9
The evolution of selfless behaviour
Altruism is one of evolution’s trickiest puzzles, and the question of why animals are so nice to one another is fiercely contested. If you accept that evolution is all about selfish genes, then what role has the group to play? Survival of the fittest means survival of the fittest DNA, which makes you and I mere vehicles in which our genes are hitching a lift on the road to posterity. Or maybe not…
The origin of altruism
The theory of natural selection clearly explains how features such as the sharp teeth of the tiger, the thick fur of the polar bear and the camouflage of the moth evolved. Many social species, however, have traits that benefit others or the group as a whole, often to their own cost. It is much harder to see how such traits evolve. Darwin proposed that it was a result of the survival of the fittest groups, rather than the fittest individuals. Group selection, as this idea is known, has a long and complicated history and is one of the most controversial issues in evolutionary biology today. In ‘What makes animals altruistic’ (below), David Sloan Wilson, a leading proponent of group selection outlines the history of the field, and makes the case why group selection is a powerful force in evolution. There is also an interview with sociobiology pioneer and group-selection proponent Edward O. Wilson. Later in the chapter, we hear why other biologists disagree, and Richard Dawkins explains why his gene-centred view of evolution is sufficient to explain altruism.
What makes animals altruistic?
Many animals help other members of their group, often to their own cost. Evolutionary biologist David Sloan Wilson explains how such traits evolve, and makes the case for group selection.
Wolves share food with other members of the pack. Vervet monkeys make alarm calls that risk attracting a predator’s attention to themselves. Bees sacrifice themselves to defend a hive. But why? It is hard to see how such traits evolve by natural selection because individuals that behave in these ways would seem to have far less chance of surviving and producing offspring than more selfish members of the same group.
Darwin himself was acutely aware that the suicidal sting of the honeybee and most of the virtues associated with human morality, such as bravery, honesty and charity, posed a severe challenge to his theory. ‘It must not be forgotten that…a high standard of morality gives but a slight or no advantage to each individual man and his children over other men of the same tribe,’ he wrote in The Descent of Man (1871).
The question, then, is how can traits that are oriented toward others or one’s group as a whole – known as ‘prosocial’ traits – evolve when they appear to reduce the relative fitness of individuals within groups?
A simple solution
Darwin realized that the problem could be solved if there was selection at a group level, not just at an individual level.
If groups composed of individuals who behave in a more prosocial way outcompete groups of individuals who behave in a less prosocial way, then traits that are for the good of the group will evolve. In short, natural selection between groups will counteract the costs of prosocial behaviour to individuals within groups.
Darwin’s insight was the starting point for the modern theory of multilevel selection. According to this theory, biological systems are a nested hierarchy of units, from genes within individuals, individuals within groups, groups within a population and even clusters of groups.
At every level, the traits that maximize relative fitness within a unit are unlikely to maximize the fitness of the unit as a whole. Genes that outcompete other genes within the same organism are unlikely to benefit the whole organism. Individuals that outcompete other individuals within their group are unlikely to benefit the group as a whole, and so on.
FIGURE 9.1 Groups that can work together successfully will have an advantage over other groups.
A complex history
In the first part of the twentieth century, the idea of group selection was accepted by most evolutionary biologists. In fact, it was embraced rather too eagerly and uncritically. Influenced by the lingering idea that nature was the creation of a benign god, many biologists thought that nature was adaptive at all levels – what is good for individuals must be good for groups, and so on. As a 1949 biology textbook put it: ‘The probability of survival of individual living things, or of populations, increases to the degree to which they harmoniously adjust themselves to each other and to their environment.’
While some biologists realized that the evolution of prosocial traits would often be opposed by selection within groups, it was frequently assumed that group selection would usually prevail – a position that has come to be known as ‘naive group selectionism’. It was typified by British zoologist Vero C. Wynne-Edwards, who in 1962 proposed that organisms evolve to regulate their population size to avoid overexploiting their resources.
When the idea of group selection came under scrutiny in the 1960s, naive group selection was rightly rejected. But the backlash did not stop there. Based on arguments about how plausible it is, rather than actual experiments and studies, a consensus formed that between-group selection is almost invariably weak compared with within-group selection. As evolutionary biologist George C. Williams put it in his highly influential book Adaptation and Natural Selection: ‘Group-related adaptations do not, in fact, exist.’
This was a major change in thinking, and the rejection of group selection came to be regarded as a watershed in the history of evolutionary thought. For the ensuing decades, most evolutionary biologists interpreted social adaptations as forms of self-interest that could be explained without invoking group selection. In his 1982 book The Extended Phenotype, Richard Dawkins went so far as to compare efforts to revive group selection to the futile search for a perpetual motion machine.
Inclusive fitness and selfish genes
The rejection of group selection in the 1960s meant biologists had to come up with alternative theories to explain the evolution of social adaptations. Several were put forward, including inclusive fitness theory – also known as kin selection – selfish gene theory and evolutionary game theory.
The kin selection revolution started in a pub in the mid-1950s. Biologist J.B.S. Haldane was asked if he would give his life to save his brother. A few scribbled calculations later, he provocatively replied that he would only die for at least two brothers, or eight cousins.
Why? Because a gene coding for such altruism can only survive if it leaves enough copies of itself in relatives. Human siblings share on average half of their genes, and cousins one-eighth. Hence, two siblings, or eight cousins equal one self. This idea – that animals are more likely to show altruistic behaviour towards individuals they are related to – is called kin selection.
Haldane’s colleague William Hamilton later drafted a mathematical description of the phenomenon, known as inclusive fitness, which assigns numerical values to the costs and benefits of an animal’s actions. In theory, inclusive fitness makes it possible to calculate the extent of the spread of a given altruistic behaviour – such as staying with your parents to raise your siblings – through a population.
A revolution was ignited. Hamilton’s maths has been used for decades by biologists studying cooperation in animals. Group selection mechanisms no longer appeared to explain altruism. The individual came to be seen as the protagonist of natural selection. Soon the gene took its place. Bodies could be regarded as merely the genes’ way of making more genes, as famously expressed in 1976 by Richard Dawkins’s metaphor of the selfish gene.
Yet the following decades saw a revival of group selection. What brought about such a remarkable change?
The rejection of group selection was based on the claim that, in practice, selection within groups always beats selection between groups. However, studies have shown that traits can evolve on the strength of between-group selection, despite being disadvantageous for individuals within each group.
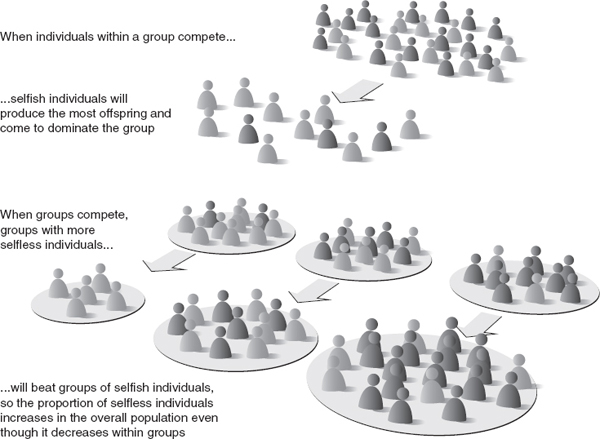
FIGURE 9.2 What happens when groups of selfish and selfless individuals compete?
Take the water strider Remigis aquarius, an insect species that skates on the surface of quiet streams. Males vary greatly in their aggressiveness toward females, and lab studies show that within any group, aggressive males outcompete non-aggressive males for females.
However, the aggressive males also prevent females from feeding and can injure them, which results in a group with lots of aggressive males producing fewer offspring than groups with fewer aggressive males. Variation among groups is magnified by females fleeing groups containing lots of aggressive males and aggregating in groups with non-aggressive males. So these studies show that between-group selection is essential for maintaining non-aggressive males in the population.
In another experiment, a team of microbiologists grew E. coli in wells on plates. They then infected some of them with a virus, and mimicked the natural spread of viruses by using robotic pipettes to transfer them between wells. The team found that in some circumstances, a ‘prudent’, slow-growing strain of the virus was more successful than a ‘rapacious’, fast-growing strain. The rapacious strain often killed off all the bacteria in a well – and therefore itself – before it had a chance to spread. The prudent strain persisted for longer and so was more likely to get a chance to colonize other wells.
In this way, the prudent strain could remain in the population even though it was always outcompeted by the rapacious strain when both were present in a single well. In other words, it was only on the strength of between-group selection that the prudent strain survived.
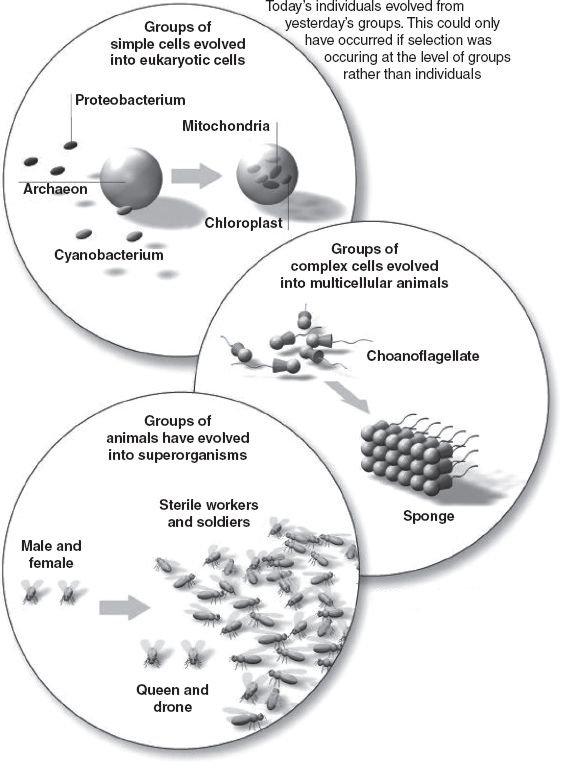
FIGURE 9.3 The evolution of today’s individuals from yesterday’s groups.
The conditions of this experiment closely resemble a scenario proposed by Vero C. Wynne-Edwards in the 1960s for the evolution of reproductive restraint in many species. Such restraints might not evolve in all species, but as this experiment shows, it is plausible that they can evolve in some species in some circumstances.
These two experiments involve very different spatial and temporal scales, but they embody the key problem and simple solution of group selection: the traits that benefit the whole group are not advantageous for individuals within the group and so require an additional layer of natural selection to evolve.
Darwin’s problem is an unavoidable fact of life for all species, including our own: prosocial adaptations usually put individuals at a disadvantage relative to other members of their group. The only way for them to evolve is if there is another layer to the process of natural selection. That layer is group selection.
Major transitions in evolution
Until the 1970s, evolution was thought to take place entirely on the basis of the accumulation of mutations over many generations. Then biologist Lynn Margulis proposed that complex cells did not evolve by small mutational steps from bacterial cells, but from symbiotic associations of different kinds of bacteria that became higher-level organisms in their own right.
In the 1990s, evolutionary biologists John Maynard Smith and Eörs Szathmáry proposed that similar major transitions occurred throughout the history of life, including the evolution of the first cells, the advent of multicellular organisms and the development of social insect colonies (see Figure 9.3). They even suggested it could explain the origin of life, with groups of cooperating molecular reactions coming together to create the first life forms. The realization that evolution takes place not only by small mutational steps but also by groups of organisms turning into higher-level organisms represents one of the most profound developments in evolutionary thought. Today’s individuals are yesterday’s groups.
For a major evolutionary transition to occur, there has to be a shift in the balance between within-group and between-group selection. A group can only turn into an individual when between-group selection is the primary evolutionary force, and this in turn can happen only when mechanisms evolve that suppress selection within groups. The rules of meiosis, for example, ensure that all genes on the chromosomes have an equal chance of being represented in the gametes. If genes can’t succeed at the expense of each other, then the only way to succeed is collectively as a group.
Major evolutionary transitions are rare events in the history of life but they have momentous consequences, as the new super-organisms become ecologically dominant. Eusociality in insects only originated about a dozen times – including in ants, bees, wasps and termites – but insect colonies comprise well over half the biomass of all insects.
These transitions are never complete as selection within groups is only suppressed, not eliminated. Some genes do manage to bias the rules of meiosis in their favour. Increasingly, cancer is studied as an evolutionary process that takes place within individuals, causing some genes to succeed at the expense of others, with tragic results for the group as a whole.
Interview: From altruism to a new Enlightenment
E.O. Wilson is the father of the field of sociobiology. In a 2012 interview in New Scientist he argued that group selection is the main driver of evolution. He is professor emeritus at Harvard University. Among his 25 books are the groundbreaking Sociobiology (1975), Consilience (1998), and the Pulitzer prize-winners On Human Nature (1978) and The Ants (1990).
In 2010 you were involved in a high-profile academic row over what drives the evolution of social traits such as altruism. Why should non-specialists care?
Scientific advances are now good enough for us to address coherently questions of where we came from and what we are. But to do so, we need to answer two more fundamental questions. The first is why advanced social life exists in the first place and has occurred so rarely. The second is what are the driving forces that brought it into existence.
Eusociality, where some individuals reduce their own reproductive potential to raise others’ offspring, is what underpins the most advanced form of social organization and the dominance of social insects and humans. One of the key ideas to explain this has been kin selection theory or inclusive fitness, which argues that individuals cooperate according to how they are related. I have had doubts about it for quite a while. Standard natural selection is simpler and superior. Humans originated by multilevel selection – individual selection interacting with group selection, or tribe competing against tribe. We need to understand a great deal more about that.
How will a better understanding of multilevel selection help?
We should consider ourselves as a product of these two interacting and often competing levels of evolutionary selection. Individual versus group selection results in a mix of altruism and selfishness, of virtue and sin, among the members of a society. If we look at it that way, then we have what appears to be a pretty straightforward answer as to why conflicted emotions are at the very foundation of human existence. I think that also explains why we never seem to be able to work things out satisfactorily, particularly internationally.
So it comes down to a conflict between individual and group-selected traits?
Yes. And you can see this especially in the difficulty of harmonizing different religions. We ought to recognize that religious strife is not the consequence of differences among people. It’s about conflicts between creation stories. We have bizarre creation myths and each is characterized by assuring believers that theirs is the correct story, and that therefore they are superior in every sense to people who belong to other religions. This feeds into our tribalistic tendencies to form groups, occupy territories and react fiercely to any intrusion or threat to ourselves, our tribe and our special creation story. Such intense instincts could arise in evolution only by group selection – tribe competing against tribe. For me, the peculiar qualities of faith are a logical outcome of this level of biological organization.
Can we do anything to counter our tribalistic instincts?
I think we are ready to create a more human-centred belief system. I realize I sound like an advocate for science and technology, and maybe I am because we are now in a techno-scientific age. I see no way out of the problems that organized religion and tribalism create other than humans just becoming more honest and fully aware of themselves. Right now we’re living in what Carl Sagan correctly termed a demon-haunted world. We have created a Star Wars civilization but we have Palaeolithic emotions, medieval institutions and godlike technology. That’s dangerous.
From all these big questions to the smallest creatures…I cannot interview the world’s best known ant expert without asking: do you have a favourite?
I do. It’s an ant called Thaumatomyrmex. In all my travels, I’ve only seen three. They’re very rare. It has teeth on jaws that look like a pitchfork. The teeth are extremely long, and when it closes the jaws, they overlap. In at least one species, the teeth actually meet behind the head. So what does this monster eat? What does it use those teeth for? I just had to know, so I sent an appeal out to younger experts in the field, particularly in South America, where these ants are found.
Eventually they discovered the answer: it feeds on polyx-enid millipedes. These millipedes have soft bodies, but they’re bristling all over like a porcupine. So the ant drives a spike right through the bristles and nails it. And what we hadn’t noticed is that the ant also has thick little brushes [on some of its limbs], and members of the colony use these to scrub the bristles off – like cleaning a chicken – before dividing it up. That’s my favourite.
Sparks fly over the origin of altruism
To the average person, it might be hard to understand why biologists get so worked up about the origins of altruistic behaviours. But arguments about this topic are perhaps the most heated in science.
Since the evolution of altruistic traits such as forgoing reproduction in order to help raise another’s offspring, has been explained by kin selection: the idea that helping your relatives – and therefore helping spread the genes you share with them – outweighs the cost of not having offspring of your own. It’s the genes that matter, not the individuals in which they reside or the groups in which those individuals live. The idea was encapsulated in Richard Dawkins’s pervasive metaphor of the selfish gene.
So when David Sloan Wilson and others revived the idea of group selection, it invoked a furious response from biologists in the gene-centred Dawkins camp. More fury was to come when, in 2008, E. O. Wilson wrote a paper outlining why kin selection was not the decisive factor in the evolution of sterile workers. He argued that once fully eusocial colonies (where a mother ‘queen’ is assisted at the nest by non-reproducing offspring) had developed, they continued to evolve by means of group selection, because groups that cooperate do better than those that do not.
This too, prompted a scathing response from many other biologists. Richard Dawkins, for example, in a comment article for New Scientist picked holes in Wilson’s ‘misleading’ group-selection terminology, and argued that ‘what really matters is gene selection’ (also, see interview later in this chapter). He ended his article with these words: ‘Evidently Wilson’s weird infatuation with “group selection” goes way back: unfortunate in a biologist who is so justly influential.’
The arguments rolled on. In the 2010 meeting at the Royal Netherlands Academy of Arts and Sciences in Amsterdam, for example, one theoretical biologist denounced three of his colleagues as ‘unscholarly’ and ‘transparently wrong’, and wondered what could have led such ‘talented, honest biologists’ to be so ‘misguided’.
The meeting was on the evolution of conflict and cooperation. The conference was just one stage of the controversy that had been raging over the work of three Harvard University scientists: mathematical biologists Martin Nowak and Corina Tarnita, and E.O. Wilson. The previous month they had published a paper in the journal Nature attacking inclusive fitness.
The details of their attack were technical and mathematical, but the consequences could be far-reaching. They said inclusive fitness is irrelevant to the real world and suggested replacing it with a series of equations that could describe the evolution of cooperation in far more detail than ever before. The problem, said Nowak and Tarnita, is that the calculations that describe inclusive fitness just don’t work in the real world because they rely on a limiting set of conditions that nature does not stick to. Nowak pointed out that, in thousands of insect species, daughters leave the nest despite being as closely related to each other as the workers in an ant colony. This suggests there is some factor other than kin selection keeping workers in the nest and driving altruistic behaviour.
To some biologists this new model was a step in the right direction. To others it was lunacy, and more than 137 leading biologists signed a letter to Nature criticizing Nowak’s paper in which they stated: ‘Nowak et al. argue that inclusive fitness theory has been of little value in explaining the natural world, and that it has led to negligible progress in explaining the evolution of eusociality. However, we believe that their arguments are based upon a misunderstanding of evolutionary theory and a misrepresentation of the empirical literature.’
The arguments around group selection and inclusive fitness are still ongoing today.
Interview: Selfish genes actually explain altruistic individuals
Richard Dawkins has inspired millions with his popular science books, yet also drawn fire for controversial remarks – particularly on religion. In 2013 he spoke to New Scientist about group selection and how he felt about whether his role as the world’s most famous atheist will eclipse his scientific legacy. He is emeritus professor of evolutionary biology at the University of Oxford.
Has what drives you changed over the years?
It hasn’t changed: a love of truth, a love of clarity, a love of the poetry of science. Insofar as I show hostility to alternatives, superstition and so on, it’s because they are sapping education and depriving young people of the true glory of the scientific world view – I care especially about children in this case. It’s tragic to see children being led into dark, pokey little corners of medieval superstition.
One battle of yours has been against group selection – the idea that evolution works by selecting traits that benefit groups, not genes. You destroyed that paradigm, but then it came back again.
Something else came back under the same name. If you look carefully it turns out to be things like kin selection rebranded as group selection. That irritates me because I think it is wantonly obscuring something that was actually rather clear.
I think part of why it came back is political. Sociologists love group selection, I think because they are more influenced by emotive evaluations of human impulses. I think people want altruism to be a kind of driving force; there’s no such thing as a driving force. They want altruism to be fundamental whereas I want it to be explained. Selfish genes actually explain altruistic individuals and to me that’s crystal clear.
What subjects currently interest you in evolutionary biology?
I’m fascinated by the way molecular genetics has become a branch of information technology. I wonder with hindsight whether it had to be that way, whether natural selection couldn’t really work unless genetics was digital, high-fidelity, a kind of computer science. In other words, can we predict that, if there’s life elsewhere in the universe, it will have the same kind of high-fidelity digital genetics as we do?
When we are able to muck around with our own genes more, where do you think it will take us?
The funny thing is that if you take the two parts of the Darwinian formula, mutation and selection, we’ve been messing around with the selection part with just about every species – except our own. We have been distorting wolves to Pekineses and wild cabbages to cauliflowers, and making huge revolutions in agricultural science. And yet with a few exceptions, there have been no attempts to breed human Pekineses or human greyhounds.
Now the mutation half of the Darwinian algorithm is becoming amenable to human manipulation, people have jumped to asking questions – what’s going to happen when we start tinkering with genes? – while sort of forgetting that we could have been tinkering with selection for thousands of years and haven’t done it. Maybe whatever has inhibited us from doing it with selection will do the same with mutation.
Do you believe there is a genetic basis to irrationality?
It would be very surprising if there wasn’t a genetic basis to the psychological predispositions which make people vulnerable to religion.
One idea about irrationality that I and various other people have put forward is that the risks we faced in our natural state often came from evolved agents like leopards and snakes. So with a natural phenomenon like a storm, the prudent thing might have been to attribute it to an agent rather than to forces of physics. It’s the proverbial rustle in the long grass: it’s probably not a leopard, but if it is, you’re for it. So a bias towards seeing agency rather than boring old natural forces may have been built into us.
That may take quite a lot of overcoming. Even though we no longer need to fear leopards, we inherit the instincts of those who did. Seeing agency where there isn’t any is something that may have been programmed into our brains.
If we are irrational, perhaps one of the reasons people bristle at you is they feel their nature is under attack.
We accept that people are irrational for good Darwinian reasons. But I don’t think we should be so pessimistic as to think that therefore we’re forever condemned to be irrational.
Would you rather be remembered for explaining science or taking on religion?
To me they amount to the same thing – they are different sides of the same coin. But I suppose I’d rather be remembered for explaining science. I would be upset if people dismissed my science because of the religion.
Taking it personally
The quest for a biological explanation for altruism is intricately linked to our ideas of goodness. No wonder that biologists have taken it personally.
The nature of altruism and its similarities to the human trait of goodness make it susceptible to political, philosophical and religious subjectivity. Studying the structure of an atom isn’t personal: studying altruism can be. It certainly was for two figures in the history of altruism, Thomas Huxley and Peter Kropotkin.
Huxley, also known as ‘Darwin’s bulldog’, outlined his thoughts on this topic in an 1888 essay entitled ‘The struggle for existence’: ‘From the point of view of the moralist, the animal world is on about the same level as the gladiator’s show…Life [for prehistoric people] was a continuous free fight, and beyond the limited and temporary relations of the family, the Hobbesian war of each against all was the normal state of existence.’ For Huxley, altruism was rare, but when it occurred, it should be between blood relatives.
Kropotkin, once a page to the tsar of Russia and later a naturalist who spent five years studying natural history in Siberia, thought otherwise. In Siberia he thought that he saw altruism divorced from kinship in every species he came across. ‘Don’t compete!’ Kropotkin wrote in his influential book Mutual Aid: A factor of Evolution (1902). ‘That is the watchword which comes to us from the bush, the forest, the river, the ocean. Therefore combine – practice mutual aid!’
Radically different conclusions
How could two respected scientists come to such radically different conclusions? In addition to being a naturalist, Kropotkin was also the world’s most famous anarchist. He believed that if animals could partake in altruism in the absence of government, then civilized society needed no government either, and could live in peace, behaving altruistically. Kropotkin was following what he saw as ‘the course traced by the modern philosophy of evolution…society as an aggregation of organisms trying to find out the best ways of combining the wants of the individuals with those of co-operation’. He saw anarchism as the next phase of evolution.
Huxley was no less affected by events around him. Shortly before he published ‘The struggle for existence’, his daughter, Mady, died of complications related to a mental illness. In his despair over Mady’s passing he wrote, ‘You see a meadow rich in flower…and your memory rests upon it as an image of peaceful beauty. It is a delusion…not a bird twitters but is either slayer or slain…murder and sudden death are the order of the day.’ It was in the light of nature as the embodiment of struggle and destruction – the antithesis of altruism – that Huxley saw the death of his daughter and it was in that mindset that he penned his essay.
A suite of other fascinating characters would follow Huxley and Kropotkin. In the US there was the Quaker ecologist Warder Clyde Allee, who did the first real experiments on altruism in the 1930s and whose religious and scientific writings on the subject were often indistinguishable; in fact, he would often swipe text from one and add it to the other. Around the same time in the UK, J.B.S. Haldane, one of the founders of population genetics, was talking of altruism and kinship, and came close to developing a mathematical theory on the subject. But he stopped short – nobody quite knows why.
The mathematics of altruism
A mathematical theory for the evolution of altruism and its relation to blood kinship would come a generation later with Bill Hamilton, who was both a passionate naturalist and a gifted mathematician. While working on his PhD in the early 1960s, he built a complex mathematical model to describe blood kinship and the evolution of altruism. Fortunately, the model boiled down to a simple equation, now known as Hamilton’s rule. The equation has only three variables: the cost of altruism to the altruist (c), the benefit that a recipient of altruism receives (b) and their genetic relatedness (r). Hamilton’s rule states that natural selection favours altruism when r × b > c.
Hamilton’s equation amounts to this: if a gene for altruism is to evolve, then the cost of altruism must be balanced by compensating benefits. In his model, the benefits can be accrued by blood relatives of the altruist because there’s a chance (the probability r) that such relatives may also carry that gene for altruism. In other words, a gene for altruism can spread if it helps copies of itself residing in blood kin.
A generation of biologists were profoundly affected by Hamilton’s rule. One them was the population geneticist George Price, an eclectic genius who became depressed when he came across Hamilton’s work. He had hoped that goodness was exempt from scientific analysis, but Hamilton’s theory seemed to demonstrate otherwise. Price went through the mathematics in the model and realized that Hamilton had underestimated the power of his own theory.
While working with Hamilton on kinship and altruism, the atheist Price underwent a religious epiphany. In an irony that turns the debate about religion and evolution on its head, Price believed that his findings on altruism were the result of divine inspiration. He became a devout Christian, donating most of his money to helping the poor. At various times he lived as a squatter; at other times he slept on the floor at the Galton Laboratory of University College London, where he was working. Price lived the life of the altruists that he had modelled mathematically.
Does ‘survival of the fittest’ justify ‘everyone for themselves’?
The phrase ‘survival of the fittest’ is widely misunderstood. Although the phrase conjures up an image of a violent struggle for survival, in reality the word ‘fittest’ seldom means the strongest or the most aggressive. On the contrary, it can mean anything from the best camouflaged or the most fecund to the cleverest or the most cooperative.
What we see in the wild is not every animal for itself. Cooperation is an incredibly successful survival strategy. Indeed it has been the basis of all the most dramatic steps in the history of life. Complex cells evolved from cooperating simple cells. Multicellular organisms are made up of cooperating complex cells. Superorganisms such as bee or ant colonies consist of cooperating individuals.
Besides, it is nonsense to appeal to the ‘survival of the fittest’ to justify any economic or political ideology, especially on the basis that it is ‘natural’.
Is cannibalism fine because polar bears do it? Is killing your brother or sister fine because nestlings of many bird species do it? Just about every kind of behaviour that most of us regard as ‘unnatural’ turns out to be perfectly natural in some nook or cranny of the animal kingdom. No one can plausibly argue that this justifies humans behaving in the same way.
Yet even though such examples expose the utter absurdity of appealing to what is ‘natural’ to judge right from wrong – the naturalistic fallacy – we seem to have a strange blind spot when it comes to evolution. Survival of the fittest has been claimed to justify all kinds of things, from free markets to eugenics. Such notions still have a powerful grip in some circles.
However, natural selection is simply a description of what happens in the living world. It does not tell us how we should behave.