FIVE
The Formation and Maintenance of Populations and Assemblages
CONTENTS
Application to Fish Assemblages
A Return to Historical Effects
Species Characteristics and Assemblage Formation
Colonization in Some Systems but Not Others
Colonization by Some Taxa and Not Others: The Role of Trophic Position
Movement and Assemblage Formation
Movement Inferred from Colonization Studies
Movement Inferred from Tagging Studies
ECOLOGISTS STUDYING FISH populations and assemblages most often are dealing with their subjects already formed, at least in the sense of ecological time. In Part 1, the examples of the colonization of new habitats that were opened up following glacial retreat showed how species might be added to such systems. Obviously, opportunities to study natural assemblages in the early stages of their formation are rare. However, opportunities do occur when assemblages repopulate after being eliminated due to loss of habitat during droughts or because of intense flooding that can cause local or regional extirpation of fishes.
This chapter deals with issues of how species are added to assemblages, both from the standpoint of how preexisting species might alter the outcome and from the standpoint of how adept fishes are in moving about their environment (see figure in Part 2). In thinking about how fish assemblages might be formed or reconstituted, a key question is how species are added to an assemblage, and whether there are “rules,” other than having a suitable habitat, that govern the order and kind of species additions. Are fish assemblages built solely on the basis of random colonization opportunities by the component species or does the presence of a certain species, or groups of species, change the probability of successful establishment by yet other species? The resolution of the question could be approached at a larger scale to include both historical (i.e., lineage) and dispersal components, although much of the ecological literature has focused on a finer scale that emphasizes dispersal (but see Gorman 1992; Winston 1995). As natural communities are increasingly besieged by nonnative taxa, the answer to the questions of how species are added to assemblages has also taken on new importance in the efforts to develop models predicting the vulnerability of native communities to invasion by nonnative taxa (see also Chapter 15).
Understanding the formation and maintenance of assemblages also requires an understanding of the vagility of fishes. Without some degree of movement, fishes would be unable to colonize new habitats or to recolonize habitats from which they have been eliminated because of biotic or physical impacts. Movement ability varies widely among fish taxa and within a given taxon by life-history stage, physiological state, and simply by individual differences.
COLONIZATION OF ASSEMBLAGES
The Search for Assembly Rules
The question of whether or not there are ecological “rules” governing how species are added to communities has intrigued ecologists for decades. Indeed, long before that, Plato provided the first attempt in the fourth century BC when he considered how human communities were assembled (Belyea and Lancaster 1999). A seminal paper by Diamond (1975) on the addition of bird species to islands initiated a debate that continues in some measure to the present. Diamond hypothesized that species composing a community are “selected and coadjusted in their niches and abundances, so as to fit with each other and to resist invaders.” A more formal definition of assembly rules was proposed by Belyea and Lancaster (1999) in an effort to make the distinction between underlying mechanisms, rules related to the mechanisms, and the resultant patterns resulting from the rules. In their definition, assembly rules are “general and mechanistic, and operate within the case-specific constraints imposed by colonization sequence and environment.”
For several decades now since Diamond’s (1975) publication, debate has centered on issues of whether communities are structured as opposed to random. Such debate is, in part, because Diamond did not first clearly demonstrate that the bird communities he studied were actually structured rather than random collections of species before he proposed specific rules for assembly (Weiher and Keddy 1995). Later studies focused on distinguishing between structured versus random assemblages by testing observed patterns of species occurrences against null models of species assembly, resulting in the demonstration that at least some of the patterns that ecologists used to invoke assembly rules could be attributed to chance (Connor and Simberloff 1979). The use of null models increased the rigor of studies on community structure and assembly rules, although philosophical differences in constructing null models, such as the appropriate species pool for building the model, have continued to foster debate (see Gotelli and Graves 1996; Box 5.1).
BOX 5.1 • Null Models in Community Ecology
The experimental tools that are conventionally available in community ecology are field, laboratory, and so-called natural experiments (Gotelli and Graves 1996). Well-designed experiments require both tests and controls so that the effect of a potential factor can be evaluated. However, ecological communities, or even a subset such as fish assemblages, are notoriously complex, making large-scale manipulations or laboratory experiments impractical from the standpoint of logistics, time, or money. For instance, assessing the impact of Species X on the colonization probability of Species Y could require eliminating Species X from randomly chosen sites, introducing Species Y, and then comparing the colonization success of Species Y between sites with and without Species X.
Coupled with the ethical or legal issues of manipulating systems that might already be at risk, the potential of being able to “manipulate” communities statistically is certainly an attractive alternative (Connor and Simberloff 1986). In the case of Species X and Y, a null model might be constructed such that the probability of the two species co-occurring in the absence of any interaction could be calculated and then compared with the actual co-occurrence. Simply put, a null model “is an attempt to generate the distribution of values for the variable of interest in the absence of a putative causal process” (Connor and Simberloff 1986). However, the challenge is to design an appropriate null model for the situation being tested! The literature on null models, relative to testing forces involved in shaping communities, is a litany of proposed methods followed by criticisms thereof. (For a few examples see Connor and Simberloff 1986; Brown et al. 2000, 2002; Stone et al. 2000; and Fox 2001.)
The use of null models in addressing the question of community assembly has focused on the interactions of species with potential sites and the interactions among species (Gotelli and Graves 1996). As outlined by Gotelli and Graves (1996), all models that have been used to test the hypotheses generated by assembly rules have identifiable shortcomings. There are key questions to consider: (1) what species to include in the species pool from which potential assemblages are randomly, or otherwise, extracted; (2) rather than species, should some other category, such as functional groups (i.e., a collection of species all of which have a similar ecological function) be used; (3) what metric is most appropriate to determine whether or not assemblage organization follows a random pattern; and (4) what is the most appropriate way to design a null model (being that the possibilities are large)? Obviously, null models, although potentially very useful, are not without their own set of issues. The use of null models in ecological applications is treated in detail by Gotelli and Graves (1996).
A challenging issue is one of choosing a preinteraction species pool—one that has not already been subjected to long periods of interaction (such as competition) that would have caused the removal or stifling of speciation of certain species or lineages (Colwell and Winkler 1984). As a consequence of sampling a postinteraction species pool, the strength of the effect being measured is consistently underestimated. The issue of sampling a postinteraction species pool is termed the “Narcissus Effect” (Colwell and Winkler 1984), named after the Greek god Narcissus who, according to one version of the story, when gazing into a pool saw only his own image and could not discern the depth of the pool.
Clearly there are challenges to providing rigorous tests of community assembly rules. One approach that has been taken is to consider predictions that would be met by assemblages if, indeed, assembly rules were important. These have primarily focused on the random versus structured dichotomy. This approach was taken by Gotelli and McCabe (2002), who conducted a meta-analysis of 96 published studies of species presence-absence matrices, comprising a wide range of taxa and geographic locations, to determine the generality of Diamond’s (1975) model of assembly rules. Their results showed that most plant and animal assemblages do follow predictions of Diamond’s assembly rules in that there are fewer species combinations than expected by chance (i.e., communities are nonrandom), a result also shown by community assembly studies using laboratory microecosystems (Drake et al. 1993). In an effort to move studies of community assembly forward, Weiher and Keddy (1995) argued that the issue of whether there are nonrandom communities has long been resolved in the affirmative and that it is time to move on to newer questions. Although fish assemblages treated by Gotelli and McCabe (2002) showed less evidence for structuring by competition than did other groups of organisms, the low sample size (n=3) precluded any meaningful generalities.
Application to Fish Assemblages
There have been relatively few studies that specifically addressed the reality of assembly rules governing fish assemblages or included fish assemblages in more general analyses. The studies of fish assemblages that have evaluated the predictions derived from community assembly rules have had mixed results in terms of finding support.
Matthews (1982) provided an early attempt to determine if assembly rules (sensu, Diamond 1975) could be applied to the speciesrich Ozark stream fish assemblages of eastern North America. The study focused on six small streams occupied by a suite of 13 largely insectivorous species in the minnow, silverside, and topminnow families. All of the species had overlapping ranges even though each species did not occur in all of the streams. One of the hypotheses tested was that pairs of the common/abundant species were associated randomly among the six small streams. The alternative hypothesis, that species occurred only in certain combinations, would support the idea of assembly rules. The outcome of this statistical experiment showed that the occurrence of species pairs based on the random assortment of species into watersheds (keeping both the number of watersheds in which a species was common/abundant and the number of species per watershed equal to observed values) was not different from the observed values. From this, Matthews (1982) concluded that no other process, other than random assortment, was required to explain the pattern of mutual abundance of the small insectivorous fishes in watersheds.
Although this simple statistical experiment is appealing, a follow-up paper (Biehl and Matthews 1984) used more sophisticated statistical approaches to reexamine the results of Matthews (1982). Relative to the previous conclusion, they found that both the way the simulations were computed as well as the way the observed and expected values were compared (chi-square test) may have affected the outcome of the study (i.e., reducing the confidence in the conclusions). Subsequent studies have built on Matthews’s work; not surprisingly, the following two examples are from biologists who did their doctoral studies under the direction of W. J. Matthews.
Winston (1995) tested predictions coming from Diamond’s assembly rules with assemblages of phoxinin cyprinids (i.e., all minnow genera present in the study area except the leuciscin, Notemigonus) inhabiting streams of the Red River drainage in Texas, Oklahoma, and Arkansas. He found that species pairs that were most similar morphologically, and thus most likely to experience greater competition, co-occurred significantly less often than expected by chance. A potential problem with this outcome is that the absence of species’ co-occurrences could be due to historical as well as ecological reasons. For instance, closely related species could have arisen through allopatric speciation and thus have different spatial distributions by virtue of being adapted to slightly different environments (Winston 1995).
In a second test, Winston (1995) was able to rule out the historical explanation by showing that species pairs that were evolutionarily most closely related (based on a cladistic analysis) did not show co-occurrence patterns that differed from random patterns. Consequently, he argued that interspecific competition was the “most likely mechanism causing the pattern.” These results support a prediction of Diamond’s assembly rules—namely that certain species combinations do not occur in nature. However, the actual mechanism(s) responsible for the nonrandom pattern of species occurrence in this study are unknown.
In a study of benthic fishes living in small headwater streams of northeastern Oklahoma, Taylor (1996) also showed that species occurrence patterns can be affected by other resident species. Taylor compared the actual species area relationship with one predicted by a null model and showed that actual species richness relative to the area of habitat declined much more steeply than expected for smaller habitats. This suggested that some factor in addition to area was influencing the number of species. By following this regional analysis with manipulative field experiments, Taylor (1996) was able to show that asymmetrical interference competition between Banded Sculpin (Cottus carolinae) and Orangethroat Darter (Etheostoma spectabile) was responsible in part for the difference between observed and expected species densities (see also Chapter 11).
Thus there is evidence that freshwater fish assemblages can be structured, as well as evidence that competition between species pairs is one of the mechanisms causing such structuring (see also Chapter 11). However, in general, studies addressing factors governing the formation of fish assemblages have been surprisingly rare and have been restricted to only one general region, the Great Plains of North America.
A Return to Historical Effects
The concept of community assembly rules, as it has developed, focuses primarily on current ecological interactions. As I have already emphasized in earlier chapters (see Chapter 2; Figure 2.1), contemporary species interactions must be viewed in the context of history. For instance, Gorman (1992) analyzed assemblage formation in several Ozark upland streams and argued that general patterns of habitat use and ecological interactions were “ancient and not the result of an ongoing process.” Consequently, the occurrence of fish species on a macrogeographic level (i.e., drainage) was best predicted by historical, biogeographic factors. In contrast, ecological processes best predicted abundance patterns in local assemblages. Thus community assembly was largely determined by historical factors, whereas contemporary ecological processes affected patterns of species’ abundances but not occurrences. The contrast with the studies by Winston (1995) and Taylor (1996) is perhaps one of scale—at what spatial level do contemporary interactions control not only abundances but also presence or absence of species?
Species Characteristics and Assemblage Formation
Are species characteristics such as body size, geographic range, trophic position, and population size related to their ability to colonize habitats? In a long-term study of fishes in the Cimarron River in Oklahoma, Gotelli and Taylor (1999b) found that colonization potential of native fish species at 10 study sites was, not surprisingly, correlated with species’ abundance, but that geographic range size and body size were not. The species pool considered by Gotelli and Taylor (1999b) was based on the known native fish fauna of the Cimarron River from the Oklahoma study sites, so the concern of adaptation to the regional environment was not an issue.
Other studies have examined the colonization potential of species that are not native to the general region of the study systems. Information on these situations can provide general insight into factors important in the addition of species to assemblages (see also Chapter 15).
THE MATCH BETWEEN SPECIES TOLERANCES AND THE PHYSICAL HABITAT Moyle and Light (1996a, b) developed a set of empirically based predictions regarding colonization success in fishes. Although the predictions were largely developed to understand invasions of nonnative species, most are also applicable to colonization potential of native species with the caveat that they were developed for western (California) fish assemblages. A principal conclusion of Moyle and Light’s work in California streams and estuaries was that “the most important factor determining the success of an invading fish species is the match between the invader and the hydrologic regime.”
Colonization in Some Systems but Not Others
In general, most studies support the view that all aquatic systems are potentially invasible, although systems that have been highly altered tend to be more prone to invasions (Moyle and Light 1996a). Numerous authors have suggested that species-rich assemblages, such as those found in the southeastern United States, are more resistant to invasion by nonnative taxa when compared with species-poor systems, although there are exceptions to this (e.g., Ross 1991; Lodge 1993). However, given the condition that where there are species-rich systems, and that these systems have gained species richness both by in situ speciation and colonization, it stands to reason that species with appropriate physiological tolerances and ecological characteristics can enter and become established.
Colonization by Some Taxa and Not Others: The Role of Trophic Position
Resource use, and especially trophic position, are thought to be important in predicting the invasion success of nonnative fishes. In California streams, successful invaders of generally unmodified aquatic systems tend to be at one extreme or the other of the trophic pyramid—either top predators or detritivores/omnivores. In both cases, these are trophic levels that tend not to be food limited (Moyle and Light 1996b). A more recent study incorporating stable isotope analysis (see Chapter 11 for details on stable isotope analysis) tested the Moyle and Light hypothesis, focusing on the base of the food web (Gido and Franssen 2007). In fact, the trophic position of nonnative fishes tended to be lower than native fishes in four of the five study rivers, although the differences were small and not significant at the 0.05 level. Because their study included five streams located from New Mexico to Kansas, they were also able to compare the trophic position of four species in streams where they were native and in those where they were nonnative. In this case, three of the four species showed significantly lower trophic positions in assemblages where they were nonnative relative to where they were native. Thus this study is at least consistent with the hypothesis that low trophic level facilitates colonization.
The San Juan River, a major tributary of the lower Colorado River, has been changed by impoundments and flow modifications and has a fish fauna that includes both native and nonnative species (Gido and Propst 1999). The food web of small-bodied fishes from secondary channels and from the margin of the main channel includes three trophic levels, as determined from stable isotope analysis (δ15N ratios): a primarily detrital base, a primary consumer level dominated by midges (Chironomidae), and a secondary consumer level that includes all fishes (Gido et al. 2006). Fishes from the secondary channels showed higher food overlap than those in the main channel, indicating convergence on the same food source. Also, fishes found in swifter water in the main channel, principally Speckled Dace (Rhinichthys osculus) and Channel Catfish (Ictalurus punctatus), had slightly elevated trophic levels. Although all fishes were grouped as secondary consumers, the more common native fishes tended to feed at a higher trophic level (mean native = 3.18) than most of the nonnative fishes (mean nonnative = 3.03), a finding at least consistent with part of the Moyle and Light hypothesis (Figure 5.1).
The Moyle and Light hypothesis, predicting the invasion of nonnative fishes at the highest or lowest trophic levels, has also been tested for fishes in the Gila River system in southwest New Mexico (Pilger et al. 2010). This portion of the Gila River has received relatively low human impacts but has been subject to widespread introduction of nonnative fishes, which potentially put continued persistence of native fishes at risk. The study was based on both stable isotope ratios and gut analyses and showed that native and nonnative fishes fed at multiple trophic levels. Large-bodied native fishes, Adult Sonora Sucker (Catostomus insignis) and Desert Sucker (Catostomus clarkii), fed at low trophic levels as adults, but at a higher trophic level as juveniles. In contrast, large-bodied nonnative fishes, such as Rainbow (Oncorhynchus mykiss) and Brown (Salmo trutta) trout, Smallmouth Bass (Micropterus dolomieu), and Yellow Bullhead (Ameiurus natalis) fed at lower levels as juveniles and then at higher trophic levels as adults. Native small-bodied fishes such as Longfin Dace (Agosia chrysogaster), Spikedace (Meda fulgida), Speckled Dace, and Loach Minnow (Tiaroga cobitis), were primarily insectivorous; the Headwater Chub (Gila nigra) was the only native piscivorous fish but also consumed large amounts of algae. Overall, nonnative fishes foraged at higher trophic levels on fishes and large, predaceous aquatic invertebrates, compared to native fishes. The introduction of nonnative fishes has extended the maximum food chain length in the upper Gila River fish assemblages to higher trophic levels, although long-term survival of at least some of the native fishes is in question as a consequence (Pilger et al. 2010).
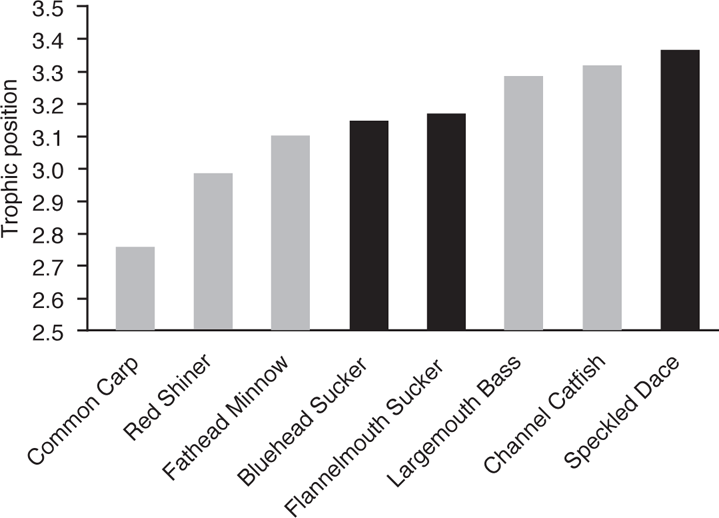
FIGURE 5.1. Trophic position, as determined from stable isotope analysis (δ15N ratios), for native (black bars) and nonnative (gray bars) fishes in the San Juan River. Based on Gido et al. (2006).
Colonization Models
Strange and Foin (2001) developed a model to predict the invasion success of six fish species (including native and nonindigenous) to cool-water (trout zone) streams, the Sagehen and Martis creeks, located on the eastern slope of the Sierra Nevada mountains near Lake Tahoe. The model incorporated life-history responses to biotic factors of competition and predation and the physical factor of stream discharge. Predictions of the model were tested against a long-term data set of relative abundance of fishes for Martis Creek from 1979 to 1994 and Sagehen Creek from 1952 to 1961. Model predictions generally captured major trends in species’ relative abundances for Martis Creek and showed excellent fit overall to data for Sagehen Creek, although some species showed better fit than others (Figure 5.2). Key points from this study of a six-species system are that invasion sequence and biotic interactions (competition and predation) were all important, but that the relative importance of these factors varied based on physical conditions. In particular, the sequence of floods and droughts had a major impact on the relative abundance and potential for invasion of each species. One of the lessons from studies such as this, which incorporate the impact of physical habitat variation, is that the formation of fish assemblages is largely nondeterministic—that is, the final assemblage cannot be totally predicted by knowing the species pool available for colonization. As stated by Strange and Foin (2001), “assembly from the same species pool is likely to result in multiple or alternate states as the physical regime alters rates of biotic processes.”
Another set of studies used the extensive database on successful and unsuccessful introductions of fishes in California to develop models predicting characteristics of successful colonizers (Marchetti et al. 2004a, b). In terms of becoming established, the best predictive model included physical characteristics of the invading species (body size and physiological tolerance), demographic characteristics (area of population), niche characteristics (trophic position), and behavioral characteristics (parental care of young). Of these, greater physiological tolerance and the degree of parental care seemed particularly important in aiding establishment.
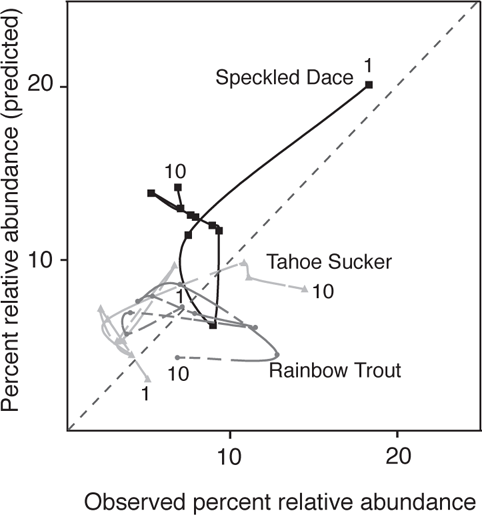
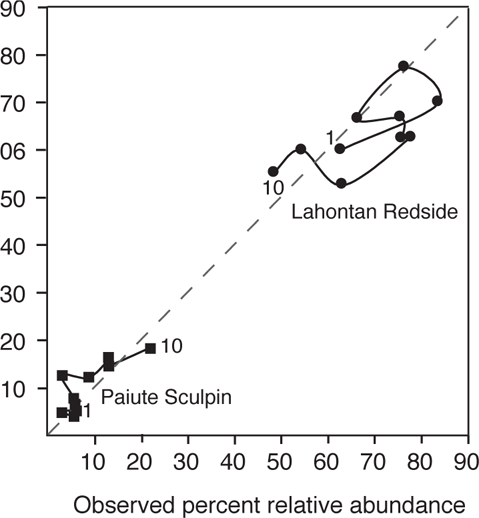
FIGURE 5.2. Relative abundances of five of the six species from Sagehen Creek, California, based on model predictions and observed values over a 10-year period. Numerals indicate starting and ending years; the dashed line shows a 1:1 relationship. Based on data from Strange and Foin (2001).
MOVEMENT AND ASSEMBLAGE FORMATION
Freshwater fishes, with few exceptions, are naturally limited in their access to new habitats by suitable aquatic connectivity. Thus understanding how fish assemblages are formed also requires understanding the movement capability of potential colonizers found in regional species pools. To a certain extent, this tempers what one can learn about community assembly by examining nonnative taxa, given that such taxa often move more through human intervention than by natural processes (see also Chapter 15).
The terms “dispersal” and “migration” are often used to describe the movement of organisms outside of their home range. Briefly, dispersal has most often been used to refer to a general outward spreading of individuals away from each other, such as from a starting group. Consequently, it is a group definition because it is based on the movement of one individual relative to the movement of one or more other individuals (Begon et al. 1996). Dingle (1996) recommended the term “ranging” in place of dispersal. Ranging refers to individual behavior and is “the departure from the current habitat patch (emigration), the seeking of a new patch, and the occupation of the first available and suitable habitat patch discovered (immigration).” In older works, the definition of migration was constrained to include movement to and from an area or areas. For instance, Harden Jones (1968) defined migration as “a class of movement which impels migrants to return to the region from which they have migrated.” If an organism did not return, however far or periodic its travels, then the term migration was deemed inappropriate. A recent trend is to recognize migration as a persistent movement of individuals of a species from one habitat to another so that the location of each life-history stage is optimized in terms of needed resources (Begon et al. 1996; Dingle 1996). Migration usually involves distinct pre- and postmigratory behaviors and includes physiological changes, such as the reallocation of energy to support long-distance movement (Dingle 1996).
In the previous section, the match between a habitat and a potential colonizer was an important factor in understanding the establishment of nonindigenous species. However, the match between an organism and a potential habitat means nothing if the organism lacks access to the habitat. For instance, a study of the ranging ability of various aquatic taxa, including taxa from bacteria to fishes, hypothesized that the patterns of occurrence in lakes for species with weaker movement abilities would be better predicted by regional patterns of connectivity, whereas the occurrence of taxa with greater movement abilities would be better predicted by local conditions (Beisner et al. 2006). Not surprisingly, the patterns of occurrence of freshwater fishes were best predicted by the spatial distribution of lakes and patterns of aquatic connectivity. In contrast, for taxa with high movement ability, such as bacteria, the local environment was a better predictor of occurrence than measures related to aquatic connectivity.
Movement of some sort is integral to a wide range of fish behavior, from finding food or mates, to avoiding or reducing predation risk, to defending feeding or breeding territories, to long distance, often annular, movements frequently associated with breeding. In the context of this chapter, the categories of interest include those that result in an organism moving outside of its home range such that new habitats could be colonized or former habitats recolonized. Other aspects of movement are treated in subsequent chapters (especially Chapters 9 and 14). Evidence for the movement abilities of fishes comes from a variety of sources including observational studies on the colonization of new habitats and from studies using some sort of tagging procedure.
Movement Inferred from Colonization Studies
Colonization of newly available habitats by fishes has been studied on a variety of temporal and spatial scales, including those studies examining movement of fishes into newly available habitats following the retreat of the Wisconsinan ice sheets, the recolonization of habitats after amelioration of harsh conditions, such as complete dewatering of a stream reach, or the response of fishes, usually over a short time period, to newly created stream habitats.
During the Wisconsinan glacial advance, the Laurentide ice sheet covered eastern Canada including Ontario (Chapter 3; Figure 3.4) so that there were no ice-free habitats available for freshwater fishes (Mandrak and Crossman 1992). As the ice began retreating, large meltwater lakes and connecting rivers formed along the southern boundary of the ice sheet. Lake Algonquin was one such meltwater lake that covered parts of modern-day Lake Huron, Lake Superior, and Lake Michigan approximately 12,000 years ago (Hinch et al. 1991; Mandrak and Crossman 1992) (Figure 5.3). The area inundated by Lake Algonquin in Ontario also included the future basins of a number of smaller lakes and, as Lake Algonquin subsided, its fish fauna would have had access to these smaller emerging lakes (Hinch et al. 1991). In addition, fishes from Lake Algonquin also colonized lakes at higher elevations that were not directly inundated by the glacial meltwater—in this case by moving up the lakes’ outlet streams that flowed into the discharge of Lake Algonquin (Figure 5.3).
Prairie streams of the central United States, on the edge of the Great Plains and Osage Plains, are often subjected to periods of intense drought, such that streams may be reduced to a few isolated pools or totally dewatered (Matthews 1988). In spite of such extreme conditions as total drying of long sections of streambed, the fish faunas of these streams tend to be surprisingly resilient and fairly quickly recolonize reaches after stream flow resumes (Matthews 1987, 1988). The fish fauna of Brier Creek, an Oklahoma tributary to the Red River arm of Lake Texoma, has been particularly well studied since the late 1960s and provides an example of the ability of fishes to recolonize newly watered habitats (C. L. Smith and Powell 1971; Ross et al. 1985; Matthews 1987; Matthews et al. 1988).
In 1980, the region of southern Oklahoma was hit by a severe drought and high temperatures, resulting in the dewatering of large stream sections and even heat death of one of the common species (Orangethroat Darter) (Matthews et al. 1982; Ross et al. 1985). In spite of such extreme conditions, by the following year the fish assemblage of Brier Creek was essentially unchanged from previous years (Ross et al. 1985), indicating that fishes had recolonized habitats once stream flow resumed. Drought returned in the summer of 1982, and a 1.5 km section of the stream in the headwaters (station 2, Figure 5.4) was totally dry by autumn and remained so until heavy rains in March of 1983 (Matthews 1987). Once flow resumed, monthly collections showed that the fish fauna was rapidly rebuilt by species colonizing from refugia outside of the dewatered reach, followed by spawning of some of the colonizers; the fish fauna of this area of Brier Creek typically includes only 9–10 species, so colonization essentially reestablished the entire fauna in approximately four months (Ross et al. 1985; Matthews et al. 1988). The favorable stream conditions were, however, short lived. Flow ceased in July, and by September the section was reduced to one pool with no surviving fish (Figure 5.4) (Matthews 1987).
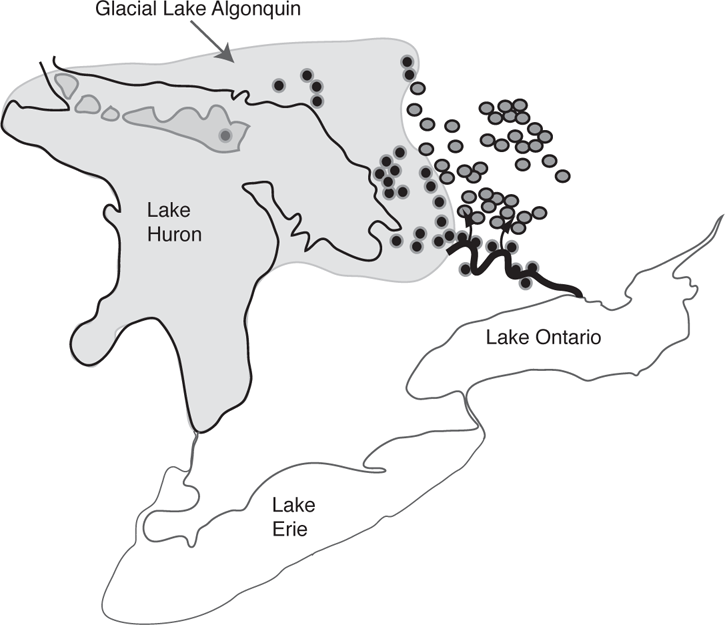
FIGURE 5.3. Dispersal from glacial meltwater lakes: Glacial Lake Algonquin in Ontario, Canada, and modern lakes in Canada and the United States. The black circles indicate lakes within the borders of Glacial Lake Algonquin or along the glacial outlet and colonized as Lake Algonquin subsided; the gray circles indicate higher elevation lakes colonized by fishes moving from the outlet of Lake Algonquin up through streams draining the lakes, as indicated by the arrows. Based on Hinch et al. (1991).
A long-term study to evaluate the impact on trout populations of new habitats created by the placement of log weirs in high-elevation streams in the Rocky Mountains included 500-m reaches of six streams and involved four species and three genera of salmonids (Brook Trout, Salvelinus fontinalis; Cutthroat Trout, Oncorhynchus clarkii; Brown Trout; and Rainbow Trout) (Gowan and Fausch 1996). Newly available habitats created by the weirs were primarily colonized by trout that were coming from outside of the 500-m study reaches; however, once the trout were in the new habitats they tended to remain there. Marked fish within the study reaches tended to be sedentary, but the presence of unmarked fish in the new habitats is a strong indication of the ability for long distance movement.
Movement Inferred from Tagging Studies
Clearly, the previous examples indicate that fishes are capable of moving through connecting waterways, but for a long time there have been questions of how far fishes routinely or periodically move and what proportion of a population is involved in movement. In 1959, Shelby Gerking summarized what was then known about movement in both marine and freshwater fishes. He identified 34 fish species (24 freshwater taxa; 10 marine taxa) characterized by restricted movement that was not associated with spawning. Following the classic work by Gerking, a view referred to as the “restricted movement paradigm” developed among fish ecologists: namely, that freshwater fishes for the most part had local populations and that extensive movement was uncommon. However, Gerking was careful to qualify his conclusions. First, he emphasized the importance of experimental design on the outcome of movement studies. Even if fishes tend to remain primarily in a home range, if the resampling area is small relative to the home range of the fish (keeping in mind that the home range is not known), then there is a bias toward finding few fish and concluding that long-range movement is common. Conversely, if the resampling area is large relative to the size of the unknown home range, then there is a bias toward finding many fish and concluding that fishes have restricted movement. Gerking (1959) was also careful to point out that restricted movement behavior was not universal among any given population and said, “That stray fish occur has never been doubted, and their importance in repopulation of decimated areas and the distribution of the species is not questioned.” In fact, Funk (1955), as cited by Gerking, proposed that lotic freshwater fishes had populations composed of a mobile group and a sedentary group (often referred to as “movers” and “stayers”). Funk (1955) also pointed out the problem of bias toward finding limited movement for fishes when sampling was concentrated in “limited areas near release sites.”

FIGURE 5.4. A. Brier Creek, a tributary to the Red River arm of Lake Texoma in Oklahoma. Numbers show sampling stations. Map based on C. L. Smith and Powell (1971) and Ross et al. (1985).
B. Recolonization of a dewatered section of Brier Creek, Oklahoma, following resumption of stream flow. Drying occurred in a 1.5 km reach at station two. Severe drought dewatered the stream again by September. Data from Matthews (1987).
Movement of freshwater fishes is generally studied by marking individuals in some way and then attempting to recapture them later (termed mark-recapture). As pointed out by Funk, sampling is almost always greater near the point of release compared to sampling at great distances from the release, especially if there are many potential routes of fish movement (as in lakes or in streams with numerous tributaries). As a consequence, sampling effort is unequal over distance from the point of release, so that short movements tend to be recorded more often than long movements. The change in sampling intensity with increasing distance from the point of release is referred to as “distance weighting” (Porter and Dooley 1993; Albanese et al. 2003). Thus unless the study compares recoveries against the probability of recapture (i.e., capture probability decreases as fishes disperse outward from the point of release) or, preferably, is designed to alleviate the issue of distance weighting, there would be strong bias for interpreting recovery data as supporting limited fish movement (Box 5.2).
BOX 5.2 • Distance Weighting in Studies of Fish Movement
An appropriate experimental design is critical for assessing fish movement using mark-recapture approaches. This is especially so because the likelihood of capturing a marked fish declines with distance from the point of release, leading to the risk of underestimating longer movements. A robust experimental design is an important issue even with essentially linear stream systems—additional complexity of the aquatic system (e.g., tributary streams, lakes with numerous coves, etc.) further increases the challenge of obtaining reliable data. Following Rodríguez (2002), in quantitative terms, the density of marked fish multiplied by meters away from the region of their release, n(x), is given by
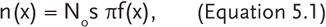
where No is the number of fish originally marked, s is the probability of their surviving to the sampling period, π is the catchability of the fish, and f(x) describes the decline in density as a function of the distance from the release area (referred to as a dispersal function). The key point that this equation makes is that the number of recaptures at any given location must be evaluated relative to the probability of recapture.
Albanese et al. (2003) evaluated the impact of distance weighting on the assessment of movement by three species of southeastern stream fishes. He used a modeling study to illustrate the impact that increasing the number of 50 m sampling sections would have on distance weighting. Fish were considered marked in ten 50 m sections. The modeling approach showed how the zone of uniform sampling (i.e., sampling at or near 100%) changed as the sampling area was increased. If sampling only occurred within the 500 m marking section, then the impact of distance weighting was extreme. The proportion of total possible movements sampled (PSd) was only 100% for sampling within ± 50 m (i.e., the zone of uniform sampling) and declined sharply for movement distances > ± 50 m. Clearly, a study design that only included sampling within the same stream reach used to mark fish would be strongly biased toward short-term movements. With a 1,000 m sampling effort, the PSd values were 100% for fish movements within ± 250 m, and with a 2,000 m sampling effort, PSd values are 100% for fish movements within ± 750 m. Both of these are much more robust designs in terms of understanding movement. However, even with the 2,000 m sampling effort, any fish movements greater than ± 1,200 m would have been undetected.
In the following figure, based on Albanese et al. (2003), the modeling study shows the proportion of total possible movements (PSd) that would be detected for three different sampling designs: one in which the sampling reaches were the same as the marking reaches (500 m), one in which the sampling reaches (1,000 m) were twice that of the 500 m marking reach, and one in which the sampling reaches (2,000 m) were four times that of the 500 m marking reach. Solid lines show PSd values; dashed lines show the sampling lengths. Movement can be either upstream (+) or downstream (−) from the marking section.
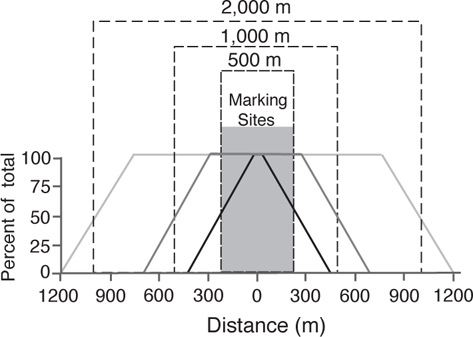
The effect of the size of the resampling areas (indicated by dashed lines) on the ability to detect marked fishes, relative to the distances that the fishes move. As determined by a modeling study, the largest resampling area (2,000 m) can detect 100% of fish movements up to 750 m upstream or downstream from the marking site (indicated by shading). The smallest sampling area (500 m and the same as the marking area) could only detect 100% of fishes that moved less than approximately 50 m. Based on Albanese et al. (2003).
In theory, the probabilities of capture from distance weighting could be used to adjust the observed captures of fish (see Albanese et al. 2003). In actuality, outside of the zone of uniform sampling, the numbers of fish captured were so low that adjustments generally were not possible—adjustments are not possible if no fish are captured! The take-home message is that an understanding of distance weighting using PSd values is most useful for the a priori design of the sampling study. The a posteriori application of correction factors generally cannot correct for a poor study design.
In spite of the cautionary words by Gerking (1959) about the problem of experimental design and the presence of fish straying, and the suggestion by Funk (1955) about sedentary and mobile groups, the restricted movement paradigm became entrenched in the literature. One of the first to take issue with the restricted movement paradigm, specifically in regard to salmonids, was Gowan et al. (1994), who pointed out that although most tagging studies captured the majority of fish near the point of release, in 78% of the salmonid studies that they reviewed, over half of the fishes were never seen again after being marked. Whether these fish represented mortalities or fish that simply moved much greater distances is the crux to understanding the level of movement of fishes. Gowan et al. (1994) also suggested that the mobile fraction of fish populations had been downplayed through the use of such deprecating terms as “strays” and argued that more attention needed to be given to the experimental design of fish movement studies and to the underlying mechanisms involved in fish movement. The greater realization of the often high degrees of movement shown by freshwater fishes has had important consequences for the better understanding and management of fish populations (Fausch et al. 2002).
A study of fish movement in a small Ouachita Highlands stream (Arkansas) involved four species of stream fishes (Creek Chub, Semotilus atromaculatus; Blackspotted Topminnow, Fundulus olivaceus; Green Sunfish, Lepomis cyanellus; and Longear Sunfish, L. megalotis) (Smithson and Johnston 1999). The study area was 500 m long and consisted of 10 pools, and the possibility of movement of fishes outside of the study area was determined to be unlikely. Most fishes were recaptured in the same pool where they were initially marked; however, there were differences among the species. Compared to the other three species, Blackspotted Topminnow moved significantly greater distances. For all species, there were some individuals that moved greater distances than others, although there were no apparent morphological correlates associated with greater movement and, in fact, the same individuals switched between static and mobile behaviors over the course of the study. This suggests that individual fish periodically engage in exploratory travel, perhaps assessing habitat quality in areas outside of their home pool.
Field and theoretical approaches were used by Skalski and Gilliam (2000) to examine characteristics of movement of primarily four species (Bluehead Chub, Nocomis leptocephalus; Creek Chub; Redbreast Sunfish, Lepomis auritus; and Rosyside Dace, Clinostomus funduloides) in a small southeastern stream. In contrast to Smithson and Johnston (1999), species showed only weak differences in their degree of movement. However, within a species there was evidence for both “movers” and “stayers.” In addition, the distribution of distances moved tended to be leptokurtic, with more short and long movements and fewer intermediate movements compared to a normal distribution. Relationships between the propensity to move and morphological characteristics were complex and varied among species. Within Bluehead Chub, the probability of movement increased with size for individuals that had slow growth but decreased with size for those having fast growth. In Creek Chubs the probability of movement increased with body size but was not related to growth rate. There was no relationship between body size and the probability of movement in Redbreast Sunfish or Rosyside Dace. Correcting the movement data to account for distance weighting had little effect. However, this does not negate the strong potential effect of distance weighting but emphasizes the importance of an appropriate sampling design that, a priori, deals with the problem of distance weighting—a conclusion also reached by Albanese et al. (2003) (Box 5.2).
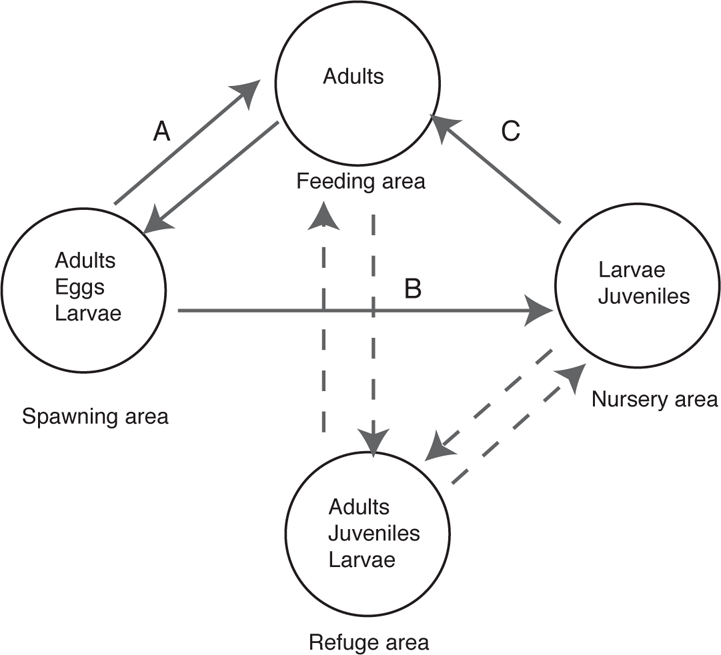
FIGURE 5.5. Patterns of migration in fishes. The circles enclose life-history stages using a particular resource. Solid lines indicate regular movement; dashed lines indicate aperiodic movement to refuge areas when there are harsh environmental conditions. Distances (A, B, C) between circles can vary greatly from a few meters to hundreds or thousands of kilometers. Adapted from Harden Jones (1968) and Schlosser (1995).
A general pattern that emerges from these studies of fish movement is that although the majority of fishes are often sedentary, there is often another, albeit smaller, group that undertakes much more extensive movements. Although use of the terms “movers” and “stayers” is descriptively appealing, several studies, including Smithson and Johnston (1999), have shown that the same individuals may switch from static to mobile behavior. In other words, fishes are continually responding to their local environment relative to their physiological needs, and the variation in when and how much they move reflects this. Hence a “mover” one day might be a “stayer” the next.
Ontogeny and Movement
During their life cycle, freshwater fishes are faced with many challenges, including feeding, growth, predator avoidance, and reproduction. In some species and/or habitats, these activities may occur over a small spatial scale on the order of meters, whereas in others, tens or hundreds of kilometers may separate these and other activities (Figure 5.5). An important point that Figure 5.5 illustrates is that movement can occur at any life-history stage. For egg and larval stages, movement is generally passive and occurs via transport by water currents. The downstream drift of fish eggs and/or larvae is widespread, but by no means universal, among freshwater fish taxa (Gale and Mohr 1978; Brown and Armstrong 1985; Pavlov 1994). Entry into the drift by larval fishes can be due to turbulence that dislodges larvae (termed catastrophic drift) or because of active choice, as larvae swim up off the bottom. As described by Pavlov (1994), once in the water column, drift can be passive (most common for early larval stages), drifting downstream but not oriented to the direction of flow. Drift can also be active, where fish are actively swimming downstream, or active-passive, where fish show orientation to the current but only weak swimming ability. Several examples of species from different regions and habitat types in North America serve to illustrate these patterns.
MOVEMENT AT THE FERTILIZED EGG STAGE Various minnow species in Great Plains streams have adapted to life in large, turbid rivers with shifting sand substrata by having a semibuoyant egg stage. This is apparently an adaptation to the unpredictable summer flows, since eggs are released during high-discharge periods (Platania and Altenbach 1998; Dudley and Platania 2007), and perhaps to avoid the risk of suffocation by sediment accumulation that would threaten eggs deposited directly on a shifting substratum. In the Rio Grande drainage (including the Rio Grande and Pecos rivers), four native species in three genera form a reproductive guild of broadcast spawners with semibuoyant eggs. The species are the Rio Grande Silvery Minnow, Hybognathus amarus; Speckled Chub, Macrhybopsis aestivalis; Rio Grande Shiner, Notropis jemezanus; and Pecos Bluntnose Shiner, N. simus pecosensis (Platania and Altenbach 1998). Two additional taxa that were endemic to the Rio Grande (Phantom Shiner, Notropis orca; Rio Grande Bluntnose Shiner, Notropis simus simus) were also likely members of this guild; unfortunately these taxa are now extinct (Bestgen and Platania 1990; Platania and Altenbach 1998).
Embryonic and early larval development of fishes in this guild occurs as they drift downstream with river flow. The distances required for the egg and early larval development to occur are impressive—during the time of passive transport of the eggs, they could travel some 144 km. The newly hatched protolarvae, which also remain in the water column, could be carried an additional 216 km depending on water temperature (which controls the rate of development) and current speed (Platania and Altenbach 1998). Clearly, this guild of Rio Grande fishes shows that the eggs and early larval stages of certain species have the ability to move great distances. Upstream movements of juveniles and adults counter the downstream movements of the eggs and protolarvae. In relation to Figure 5.5, the distance (A) from adult habitat to spawning habitat can be quite short. In contrast, the distance (B) from the spawning habitat (open water) to the nursery habitat (downstream in shallow, slow water along the shoreline) can be a hundred or more kilometers. This occurs following the protolarval stage. Later larval stages and juveniles inhabit the shallow, warm, and productive river margins as they gradually move upstream. This upstream distance (C) is also on the same scale as the distance from the spawning habitat (B). This life-history pattern, while demonstrating the ability for long distance movement of early life-history stages and adults, is obviously very susceptible to man-made barriers in rivers and to flow modifications (Winston et al. 1991; Dodds et al. 2004; Dudley and Platania 2007). The semibuoyant eggs require at least some current speed to remain in suspension. Consequently, if they enter the slack water of a reservoir they tend to sink and die from suffocation. The high concentration of nonnative predators in reservoirs is also generally lethal to drifting eggs and larvae (Dudley and Platania 2007). Dams also preclude the upstream return movement of juveniles and adults. As a consequence, reproductive output of most of the breeding adults of silvery minnows seems to be lost as developing embryos and larvae drift into impoundments (Alò and Turner 2005); it is no surprise that most species of this guild are extirpated or have their ranges greatly reduced and have required federal listing as threatened or endangered.
MOVEMENT AT THE LARVAL STAGE In contrast to the previous examples of fishes with semibuoyant eggs, most freshwater fishes have eggs that are demersal and adhesive, remaining attached to bottom materials such as gravel, sand, wood, or other solid materials prior to hatching. However, larvae of many species do have a free-swimming stage and can enter the water column of streams. The diverse arrays of invertebrate and vertebrate organisms that are carried in the water column are collectively referred to as drift. The drift of larval fishes can occur at any larval stage but is most prevalent at the earliest (protolarvae) and the intermediate (mesolarvae) larval stages and can result in downstream movement on the scale of meters to hundreds of kilometers. Relative to Figure 5.5, distances of larval drift (B) and movement of juveniles to adult feeding areas (C) are generally roughly equivalent to distances moved by adults to the spawning ground (A). The duration of larval drift varies widely among taxa, from very long periods of drift in various species of minnows, as discussed previously, to very short time periods in the drift, as in certain species of darters (Slack et al. 1998). The density of drifting fish larvae can be impressive. In the Smith River, a coastal river of northern California, White and Harvey (2003) found that some 2.5 billion sculpin (Cottus) embryos and larvae move down the river to the estuary over a four-month period.
Movement via larval drift is common in many North American fish families. Of the 15 families that make up 90% of North American species (Chapter 1; Table 1.1), larval drift or drifting by early juvenile stages commonly occurs in 10 families (Table 5.1). Exceptions to this are livebearing fishes (families Poeciliidae and Goodeidae), fishes in the families Cyprinodontidae and Fundulidae where larvae tend to remain on or near the bottom (Foster 1967), and fishes where young are often guarded in a confined nesting cavity, such as madtom catfishes (Noturus), or closely guarded by parents (family Cichlidae).
The numerically dominant cyprinids are widely represented in drift throughout North American streams, including the western and central United States (Robinson et al. 1998), northern rivers in Canada and the United States (Muth and Schmulbach 1984), and southeastern streams (Gallagher and Conner 1980; Slack et al. 1998). As shown in the previous section for species with drifting eggs and larvae, distances traveled by larvae can exceed several hundred kilometers. In the Colorado River drainage, estimates of movement of native cyprinid larvae range from approximately 9 km for Humpback Chub (Gila cypha) and Speckled Dace in the Little Colorado River (Robinson et al. 1998), to over 200 km for Colorado Pikeminnow (Ptychocheilus lucius) in the Green River (Tyus and Haines 1991).
For percids, larvae of the commercially and recreationally important Walleye (Sander vitreus) and Yellow Perch (Perca flavescens) exhibit substantial drift with greatest abundances occurring at night (Gale and Mohr 1978; Corbett and Powles 1986; Johnston et al. 1995). The occurrence in drift samples of darter species in the genus Etheostoma has been documented in several studies, with peak abundances typically at night from 2100 to 0300 h (Gale and Mohr 1978; Lathrop 1982; Brown and Armstrong 1985; Paller 1987). Even Bayou Darters (Nothonotus rubrum), members of a genus that inhabits swift water and coarse substrata, show downstream drift of at least several hundred meters. This is at least far enough for them to travel between patches of riffle habitats that juveniles and adults of this species selectively occupy (Slack et al. 2004). Larval drift also occurs within the darter genus Percina. For instance, larval Snail Darters (Percina tanasi) show downstream transport of up to several kilometers, followed by return upstream movement of juveniles and adults (Kuehne and Barbour 1983). As shown earlier, this type of life cycle that includes downstream drift followed by upstream movement of juveniles and adults is greatly at risk from man-made barriers to movement. Indeed, the population of Snail Darters in the Little Tennessee River was extirpated by the infamous Tellico Dam (Ono et al. 1983). More recently, additional populations of Snail Darters have been discovered in several Tennessee River tributaries, although of the nine known populations, six are considered marginal (Williams et al. 1989).
TABLE 5.1 Prevalence of Larval and Early Juvenile Drift in Numerically Dominant Freshwater Fish Families of North America
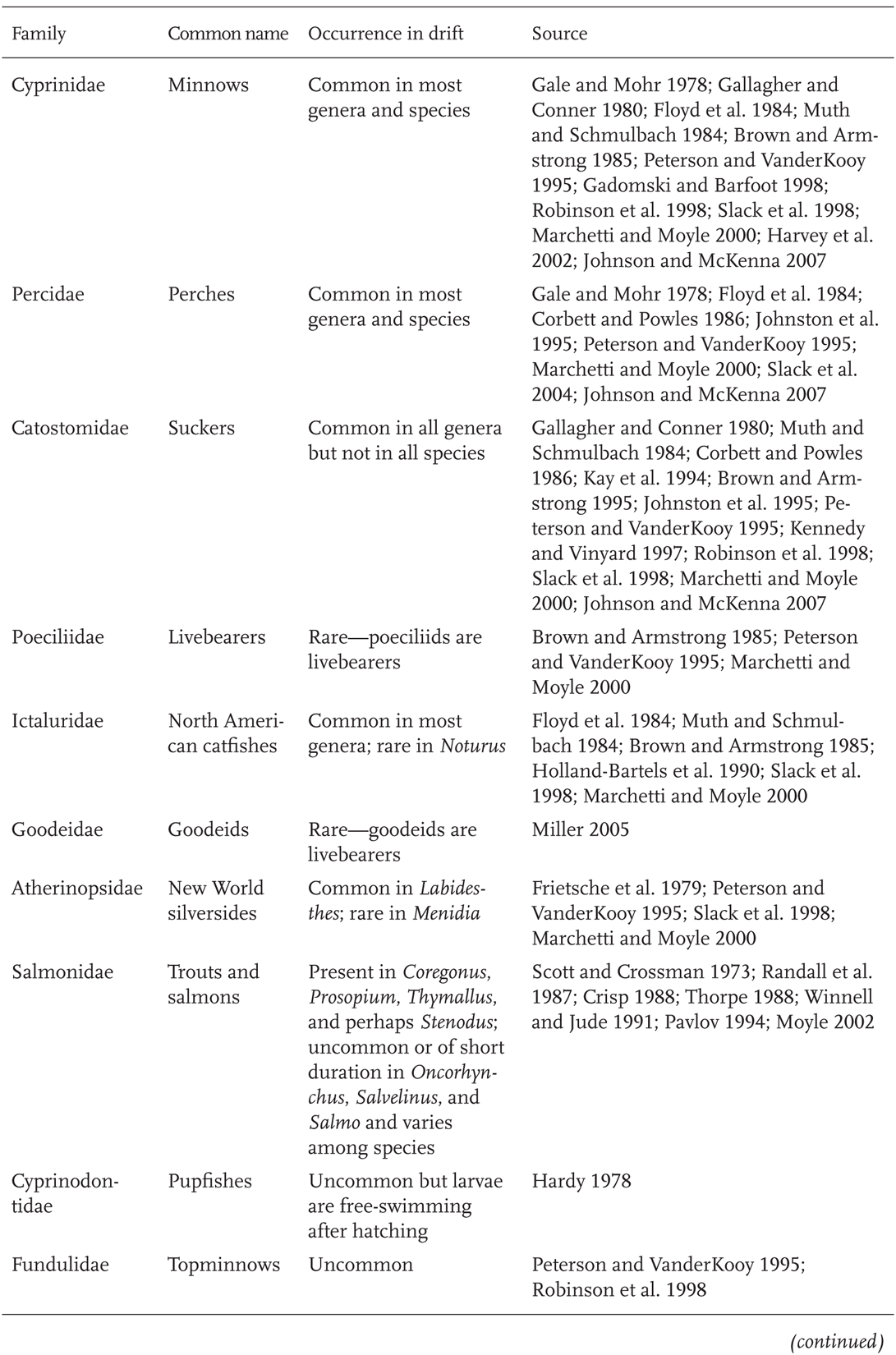
TABLE 5.1 (continued)
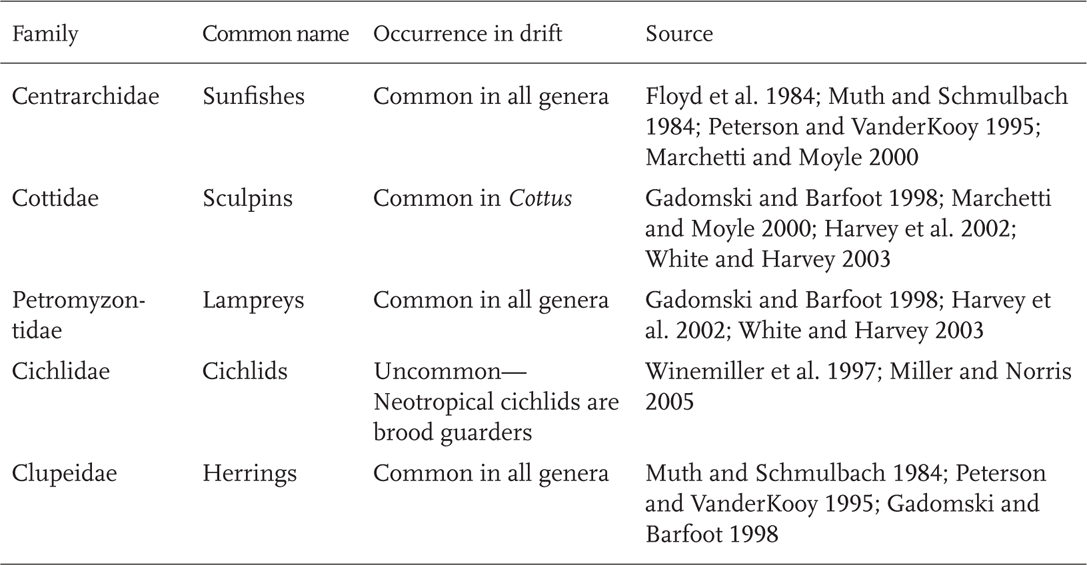
Catostomids, the third most speciose North American fish family, are also well represented in larval drift. Historically, many western catostomids drifted long distances downstream from spawning areas, subsequently followed by upstream spawning movements of adults. In the Little Colorado River, Bluehead (Catostomus discobolus) and Flannelmouth (C. latipinnis) sucker larvae drifted at least 9 km downstream from the spawning area (Robinson et al. 1998). Another Colorado River endemic, the Razorback Sucker (Xyrauchen texanus) has even more extensive movements as larvae. Because of the close linkage of adult and larval distances, Razorback Suckers, along with other examples of larval drift, are included in the following section on adult movement.
MOVEMENT AT THE ADULT STAGE In the Great Lakes of North America, Lake Sturgeon (Acipenser fulvescens) adults make spawning migrations out of the lake habitats and into tributary rivers. For example, in Lake Superior, adult sturgeon travel upstream for spawning at a single riffle in the Sturgeon River, Michigan, a distance of 69 km (Auer and Baker 2002). After hatching, larvae drift downstream at least 45 km and in some cases 61 km. In reference to Figure 5.5, the distance from adult feeding area to the spawning area (A) is 69 km; the nursery area is essentially the lower 10 km of river habitat downstream of the spawning site, and so this distance (B) is approximately 59–69 km; the other distance (C) would include the movement within Lake Superior to juvenile and adult feeding grounds.
Most suckers (family Catostomidae) also exhibit seasonal spawning migrations in which they move upstream into small tributaries from larger streams or lakes. Depending on the species, distances moved vary greatly, and movement may occur in groups or in larger schools (Curry and Spacie 1984) (see also Chapter 14). Adults make the return downstream migration after spawning, whereas newly hatched larvae are passively transported downstream by water flow, with larval numbers often increasing in surface waters at night (Gale and Mohr 1978).
A striking example of extensive travel to spawning habitats by a catostomid is provided by the Razorback Sucker, a species endemic to the Colorado River system. In the Green River and its tributaries, movements of Razorback Suckers are bounded upstream by the Flaming Gorge Dam and downstream by Lake Powell and the Glen Canyon Dam. Within this reach there are two spawning sites known for Razorback Sucker—one in the Yampa River upstream from its confluence with the Green River, and one in the Green River downstream of the Yampa River (Tyus and Karp 1990; Modde et al. 1996; Figure 5.6). Fish in breeding condition may travel at least 30–106 km to reach the spawning sites, followed by equivalent downstream movement (Tyus and Karp 1990). After hatching from demersal, adhesive eggs, larvae drift downstream into nursery habitat (historically provided by large backwaters) (Modde et al. 2001). Another Colorado River endemic, the Colorado Pikeminnow, makes equally impressive long distance spawning movements (Tyus and McAda 1984).
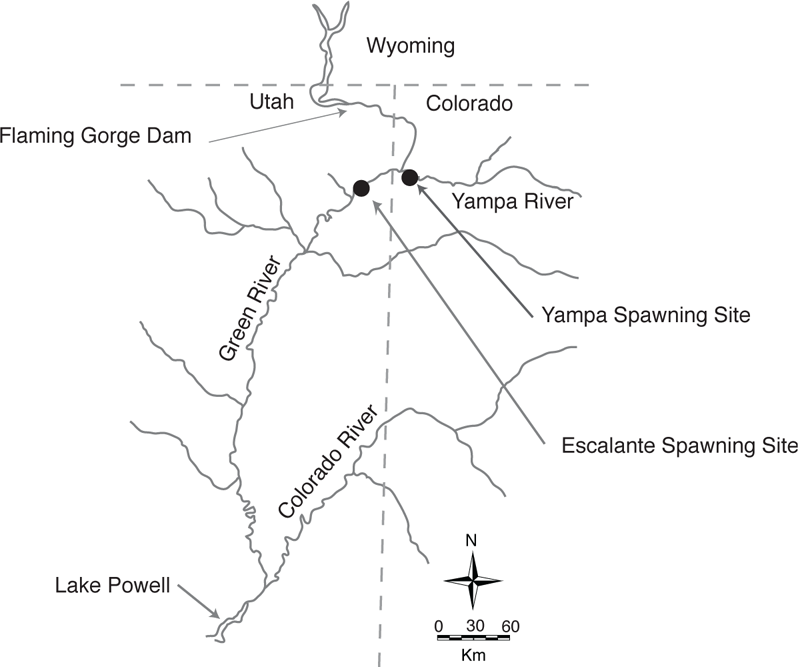
FIGURE 5.6. Spawning movements in fishes as illustrated by the Razorback Sucker, (Xyrauchen texanus), an endemic catostomid in the Colorado River drainage. Spawning locations of Razorback Sucker are indicated by black dots. Based on Tyus and Karp (1990), Modde et al. (1996), and Modde and Irving (1998).
The most impressive long-distance movements occur in fishes that travel between salt and fresh water for purposes of spawning (diadromy) (see also Chapter 9). In fishes that spawn in fresh water and then spend part of their life in the sea where they feed (anadromy), one-way distances traveled can be hundreds or even thousands of kilometers. For instance, Chinook Salmon (Oncorhynchus tshawytscha) travel almost 2,000 km as the spawning adults move from the Pacific Ocean upstream to spawning sites in the Yukon River (corresponding to distance A in Figure 5.5) (Scott and Crossman 1973). Post-yolk-sac fish (fry) as well as parr (young salmonids during the first year or two of life) and smolts (older juveniles ready to return to the sea) make the return journey downstream and then out to sea (distances B and C in Figure 5.5). Once in the open ocean where they are actively feeding, Pacific salmon may travel over thousands of kilometers during that time (usually 1–6 years) they spend at sea (Healey and Groot 1987; Thorpe 1988; Walter et al. 1997).
SUMMARY
The process of forming fish assemblages, although complex, involves characteristics of the environment, characteristics of the fish species in the regional species pool, and characteristics of the fishes and other biota in the local environment. Fish assemblages tend to be structured rather than random groupings of species, although random processes may at times be important. Also, very few studies representing even fewer geographical regions have rigorously addressed the issue of structure in freshwater fish assemblages. A major factor seems to be the fit of the potential colonizer with the environmental features of the new habitat. Following this, trophic position (low or high rather than intermediate), and if there is parental care of young, are important attributes of successful colonizers.
Fishes show the ability to move long distances, and depending on the species, movement may occur at any life-history stage. Even within fish populations that are relatively sedentary, individuals may make periodic or aperiodic movements, most likely in response to assessing resource availability or the risk of predation. Although the terms movers and stayers have been used to describe the differences in movement among individuals in a population, data seem to indicate that the same individual can shift between the two states. Hence the terms apply more to the state of an individual rather than differences among individuals. As long as there are periodic water connections and sufficient time, the well-developed capability for movement in most fish taxa allows fishes to colonize new areas or to enter preexisting assemblages.
SUPPLEMENTAL READING
Albanese, B. W., P. L. Angermeier, and C. Gowan. 2003. Designing mark-recapture studies to reduce effects of distance weighting on movement distance distributions of stream fishes. Transactions of the American Fisheries Society 132:925–39. Explores the role of changing capture probabilities with distance from the release point in estimates of fish movement.
Belyea, L. R., and J. Lancaster. 1999. Assembly rules within a contingent ecology. Oikos 86:402–16. An overview of the literature on community assembly.
Fausch, K. D., C. E. Torgersen, C. V. Baxter, and H. W. Li. 2002. Landscapes to riverscapes: Bridging the gap between research and conservation of stream fishes. BioScience 52:483–98. Emphasizes the need to view streams and rivers as complex landscapes and the importance of access to these varied habitats by fishes.
Gowan, C., M. K. Young, K. D. Fausch, and S. C. Riley. 1994. Restricted movement in resident stream salmonids: A paradigm lost? Canadian Journal of Fisheries and Aquatic Sciences 51:2626–37. A reanalysis and counter argument to the restricted movement paradigm.
Mandrak, N. E., and E. J. Crossman. 1992. Postglacial dispersal of freshwater fishes into Ontario. Canadian Journal of Zoology 70:2247–59. An important paper on the glacial refugial origins of Ontario freshwater fishes.
Taylor, C. M. 1996. Abundance and distribution within a guild of benthic stream fishes: Local processes and regional patterns. Freshwater Biology 36:385–96. Uses field collections and manipulative studies to test predictions of hypotheses regarding the abundance and distribution of fishes.