1. Feedbacks of landscapes on the atmosphere: Theory
2. Observing feedbacks—comparing different sites
4. The nocturnal boundary layer
This chapter investigates some of the ways in which a vegetated landscape can influence its own climate within the planetary boundary layer. During the daytime, impacts may be exerted up to 1–2 km above the surface and are caused by changes in energy exchange, predominantly evapotranspiration, such as that resulting from deforestation or leafing out in deciduous forests. At night, it has been shown that desert vegetation can significantly warm the air above, probably by the emission of greenhouse-active hydrocarbons. A major problem exists in identifying cause and effect in the recorded changes of the planetary boundary layer, as there appears to be no certainty that the landscape is causing all of the described effects, especially where climate is subjected to chaotic dynamics of varying origins. This feature is also true for climatic change studies at the global scale, where it is difficult to identify with certainty the causes of current climatic changes.
evapotranspiration. The evaporation of water vapor from surfaces plus the evaporation of water through the plant and leaf stomata by transpiration
inversion layer. The cap of the planetary boundary layer, where there is little or no vertical mixing and where the temperature may increase or remain constant
latent energy exchange. The exchange of energy by the evaporation of water
planetary boundary layer (PBL). The lowest part of the atmosphere where the surface influences wind movements, humidity, and temperature over time periods of about 1 hour and up to 1 to 2 km above the surface
sensible heat exchange. The exchange of energy as heat
At the scale of a few meters, it is quite possible to experience feedbacks between the biosphere and the atmosphere. Under bright summer sunshine the air moving past your legs will be warmer if it has moved across dark dry sand than if it has passed through vegetation, primarily because of differences in radiation absorption and evapotranspiration. At the global scale, modifications of carbon dioxide exchange by the terrestrial and marine biospheres can change the global climate. Landscape-scale interactions are somewhere between these two extremes of scale, in the range of 10 to about 20,000 km2.
The relevant part of the atmosphere that interacts with the landscape scale defined above is the lower part of the troposphere, the planetary boundary layer (PBL). This typically extends up to 2 to 3 km above the surface during the day. Convective exchange by buoyant thermals and mechanical turbulence by wind transport lead to energy and mass being exchanged between the surface of the biosphere and the PBL. At this time the surface is warmer than the air above. The top of the PBL is defined by sudden changes in temperature and decreases in specific humidity between it and the free atmosphere above. This inversion layer acts as a cap to the PBL and may incorporate clouds. The PBL grows in depth during the day, driven by increases in solar radiation, which warm the surface and lower layers of the PBL, increasing turbulence and entraining air from the free atmosphere above. At around sunset, thermals cease to form as solar radiation reaches zero, and the PBL collapses to form the nocturnal boundary layer (NBL). At this time, the surface generally becomes colder than the air above, and a shallow inversion layer tens to hundreds of meters in depth forms above the landscape.
Plants in the landscape have the potential to influence the PBL both day and night, with active water and energy exchange during the day exerting influences on PBL development. Topography can also exert a marked and obvious effect, and valley bottoms may often be shrouded in cloud at sunrise. This results from the drainage of cold and dense air into the valley bottom; this layer underlies warmer air above, and water condensation occurs where the warmer air is cooled to dew point temperature.
The PBL, therefore, is the part of the atmosphere that is influenced in some way by the underlying landscape. In the typically heterogeneous landscapes on Earth, lateral wind movements combine with the convective upward and downward exchanges in the PBL (figure 1). The PBL consists of several layers that are affected to different extents by the surface. The canopy layer is directly influenced by the height of the vegetation and by exchanges of energy and mass with the vegetation in a way that is directly analogous to the example mentioned above of walking over dark sand or vegetation. The effects are noticeable within the canopy layer but are rapidly diminished by mixing after the air has moved over the canopy. The wake layer above is mixed by interactions between the mean wind flow and backward movements of air derived from the drag exerted by the canopy on the air flow. The surface layer above averages out the effects occurring in the canopy and wake layers, and these effects are averaged further into the mixed layer extending to the top of the PBL.
The question addressed here is how much effect does the landscape exert on the PBL above, given the extensive mixing that occurs? Observations made at meteorological stations will include these feedbacks of interest but only as a small component of the regional (synoptic) weather and climatic conditions. Two approaches can be used to identify feedbacks: one compares adjacent sites differing in some landscape characteristic, such as differences in vegetation type; the other depends on correlating time series of climate or weather with changing events, such as vegetation phenology, in the landscape.
Vegetation type and climate both interact in determining the daily rate of growth and the temperature and humidity of the PBL. A Brazilian tropical forest had similar heights and rates of growth of the PBL in both the wet and dry seasons (figure 2). By contrast, a nearby pasture, with shallow access to water in the soil, had very different PBL heights. In the dry season, the PBL over the pasture was higher than that over the forest because less water was available for evapotranspiration, which caused more solar radiation to be converted instead to convection. The forest has similar convective heat exchanges in both the wet and dry seasons, indicating that damper soils that are sheltered from the drying sun by the tree canopy account for the uniformly high convective heat exchange from the tall forest. In the wet season, the PBL was shallower over the pasture because the convective heat exchange was smaller than in the dry season, with greater heat loss by evapotranspiration. Energy exchange by evapotranspiration exerts a much smaller effect than convection on daily PBL growth. The greater dry season evapotranspiration rate from the forest exerts a significant impact on the PBL, which is cooler (–1 to –2°C) and more humid (+10%) than it is over the pasture.
Figure 1. Diagram of the planetary boundary layer and its component layers.
Figure 2. Change in planetary boundary layer height for tropical forest and pasture during the wet and dry seasons.
The boundary between native forest and deforested pasture can itself lead to atmospheric feedbacks. In Rondonia, Brazil, the boundary develops increased cloudiness as a result of the production of humid forest breezes that interact with the warmer and drier air over the pasture. The increased convection and strong thermals drive the more humid air upward in the PBL, leading to cloud formation as cooling occurs toward the cap of the PBL.
The potential for changes in landscape evapotranspiration to impact the PBL and climate has been investigated in the state of Nebraska. In this part of the Great Plains, increasing areas of native grassland have been replaced by crops over the twentieth century. In addition, much of the area of crops has also been changed from nonirrigated to irrigated, with anything up to a 40% increase in evapotranspiration, which would be expected to produce feedback on the PBL, as for the case of the pasture and forest.
Table 1. Decadal twentieth-century changes in growing season temperature (°C) over irrigated and nonirrigated areas of Nebraska

Different trends in growing season climate were observed over the irrigated and nonirrigated areas (table 1). Mean, maximum, and minimum growing season temperatures increased where there was no irrigation. The mean minimum temperature of the growing season increased with irrigation, whereas the mean and maximum temperatures decreased. Greater rates of evapotranspiration decrease convective heat loss and cool the vegetation during the day.
The idea that forests control and enhance rainfall has been believed since at least the time of Christopher Columbus, who based this bold assertion on observations that the formerly wooded islands of the Azores and Canaries became desertified after deforestation by the Portuguese. More current observations indicate that anything from 10% to 70% of the water vapor in the atmosphere comes from evapotranspiration. The general connection between deforestation and precipitation is clear; however, the relationship between the extent of deforestation and the impact on precipitation is less clear.
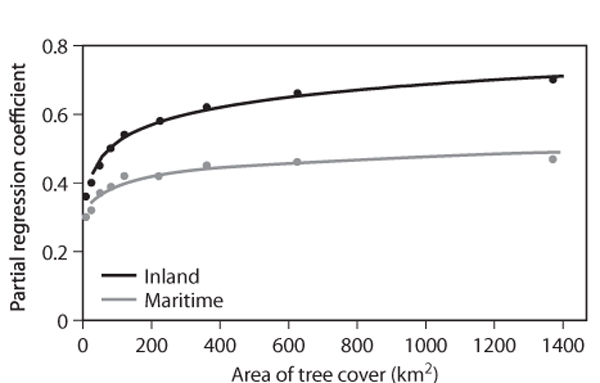
The Atlantic forest of eastern Brazil is a biodiversity hot spot but has suffered significant loss since European settlement. In São Paulo state, forest cover was about 80% before settlement and is now as low as about 10%, with much of the remaining forest in fragments of differing size. With use of remote sensing and detailed analyses of climate records, it has proved possible to demonstrate that there exists a positive correlation between the number of rain days and the spatial extent of the remaining forest fragments (figure 3). Increasing the forest cover to an area of about 200 km2 exerts a positive and significant impact on days of rain and precipitation. The increase then slows but is still positive as the area of tree cover increases to 1400 km2. It is also interesting to note that the correlation is significantly less for coastal forests, and this may reflect a combination of specific maritime climatic features; in particular, the sea breeze circulation and the high humidity of maritime air are relatively little influenced by landscape evapotranspiration.
Figure 3. The correlation between area of forest fragments and rain days in Brazil for inland and maritime areas. An increasing coefficient indicates a closer correspondence between forest cover and rain days.
Cold temperate forest landscapes have a predictable period of leafing out in the spring. Such an event provides a clear step change in the vegetation activity within the landscape, with an expectation that the onset of transpiration will lead to some humidification of the PBL. Long-term measurements of energy fluxes, climate, and vegetation phenology have been available for a range of sites globally and can provide data to test and quantify the humidification proposal. One such site is the Harvard Forest Long-Term Ecological Research Site of Harvard University (http://harvardforest.fas.harvard.edu). Flux data from the site and others are available at http://cdiac.esd.ornl.gov/programs/ameriflux/data_system/aamer.html.
Harvard Forest is located at Petersham, in the state of Massachusetts, which has about 60% cover as forest and therefore a potential to influence PBL activity and composition. Daily averages of energy fluxes for the period from 1994 to 2000 (figure 4) show a peak of net radiation (incoming less outgoing fluxes of solar and long-wave radiation) in about early to midsummer, but with different trends in sensible (convective) and latent heat (evapotranspiration) transfer. Low vegetation activity during the winter months leads to limited latent heat transfer, and convective transfer dominates. Convective transfer declines quite abruptly from day 160, whereas latent heat transfer increases up to midsummer, peaking at the same time as net radiation.
Maximum and minimum temperature and humidity (figure 5) also peak in midsummer. It is interesting to note that the completely frost-free period is from day 133 and that latent heat transfer increases markedly from just a few days before. Is there a connection, as leafing out of all the major deciduous tree species occurs between about days 115 and 152? Increasing transpiration after leaves have expanded could lead to an increase in the water vapor concentration of the PBL, which would raise the dew point temperature and reduce the impact of frosts on frost-sensitive leaves and, in addition, would be likely to increase cloudiness, another feature that reduces the occurrence of radiation frosts.
Figure 4. Energy transfer from the Harvard Forest. Dashed line, net radiation; black, latent heat (evapotranspiration); gray, sensible heat (convection).
Figure 5. Climatic conditions over the Harvard Forest. Black, maximum temperature; gray, specific humidity; dashed line, minimum temperature.
The impact of leafing out has been assessed at Harvard Forest by determining the fluxes of energy into the PBL. These are calculated by differentiating the trends of specific humidity and mean daily temperature (figure 6) and averaging over the period from 1994 to 2000. This provides a rate of change: multiplying the rate of temperature change by the specific heat of air provides the sensible heat flux into the PBL, and multiplying the rate of change of specific humidity by the latent heat of vaporization provides the latent heat transfer. Averaging over years minimizes the impacts of weather fronts that in any one year may influence the PBL at a particular time. It is assumed that the PBL grows during the day to a maximum height of 1 km (figure 2).
Figure 6. Energy fluxes from the landscape into the planetary boundary layer. Black, latent heat; gray, sensible heat; dashed bar, tree bud burst.
The quite specific temporal patterns of sensible and latent heat transfer measured from the forested landscape (figure 4) can also be recognized in the PBL (figure 6). Sensible heat transfer is positive early in the year before leafing out, and this warms the PBL. Sensible heat transfer then declines from about day 95, whereas latent heat transfer increases up to day 155. Latent heat transfer increases during the period of leafing out, but there is also a noticeable increase before leafing out. It is most likely that this increase results from evapotranspiration from the ground flora and vernal species that are active before the forest trees.
The forested landscape does, therefore, influence the PBL by sensible and latent heat transfer, but this is achieved by only 5% to 6% of the landscape flux converging into the PBL, the remainder being dissipated, predominantly throughout the canopy layer (figure 1).
The nocturnal boundary develops after the convective PBL collapses when solar radiation declines toward zero. The NBL then develops, dependent on three major processes: turbulent mixing, losses of long-wave radiation, and energy exchange with the soil. Under clear, still nights, temperatures can drop significantly at plant level within the NBL, with the potential for frosts to occur. This has consequences in horticulture, for example, with frost-tender flowers on fruit trees. In such situations the flowers are sprayed with water, and external heaters are used quite regularly to minimize frost damage. Evapotranspiration from vegetation and soil increases the humidity of the NBL, which raises the dew point temperature and therefore reduces the risk of frost. It is notable, however, that in some desert areas such as the Southwest of the United States, where the air is dry and dew point temperatures are low, plants in the landscape are quite frost tender. Many desert plants emit a broad suite of hydrocarbons, both gaseous and particulate, some of which are detectable by human smell. Hydrocarbons generally absorb and emit long-wave radiation, so it is feasible that these hydrocarbons act to provide a minigreenhouse effect at the landscape scale, which is particularly important when the NBL develops on clear nights and where radiative cooling increases the potential for frosts (Hayden, 1998).
This potential effect has been investigated for the Sonoran Desert in Arizona. Two meteorological stations in the Arizona Meteorological Network (http://ag.arizona.edu/azmet/) have been selected, Roll and Marana. Maps of the desert (http://www.cast.uark.edu/pif/main/west/82table.html) indicate that Roll has little natural vegetation, whereas Marana is vegetated with creosote bush (Larrea tridentata) and species of sagebrush (Artemisia spp.), both of which are noticeably aromatic. The evergreen nature of many desert shrubs, including Larrea, indicates a requirement to endure frosts during the winter.
The meteorological characteristics of the NBL for Roll and Marana were determined as averages over the period 2000 to 2006, using daily maximum, minimum, and dew point temperatures and maximum and minimum observations of relative humidity. Daily observations were extracted for January and February, when there was either no or very little precipitation (this reduces the impact of high-humidity conditions and wet soils on the NBL), when there was probably little if any irrigation for any crops, and when temperatures can potentially fall into the freezing range during the night. The analysis aimed to detect whether the vegetated landscape around Marana can exert an impact on the temperatures within the NBL, particularly at plant height, and whether this impact is more significant than at Roll, with less extensive natural vegetation.
Dew point temperature and minimum temperature at Roll (figure 7) are closely correlated (regression slope is 1.02 ± 0.02, intercept–1.26 ± 0.23), with little scatter. By contrast, there is more scatter at Marana, with many occasions when the minimum temperature is higher than the dew point temperature (slope 0.75 ± 0.03, intercept 5.6 ± 0.25). The intercept for Marana indicates that, on average, the minimum temperature is 5.6°C greater than the dew point temperature, whereas at Roll, the minimum temperature is 1.3°C less than the dew point. Therefore, temperatures in the NBL at Marana appear to be elevated above the dew point temperature and more so than at Roll. The effect of this response is clearer and more ecologically and physiologically relevant when the temperatures are investigated in the chilling and freezing regions (less than 10°C) (figure 8).
Figure 7. Relationship between daily minimum and dew point temperatures in the months of January and February for Roll and Marana, Arizona.
Separating the responses to periods when the maximum relative humidity (RH) is greater than or less than 70% also differentiates between the two locations (figure 8). At Roll, the slopes of the relationships between dew point and minimum temperature are not significantly different, although the intercept occurs at a higher temperature when the RH is less than 70%. This reflects the greater opportunity for temperatures in the NBL to drop below the dew point when the air is dry.
At Marana, the two RH ranges differentiate two types of responses within the NBL. When the RH is greater than 70%, the response is similar to that in Roll, with close correlation between the minimum and dew point temperatures. This indicates for both sites that when the RH of the air is quite high, then nocturnal cooling will occur to about the dew point temperature. At that point, heat is released when water vapor condenses into the NBL, tending to stabilize the temperature. This effect also occurs when the RH is lower at Roll, but the stabilizing effect on temperature is less, as less condensation occurs in the drier air. At Marana, the relationship between minimum temperature and dew point at humidities less than 70% is quite different from that at higher humidities, and this difference becomes more marked when the dew point temperature drops below zero. The slope of the response is less steep, and there are many instances of high minimum temperatures. Although some of these observations may reflect the occurrence of warm weather fronts, the regularity of the occurrence suggests other activities in the NBL.
Figure 8. Relationship between daily minimum and dew point temperatures, for minimum temperatures less than 10°C in the months of January and February, for Roll and Marana, Arizona. Black circles, maximum relative humidity >70%; gray circles, maximum relative humidity <70%.
The low relative humidities at Marana are part of a crucial activity within the NBL and the PBL. When RH exceeds 70%, particulate hydrocarbons are enveloped with water, reducing any greenhouse-type effect of the hydrocarbons, and this appears generally to be the case at Roll but not Marana. During the day, this difference would be visible in the PBL with a blue haze at humidities lower than 70% and a white haze at more than 70%—visual verification of landscape impacts on the boundary layer. If the desert shrubs at Marana influence the NBL in this way, then there should be differences in long-wave energy exchange between the two sites. At Roll, increased long-wave energy loss from the landscape surface, at night, will cool the NBL and be little different between the two humidities if hydrocarbon particulates are not major components in the NBL. By contrast, particulate and other hydrocarbon emissions from the desert shrubs at Marana should exert significant impacts on long-wave energy emission in the NBL when the humidities are low.
The divergence of the minimum temperatures between the two sites has been investigated when the dew point temperature drops below zero and plants may be subjected to frost. At Roll, increasing long-wave energy loss from the landscape leads to cooling and to a decrease in the minimum temperature under all humidities with no significant differences in the slopes between the two humidity ranges (figure 9). At Marana, increasing long-wave emission leads to a decline in the minimum temperature when the RH is greater than 70%; however, there is no relationship when the RH is less than 70%. Long-wave emission is correlated with maximum temperature for both sites and all humidities, suggesting that day temperatures may play a key role in the long-wave emissions during low humidities at Marana.
The relationship between daytime maximum temperature and the daily temperature range (figure 10) indicates that at Roll there is a simple positive and linear relationship between the maximum temperature and the temperature range. This is also the case at Marana, but only when the maximum humidity is greater than 70%. At lower humidities, the relationship is significantly flatter, with a 0.29 ± 0.07°C increase in the temperature range for a 1°C increase in maximum temperature, compared with a 1.00 ± 0.16°C increase when humidities are over 70%. This suggests that high day temperatures are exerting an influence on temperatures in the NBL and not simply a warmer night following a warmer day, as seen at Roll. Hydrocarbon emissions from plants increase with temperature, and this appears the most likely explanation: the vegetated landscape is protecting itself from excessively low temperatures at night by producing a minigreenhouse effect with larger effects occurring following a warmer day.
Figure 9. Relationship of minimum temperature and net longwave emission for Roll and Marana. Black circles, maximum relative humidity >70%; gray circles, maximum relative humidity <70%.
The impact of the vegetated landscape on local climate is dependent on the response of the mixed PBL above the landscape. Mixing of heat and moisture from outside the area of study occurs continually, as does the inclusion of drier air from above the PBL. These continuous processes mix in the typically humid air released from the vegetated landscape. The end result of a step change in vegetation activity, such as bud burst and leafing, is an identifiable impact on the humidity and heat content of the PBL, but the landscape influence is in the order of about 5% (figure 6) for the Harvard Forest. In Nebraska, increasing the area of irrigated crops leads to a substantial 40% increase of evapotranspiration and lower temperatures (table 1). The temperature of the irrigated landscape was 1.01°C less than in 1945.
The largest impacts of the landscape appear to occur at night in the NBL. The NBL is much thinner than the PBL of the day, and with no solar radiation, long-wave radiation exchange is the key driver of the NBL climate. The impact of landscape emissions on minimum temperatures is very large, in the order of 5° to 10°C, when humidities are small and dew point temperatures are close to freezing (figures 7 to 10). Minimum temperatures at night increase with the water vapor content of the air (figure 5). As a consequence, minimum temperatures in the Sonoran desert should regularly fall below freezing, but this rarely occurs to any great extent (figure 7), which appears to be a result of significant hydrocarbon emissions from the desert plants in the landscape.
Figure 10. Relationship of maximum daily temperature range and maximum temperature for Roll and Marana. Black circles, maximum relative humidity >70%; gray circles, maximum relative humidity <70%.
The work described in this chapter has been inspired by the work of Bruce Hayden, at the University of Virginia. He is an expert in climatology and ecology, and his key work (Hayden, 1998) builds on this twin expertise to describe a wide range of landscape-scale feedbacks on climate. In addition to this description, Hayden’s work also identifies the problem in ascribing changes in the planetary layer to landscape activities as opposed to other independent regional climatic effects. This subject has been expanded here, and although it is possible to identify likely feedbacks, there is always the slightly nagging uncertainty associated with the absence of a certain cause-and-effect relationship between landscape and climate. Similar problems exist at the global scale when we consider human influences on climate. In this context, it is interesting to note that when global-scale models of vegetation are driven by fields of observed climate, this observed climate already includes the effects of climatic feedback.
Betts, A. K. 2004. Understanding hydrometeorology using global models. Bulletin of the American Meteorological Society 85: 1673–1688. General description of energy transfer into the planetary and NBLs.
Fisch, G., J. Tota, L.A.T. Machado, M.A.F. da Silva Dias, R. F. Lyra, C. A. Nobre, A. J. Dolman, and J.H.C. Gash. 2004. The convective boundary layer over pasture and forest in Amazonia. Theoretical and Applied Climatology 78: 47–59. Describes the impacts of the wet and dry seasons on the planetary boundary layer over forested and grassland landscape; a source for figure 2.
Fitzjarrald, D. R., O. C. Acevedo, and K. E. Moore. 2001. Climatic consequences of leaf presence in the eastern United States. Journal of Climate 14: 598–614. A description of an approach to quantify the impact of greening up on the planetary boundary layer; a background to the calculations for figures 4 to 6.
Geron, C., A. Guenther, J. Greenberg, T. Karl, and R. Rasmussen. 2006. Biogenic volatile organic compound emissions from desert vegetation of the southwestern US. Atmospheric Environment 40: 1645–1660. Examples of biogenic emissions from a range of plant species from the Mojave and Sonoran deserts.
Hayden, B. 1998. Ecosystem feedbacks on climate at the landscape scale. Philosophical Transactions of the Royal Society, Series B 353: 5-18. A classic exposition of an array of landscape feedbacks on climate.
Mahmood, R., S. A. Foster, T. Keeling, K. G. Hubbard, C. Carlson, and R. Leeper. 2006. Impacts of irrigation on 20th century temperature in the northern Great Plains. Global and Planetary Change 54: 1–18. Source of data for table 1, indicating how irrigation influences temperatures at the regional scale.
Webb, T. J., K. J. Gaston, L. Hannah, and F. I. Woodward. 2006. Coincident scales of forest feedback on climate and conservation in a diversity hot spot. Proceedings of the Royal Society, Series B 273: 757–765. Novel analysis using climatic and remote sensing data to determine the impacts of deforestation on climate; source of data for figure 3.