1. What do species do in ecosystems?
2. Response capacity of the biota
3. Species dominance and feedback loops
Ecosystem services are provided by biological processes and structures as well as by the geophysical environment. Biodiversity is a measure of the variation in life forms and is the result of many biological and geological processes and constraints. To understand what species do in ecosystems, particular attention must be paid to the traits of these species, such as their optimal temperature for growth, their ability to avoid predators, or their nutrient uptake capacity. It is the distribution of traits relevant for particular ecosystem services, so-called trait spectra, that determines the performance of the biological community as a whole. The variation in particular traits within the community, such as the range of temperature tolerances, is a measure of the response capacity, i.e., the overall ability of the community, species, and individuals to respond to changes in environmental factors, such as temperature changes. Greater response capacity reduces variability in ecosystem services under environmental variability. This is particularly important for systems that are susceptible to critical transitions. Landscape patterns and processes provide a regional source of trait variability for local communities and thus maintain response capacity.
abiotic environment. The chemical, geological, and physical part of the ecosystem.
critical transitions. A change of the dominating feedback processes in an ecosystem, with implications for ecosystem structure and functioning. Systems undergoing a critical transition may be profoundly different before and after the transition.
response capacity. The ability of a local community to respond to changes in environmental drivers.
trait spectra. The abundance-weighted distribution of particular traits in the community.
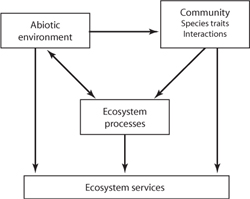
Ecosystem services are sustained by an interaction between abiotic and biological processes. Thus, biological processes such as primary production account for only a part of ecosystem services. Water provisioning depends on biological processes as well as physical ones, such as those that drive the climate system. Similarly, it may be the presence of a particular striking feature, such as a blue whale or the Grand Canyon, that delivers a service, largely independent of any current biological processes (although historic biological processes were required to produce the blue whale and the Grand Canyon to begin with). The abiotic environment, such as the geomorphology of the landscape or the climate and the hydrosphere, largely sets the constraints within which biological communities develop. The abiotic environment is, however, by no means unaffected by the biological system. Water flow, for example, is determined by the climate and hydrology, but trees may play a large role in channeling water back to the atmosphere, thereby potentially affecting groundwater levels and even large-scale climate patterns (figure 1).
The role of particular species in the ecosystem processes is determined by functional and morphological characteristics, i.e., traits. Functionally similar species may be involved in sustaining particular ecosystem processes either directly (e.g., provisioning services) or indirectly (e.g., support or regulating services). The attributes (traits) of particular species and the effects of environmental variables on the species as well as on physical processes fundamentally determine which ecosystem services are provided and at what level. To understand the role of species traits and the role of environmental variables in ecosystem services, we first need to elaborate on what species do in ecosystems.
Figure 1. Ecosystem services are provided by the abiotic environment, the biological community, and the ecosystem processes. Although the abiotic environment largely constrains the biological communities, the ecological processes can also affect the abiotic environment through such processes as withering and water vapor flow
The year 2007 marked the 300th birthday of Carl Linnaeus, the father of taxonomy. His Systema Naturae was used to give name, and thus identity, to most species known to humans. Thus, one might argue that he also was the father of the concept of species richness because, in order to name species, one needs to be able to identify them. However, a name does not reveal much about the role of a species in ecological processes. In practice, species classifications have also been based on morphology or, more recently, on their genetic code rather than on their function or what they do. Biodiversity should be a full measure of variation in life forms, and thus, when it comes to understanding how communities function, one should add some measures of traits to species’ identities. In most studies, abundance has been used as a proxy for ecosystem function because it is an easy variable to measure, and one can argue that abundance is often at least proportional to some process sustained by particular organisms. However, abundance is an imperfect measure at best because (1) the ability of a set of organisms to sustain a particular function may “saturate” and thus be unrelated to abundance; and (2) the species most abundant in the system may be functionally similar or dissimilar, with real consequences for the range of services that can thus be supported. Traits represented in the community can be related to functioning and structure in a much more direct way.
Lawton (1994) posed the question “What do species do in ecosystems?” which helped inspire a research field devoted to understanding the relationship between biodiversity and ecosystem functioning. Several reviews that summarize this research field (e.g., Hooper et al., 2005) conclude that some measure of biodiversity (usually focused on species rather than, say, variation in communities or landscapes) provides a statistical estimate of the traits of the species in a community that in turn may tell us something about the functioning of a community. The more species, the more traits, for instance. Why then does one not focus more directly on trait distributions in communities in addition to species’ lists and relative abundances? Relevant traits are not always easy to measure. Morphological traits are generally easier to measure and are sometimes called soft traits, whereas more process-relevant traits such as nutrient uptake rate, temperature response, or predator-avoidance traits are harder to quantify. Research to elucidate how to link hard and soft traits is a growing field that holds promise in helping us understand the role of the interaction among species and the environment for ecosystem services.
Species richness is a measure of the number of tax-onomic units, generally species, within a biological community or ecosystem. Sometimes measures of species diversity include aspects of relative abundances of species, such as the Shannon index—communities with a more even distribution of species abundances are considered more diverse than those with a similar number of species but dominated by one or two species. In research focused on the role of species diversity for ecosystem functioning (often implying ecosystem services, i.e., processes or structures that are of human value), the contrasting hypothesis has often been species identity; i.e., does species diversity per se or species identity control ecosystem functioning on average? The notion “on average” is meant to imply that one tries to compare over many different sets of species in order to find effects that are statistically either determined by the number of species or by the particular set of species. If the analysis favors number of species over particular species, it does not imply that particular species do not have great importance for the performance and maintenance of ecosystem processes but, rather, that there is some even more important impact of the “average effect” of biodiversity. But this effect can be very hard to parse. For instance, scientists may manipulate conditions in order to create communities with different species numbers, but in manipulating conditions, they may also be influencing species identity. If a scientist uses existing patches to examine the relationship between diversity and functioning, then abiotic differences in the patches may be altering species identity as well as number. In other words, changes in species identity and species number may be correlated. When we disentangle this issue with further analysis, we will find that the crucial aspects really are to understand processes that either sustain or decrease biodiversity rather than biodiversity per se.
In a changing world, a fundamental support service is the ability of biological communities to respond to change without major loss in structures or processes that provide important ecosystem services. This response capacity is a multilevel phenomenon occurring from the level of the organisms to biomes as well as from daily to decadal time scales. I illustrate this with the response of biological communities to temperature changes, although many other changes and adaptations are important. At the most fundamental level, any protein has a temperature dependence that determines the rate at which it catalyzes chemical reactions. Proteins are also sensitive to high temperatures, which disrupt the delicate folding structures that give them their catalytic properties. In the balance between these rate-enhancing and rate-disrupting temperature dependencies, there is an optimum temperature that gives the highest rate of functioning for a particular protein. But there is also a quite broad range of temperature at which a particular protein will work well, albeit not at its optimal rate. This is the physiological response capacity. Many species have several varieties of each protein type encoded, which have slightly different temperature dependencies. With temperature-dependent cues, these proteins can be turned on or off in the genome and thus provide a phenotypic plasticity that broadens the temperature tolerance of an organism. Within a population, there are many individuals, and because of genetic recombination and mutations during reproduction, each individual may have different combinations of these proteins. Thus, within a population, the response capacity may be even broader because of additional variation in the traits attributed to protein temperature sensitivity. On evolutionary time scales, the within-species variation of these traits is also directly proportional to the rate at which natural selection can change the mean trait in the population in response to selective forces, e.g., an increase in temperature.
In regard to ecosystem services, there are additional layers of response capacity. Different species can contribute to the same process or structure that sustains an ecosystem service. This means that variation in temperature dependence between these functionally similar species also increases response capacity of the whole community, as species with better-adapted responses can replace less well-adapted species by means of competition and community succession. Again, we find that the capacity of a group of functionally similar species to change the mean trait of the group in response to changing environmental factors, e.g., temperature, is directly proportional to the variation among species and individuals within this group. In contrast to the evolutionary process, this occurs in ecological time, which often is a faster response than evolutionary responses.
Other sources of trait variation in a community are dispersal and migration processes. When the variation in traits increases with the investigated area, and dispersal/migration processes are present, the area can act as a source pool for local trait variation and thus enhance local response capacity (see section 4). Thus, distribution of traits, or so-called trait spectra, in a community is more informative than species richness per se in understanding how a community will function. Trait spectra are measured as relative abundances over a particular trait, such as the rate of nitrogen fixing (0 for nonfixers) or temperature optima for temperature response–related traits. The trait spectra for a trait such as the optimal temperature for growth are likely to be very narrow in areas with little variation in temperature such as the tropics and much wider in temperate areas that can sustain cold- and warm-adapted species that grow during different seasons. Thus, even though the tropical areas will have many more species, the variation in a particular trait, here temperature optima for growth, is likely to show much more variation in the relatively species-poor temperate areas. This calls into question the notion that species richness may be a good surrogate for functional trait diversity; different processes may be driving species richness and trait variation within communities. Whether or not species richness correlates to trait variation depends on the trait in question and on the historical forces shaping the community.
It should be noted that maintenance of response diversity can also carry a cost. Maintenance of high variability under constant conditions leads to lower functioning than if all individuals had an optimum at or near the mean condition. It is only when environmental factors change that wider trait spectra, and thus response capacities, increase overall productivity in the long term. The effect of trait variation on the capacity of a system to respond must be measured with respect to expected variation in the environment. That is, one cannot say that one trait variation is better than another, as it depends on how much environmental variation each trait variation has to buffer in the given location. “Too much” trait variation, while still maintaining response capacity, may reduce the overall functioning of the system with respect to expected conditions. There is a trade-off between optimal functioning and responsive capacity.
Trait variation, whether it is between or within species, reduces the variation in the performance of the whole community and hence reduces the variability in the ecosystem services they sustain. This can be particularly important if there are thresholds that may be crossed when particular processes fall below a certain level, leading to large changes in the ecosystem. A consequence of this is that there may be a tradeoff between the provisioning of an ecosystem service at some “optimal” level and the resilience of the system.
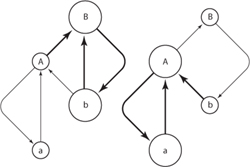
Ecosystems are made up of complex interaction networks. Food webs, for example, consist of many weak interactions and a few strong ones that have a disproportionate effect on the whole structure of the food web. In addition, some interactions can cause feedback loops that either dampen or self-reinforce different configurations of the system. An example of such a self-reinforcing interaction loop is shown in figure 2. Two species, each with two size classes, for example fish, interact. Species A is generally smaller than species B. Adult individuals of species B can prey on adults of species A. However, species A’s adults are larger than the juvenile class of species B, on which they can prey. This leads to two different feedback loops in this very simple food web. First, if there is a high abundance of species A and low abundance of species B (right-hand side of figure 2), the adults of species A can prevent population growth of B by preying on their young. A second scenario is when adults of species A are in low abundance, and species B is abundant. Then species B can control the population size of species A by preying on their adults.
These feedbacks are very common in ecological systems, and the example above may be illustrated with the Baltic food web, where this mechanism is present in the interaction between cod and sprat. Ecosystem services can be strongly dependent on species that are part of such an interaction feedback loop, the most well-known examples being kelp forests and coral reef systems. Furthermore, abiotic components of ecosystems, such as nutrient release from sediments, can also contribute to creating multiple self-reinforcing states through their nonlinear response to environmental variables. For instance, release of nutrients from sediments can, through a series of interactions, cause oxygen depletion in lakes, which can lead to further release of nutrients from sediments, creating a strong positive feedback loop. The sensitivity of particular ecosystem services to these feedback interactions depends on the relative strength of the interaction and its position in the whole network. The relative effect of a sudden shift from one feedback loop to another thus depends on the diversity of the system components and the relative distribution of interactions. Systems that have fewer interactions are more likely to have a few strongly dominating processes or feedbacks, whereas systems with many interactions may have more and relatively weaker interactions. This may suggest that systems with fewer interactions experience fewer but more dramatic shifts in feedback processes, whereas systems with many interactions may experience more frequent but less severe ones. However, what actually happens in real ecosystems depends very much on the particular species interactions, and little can be generalized from species richness alone. Thus, one could say that sensitivity to critical transition phenomena in ecosystems depends, first, on the traits of the individual interactions and, second, on average link density and number of nodes, which may, at least on average, be reflected in the aggregate measure of species richness.
Figure 2. An example of critical transition in dominant feedback processes in food webs. Species B is slightly larger than species A and can eat the adults of A (capital letter). However, adult individuals of species A can predate on juvenile individuals of species B (lower case). The system can tend to be in either of these states because the feedback loop reinforces itself. For example, it is possible, e.g., by overfishing, to reduce adults of species B until the system flips into a state dominated by species A predation on juveniles of species B.
Communities are dynamic, and thus, so are trait spectra and species richness. Even if species richness remains constant, there can be a large turnover of species because of species responses to environmental drivers and of dispersal or movement of species. This can result in rapid changes in community characteristics such as the trait spectra or interaction networks and thus also affect major ecosystem processes. The influx of new species into the species pool is largely determined by migration and dispersal (assuming that the arriving species can thrive in the location) and, on longer time scales, by evolutionary processes.
Dispersal and migration processes are essential support services that sustain a biological community’s ability to change in response to a change in environmental drivers, such as current global climate changes. For spatial variation in biodiversity to have a positive impact on communities’ response capacity, there need to be (1) a positive relation between area and species richness (or rather trait variation) such that immigrating species (or individuals) are different from local species (or individuals) and can contribute to better performance, resource use, or structure that may support ecosystem services, and (2) dispersal/migration corridors or vectors that allow movement of individuals and dispersal vectors. Dispersal is the spatial process of redistribution of organisms or abiotic entities. Migration means an active movement by organisms. Dispersal vectors can be organisms such as pollinators or seed carriers, e.g., flying foxes. Sometimes dispersal vectors are important for transportation of abiotic components such as nutrients, e.g., herds of ungulates that fertilize areas in their migration track.
Figure 3. The relation between landscape processes and ecosystem services. Patches of habitable land, here exemplified by small areas of forests in Madagascar protected by religious taboos, are connected by the distance that species providing certain ecosystem services can travel: (top) 500 m, (middle) 1000 m, and (bottom) 1500 m. This creates different-sized components in the landscape that can often hold higher biodiversity. (Adapted from Bodin, Ö., and J. Norberg. 2007. A network approach for analyzing spatially structured populations in fragmented landscape. Landscape Ecology 22: 31–44)
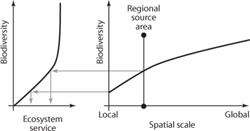
The effective area that is available as a regional pool of species for a local site depends on how this site is connected to other sites and the dispersal ability of the organisms involved in a particular ecosystem process. figure 3 shows an example from Madagascar where small patches of forest, mainly protected by religious taboos, host species that act as seed disperser (lemurs) and pollinators (bees). The effective area for the potential species pool varies greatly depending on the mean travel distance of organisms (shown in the different panels of figure 3), which determines how many local sites join a cluster of connected sites. The resulting effective regional species pool and the variability in traits held by species in this pool may have a positive effect on ecosystem processes and, if these are linked to a particular ecosystem service, have a positive effect on ecosystem service provisioning as is illustrated in figure 4.
Figure 4. Depending on the average range of movement of species, the landscape is more or less connected, which determines the area that can act as an effective regional source of species for local habitats. If species biodiversity is positively related to area, then a larger regional pool may be able to sustain higher local biodiversity. When this spatial phenomenon is combined with the general positive effect of biodiversity for ecosystem processes, we can see how landscape patterns and dispersal processes could potentially increase biodiversity and thus community response capacity, leading to less variability and better provisioning of ecosystem services.
Ecologists have spent the last decade demonstrating the connection between various measures of species identity and diversity and the functioning of ecological systems. As suggested here, and by many other scientists, it may be the suite of traits present in a system, rather than species diversity itself, that is the most important determinant of ecosystem functioning and hence the provisioning of ecosystem services. Maintaining or enhancing trait diversity in natural or managed ecosystems can buffer those systems against change—enhancing their resilience—although this can also come at a cost to optimal functioning or provisioning of services for a particular set of conditions. The emerging scientific knowledge about trait spectra and how they correlate to other aspects of the ecosystem should aid in effectively balancing efficiency and resilience as the world increasingly impacts and manages the landscapes that deliver the suite of ecosystem services upon which humanity depends.
Elmqvist, T., C. Folke, M. Nystrom, G. Peterson, J. Bengtsson, B. Walker, and J. Norberg. 2003. Response diversity, ecosystem change, and resilience. Frontiers in Ecology and the Environment 1: 488–494.
Hooper, D. U., F. S. Chapin III, J. J. Ewel, A. Hector, P. Inchausti, S. Lavorel, J. H. Lawton, D. M. Lodge, M. Loreau, S. Naeem, B. Schmid, H. Setata, A. J. Symstad, J. Vandermeer, and D. A. Wardle. 2005. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecological Monographs 75: 3–35.
Scheffer, M. Critical Transitions in Nature and Society. Princeton, NJ: Princeton University Press.
Tilman, D. 2001. An evolutionary approach to ecosystem functioning. Proceedings of the National Academy of Sciences U.S.A. 98: 10979–10980.