global warming is altering the distribution and abundance of plant and animal species. Nature 427: 107–109.
Rietkerk, Max, M. C. Boerlijst, F. van Langevelde, R. HilleRisLambers, J. Van de Koppel, L. Kumar, H.H.Y. Prins, and A. M. de Roos. 2002. Self organisation of vegetation in arid ecosystems. American Naturalist 160: 524–530.
Shugart, Hank H. 1998. Terrestrial Ecosystems in Changing Environments. Cambridge, UK: Cambridge University Press.
Tongway, David J., and John A. Ludwig. 1997. The Conservation of Water and Nutrients within Landscapes. In John A. Ludwig, David J. Tongway, David O. Freudenberger, James C. Noble, and Ken C. Hodgkinson, eds., Landscape Ecology, Function and Management. Melbourne, Australia: CSIRO Publishing, 13–22.
Tongway, David J., and John A. Ludwig. 2002. Reversing Desertification. In Rattan Lal, ed., Encyclopaedia of Soil Science. New York: Marcel Dekker, 343–345.
Turner, Monica G., Robert H. Gardner, and Robert V. O’Neill. 2001. Landscape Ecology in Theory and Practice: Pattern and Process. New York: Springer-Verlag.