The Structure of Flowers |
There are many ways in which the various flower parts can be arranged to make up the structure we recognise as a ‘flower’. This chapter sets out to describe the structure of a simple flower, and then discusses some of the variation commonly encountered. Chapter 8 includes some more complex examples.
Floral structure is described under the following headings:
Structure of a basic flower
The perianth
The reproductive organs
Variation in floral structure
Arrangement and union of parts
The perianth
The calyx
The corolla
Symmetry
Aestivation
The reproductive organs
The androecium
The gynoecium
Placentation
The style and stigma
Discs and nectaries
Relationships of parts in the flower
The floral tube
Number of parts per whorl
Unisexual flowers
The floral formula
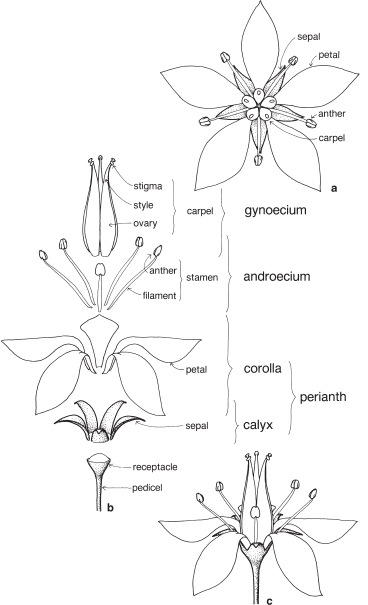
A basic flower (Fig. 1; Pl. 1) has four series of parts arranged in concentric whorls (or rings) on the receptacle, which is the name given to the expanded end of the pedicel (flower stalk). The two outer whorls are together known as the perianth, and are not directly involved in reproduction. The reproductive structures are located in the inner whorls.
The perianth
The outer whorl, known as the calyx, is composed of two or more parts called sepals, which are often green in colour and enclose the rest of the flower in the bud stage. Inside the calyx is the corolla, made up of petals, which are usually white or brightly coloured. It is usual for the sepals and petals to be equal in number.
The reproductive organs
A whorl of stamens, called the androecium, lies inside the corolla, and is generally taken to represent the male part of the flower. Each stamen has a slender filament (stalk) and, at the top, an anther in which the pollen is produced. The pollen grains carry the male reproductive units.
In the centre of the flower is the gynoecium, made up of carpels. Each carpel usually has three parts: an expanded basal part called the ovary, in which the ovules are produced; a central stalk-like section called the style; and a terminal stigma. The ovules contain the female egg-cells and the stigma surface is specially adapted to receive compatible pollen. The ovules, after fertilisation and further development, become seeds and the mature carpels with the enclosed seeds develop into the fruit. The gynoecium is generally regarded as the female part of the flower.
Figure 1b illustrates an expanded flower with the various whorls separated from each other. This places the receptacle at the bottom of the diagram, with the other parts laid out in sequence moving from the outside to the centre of the flower. Figure 1a shows the flower from above and illustrates the way in which the parts in adjacent whorls usually alternate with one another. The same flower is illustrated from the side in Figure 1c.
Variation in floral structure
Much of the variation in floral structure is embodied in four aspects:
• the number of whorls present (sometimes one or more whorls are absent)
Fig. 1 Structure of a basic flower: a from above; b expanded flower, showing the series of parts; c side view.
• the number of parts in any whorl
• the degree of union of parts, either within a particular whorl or between adjacent whorls
• the position of the ovary with respect to the other parts.
Arrangement and union of parts
Although the majority of species have their flower parts arranged in whorls, others have numerous parts arranged spirally on a more or less elongated receptacle, for example, in the genera Magnolia (Fig. 22) and Liriodendron (Pl. 6a–d).
Parts within a whorl may be united by their adjacent margins into a more or less single structure. The petals for example may be united into a tube-like form (Fig. 2j, k). The degree of union may only be small, or incomplete, for parts still to be considered united; for parts to be taken as free they should be completely so (and each then generally falls individually from the receptacle as the flower ages). Some texts use the term distinct instead of free.
The perianth
The calyx
The sepals may be separate from one another as in Figure 1 or wholly or partly united (Fig. 99; Pls. 14b, 21b). Sometimes an extra ring of sepal-like segments is attached outside the true calyx. This is known as an epicalyx and is typical of the genus Hibiscus (Pl. 21h) and of some other members of the family Malvaceae (Fig. 99; Pl. 21). Sometimes the sepals fall as the flower opens, as, for example, in many species of Papaver (Poppy, Pl. 4); in Pittosporum undulatum (Sweet Pittosporum, Fig. 131) the sepals may be shed before the petals drop. In other species the calyx persists when the flower fades, and becomes a conspicuous feature of the fruit, for example, Physalis spp. (Groundcherry, Cape Gooseberry, Pl. 30i).
The corolla
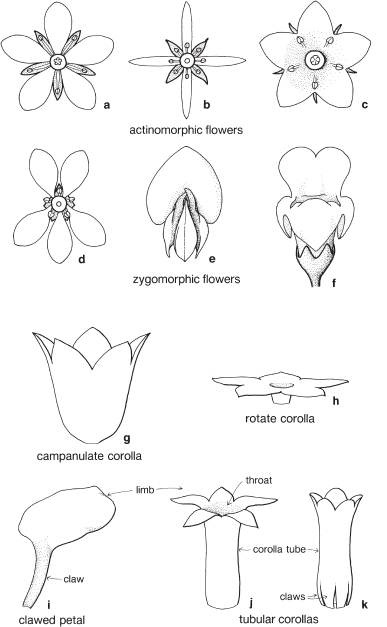
The petals may also be separate from one another or united. When united, the extent of the fusion and the shape and disposition of any free parts give rise to a variety of corolla types, some of which are illustrated in Figure 2, and Plates 28–31. The corolla tube is the term used to describe the usually tubular basal section, the throat is the top of the tube and the limb is the expanded part of the corolla above the throat and the tube (Fig. 2j). The term limb is also applied to the expanded upper section of a free petal that has a narrow basal part called a claw (Fig. 2i; Pl. 25d). Union may take the form of a very short tube, as in Sprengelia incarnata (Swamp Heath, Fig. 110), or a tube in which the petals are free at the base, as in some species of Correa and Stackhousia (Figs. 2k, 75). The corolla tube is then said to be clawed. In the family Goodeniaceae the petals are united but the corolla is usually split open down one side (Fig. 115a; Pl. 5e).
Fig. 2 The corolla: some terms describing shape and symmetry.
Appendages on the petals occur in some groups, for example, the corona or trumpet of the genus Narcissus (Daffodil and Jonquil), the coronal scales of Silene coronaria (Rose Campion, Pl. 25d), and the spur, a tubular projection from a petal seen in Velleia paradoxa (Spur Velleia).
The colour of the corolla may not be important in the identification of plants; for example, in Epacris impressa (Common Heath, Pl. 28h) it may be white, pink or red.
Sometimes the perianth segments, although in two whorls, are alike in size, colour and texture and then it is usual to retain the collective term perianth (or in this context the particular term perigon) and refer to the units as perianth parts or tepals (Figs. 23, 32, 35). Such flowers are typical of plants commonly thought of as lilies, belonging to a number of families (Pls. 8h–l, 11a–f).
In some groups of plants there is only one whorl of perianth parts. The term perianth may be used to refer to the single whorl even if it is known to be a calyx or corolla. A single perianth whorl occurs in Grevillea (Figs. 49, 50; Pl. 3a, b), Clematis (Fig. 42), and the widespread Foeniculum vulgare (Fennel, Fig. 132). Sometimes all the perianth parts are absent or are reduced in size, often becoming brown and scale-like. In Allocasuarina (She-oak, Pl. 17d–g) the perianth is absent in the female flowers and reduced to fine brown scales in the male flowers.
Symmetry
When a flower is observed from above, the sepals and petals may be arranged on the receptacle in a symmetrical manner and the flower can be divided into equal parts by cutting through its centre in more than one plane (Figs. 2a–c; Pls. 1b, 2f, 3d). Such a flower is said to be actinomorphic or regular. Alternatively, a flower may be asymmetrical and can be divided into equal parts by cutting in one plane only. This flower is said to be zygomorphic or irregular (Figs. 2d–f; Pls. 3b, 5e, g). Some zygomorphic corollas are clearly bilabiate or two-lipped, with one or two of the petals grouped on one side of the flower and the rest on the other side (Fig. 2f). Many members of the families Scrophulariaceae (Figworts, Pl. 31) and Lamiaceae (Mints) have bilabiate corollas and, in the latter family, the calyx may also be bilabiate.
Aestivation
This is the term applied to the arrangement of the floral parts, usually the petals, in the bud. If the petals come together edge to edge they are said to be valvate, but when they overlap they are described as imbricate. Various other terms define more specialised arrangements, but these are not commonly encountered in identification texts.
The reproductive organs
The androecium
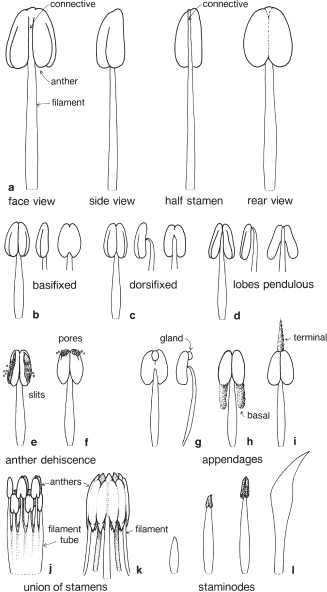
Immediately inside the corolla are one or more stamens collectively called the androecium (Fig. 1), a word derived from the Greek and meaning ‘male house’. When numerous, the stamens may be in more than one whorl. Each stamen has a filament or stalk supporting an anther. The anther usually has two lobes, often lying parallel to one another, which are united by a tissue called the connective, in many cases effectively an extension of the filament (Fig. 3a). Each anther lobe contains two sacs in which the pollen is produced.
The filament may be attached at the base of the anther lobes, when it is said to be basifixed (Fig. 3b; Pls. 1d, 15h), or at the back, dorsifixed (Fig. 3c; Pl. 4i). When the point of attachment is small, the anther may move freely with respect to the filament and is said to be versatile. If the filament is joined to the apex of the anther the lobes are described as pendulous (Fig. 3d), and if no longer parallel to one another are said to be divergent. Some stamens have extra appendages, for example, a projection of the connective above the anther lobes (Figs. 3i, 94) or an extension of sterile tissue at the base of the lobes (Fig. 3h; Pl. 28d).
When the pollen is mature, the wall of each anther lobe usually develops a longitudinal slit (Fig. 3e; Pl. 1e) or a terminal pore (Fig. 3f; Pl. 4l), through which the pollen is shed. The splitting of the anther, and the consequent shedding of pollen, is known as dehiscence. Another, less common, means of releasing pollen is seen in Lauraceae (Laurel Family) in which the anthers are provided with dehiscence flaps (Pls. 6h, 7e, h, l). Each mature pollen grain contains a generative cell that, when conditions are favourable, eventually produces two male sperm cells.
Stamens are usually separate from one another, but may be united by their filaments or anthers. For example, in Hibiscus (Pl. 21h) and in Ulex europaeus (Gorse or Furze) the filaments are united for most of their length to form a tube, but the anthers are free (Figs. 3j, 69, 99). In the genera Melaleuca and Lophostemon the stamens are grouped in bundles due to the fusion of the filament bases (Fig. 91; Pls. 19j, 20c) and in members of the Daisy Family, Asteraceae, the anthers are united to form a tube around the style but the filaments remain free (Figs. 3k, 117d, 123c, d; Pl. 32e, i).
Fig. 3 The androecium—stamens: a stamen structure; b–d anther atachment; e–f anther dehiscence (see also Fig. 77); g–i appendages (see also Figs. 36, 94, 95; Pl. 28d): j union of filaments (see also Fig. 91, Pl. 21h, i); k union of anthers (see also Fig. 117); l staminodes (see also Pls. 6h, 7i, 31d).
Stamens are not always borne on the receptacle but may be attached to the petals or tepals. Stamens are said to be epipetalous (epi = upon) if attached to the petals as in Epacris impressa (Common Heath, Fig. 104) and Brugmansia (Pl. 29h), and epitepalous if joined to the tepals as in Agapanthus praecox (Fig. 35b) and Amaryllis belladonna (Pl. 11b). Sometimes the stamens are reduced to anthers only. In Grevillea (Fig. 50c; Pl. 3a), the anthers join directly to the tepals, with no discernible filaments.
Occasionally the anther is reduced to one lobe, for example, in some stamens in the flowers of Conospermum (Smoke Bush, Fig. 47d). In some plants one or more stamens may be sterile due to a failure to produce pollen, or the anthers may be deformed or absent. In the latter case the filament may be reduced in length or modified to become petal-like (Fig. 3l). Abortive stamens are called staminodes (Pls. 6h, 7i, 31d).
In a flower with the same number of sepals, petals and stamens, the stamens nearly always alternate with the petals and so are opposite the sepals (Fig. 1a; Pl. 1b). An exception is found in some members of the family Rhamnaceae in which the petals are opposite and hooded over the small stamens, for example, Spyridium parvifolium (Dusty Miller), or Cryptandra amara (Bitter Cryptandra, Fig. 73).
The gynoecium
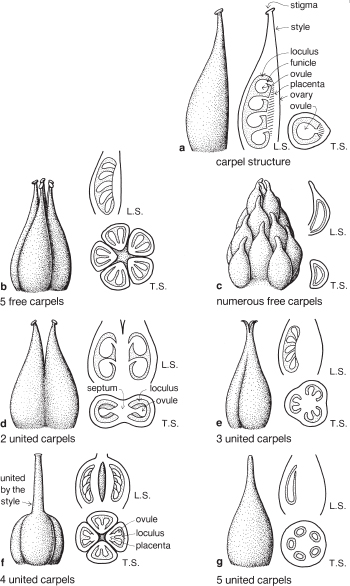
The innermost whorl of flower parts is called the gynoecium (Fig. 1), a term also derived from Greek and meaning ‘female house’. The gynoecium consists of one or more structures called carpels. An external view of a single carpel is shown in Figure 4a and Plate 3e. The swollen section at the base is the ovary, the stalk at the top of the ovary is the style, and near the end of the style is the stigma, a special tissue to which pollen can adhere and on which compatible pollen will germinate. (See the section on pollination in Chapter 4.)
Figure 4a also shows one carpel that has been cut vertically from top to bottom, seen as well in Plate 1f. This view is known as a longitudinal section or L.S. The cavity inside the ovary, called the loculus (or locule, plural = loculi or locules) contains the ovules, which become the seeds after fertilisation and subsequent development. Each ovule is attached by a funicle (a stalk, sometimes very short) to the placenta, a tissue that lines part of the inside of the ovary wall. (See also Plates 3f, 5c, 10d, 20f, g.)
A gynoecium of only one carpel is found in a number of large families including the Leguminosae (Legume Family) and Proteaceae, the family that includes Grevillea and Banksia (e.g. Pls. 3, 14, 15).
Most flowers have two or more carpels, which are either free or wholly or partly united. A gynoecium of free carpels is said to be apocarpous (Figs. 4b, c; Pls. 1, 2), and is typical of the families Magnoliaceae (Fig. 22), Ranunculaceae (Buttercups, see Fig. 42c; Pl. 2a–c), and Crassulaceae (Stonecrops) which includes many succulent rockery plants, such as Crassula (Pl. 1) and Sedum (Fig. 55).
A gynoecium of two or more carpels fused together into a single structure is said to be syncarpous (syn- = with, i.e. united). The degree of union can vary, and some of this variation is illustrated in Figure 4d–g
and Plates 4, 9c, d, 25d.
A gynoecium of two fused carpels is shown in Figure 4d. The ovary here is completely divided into two loculi (cavities) and the wall between them is called the septum (plural = septa). In this case the septum bears the placentas. The styles and stigmas are free in this diagram indicating incomplete union, but often it is complete and there is then only one style and one stigma (Pls. 4l, 20f, i, 22g).
Sometimes a syncarpous gynoecium will contain only one loculus. In such cases the number of carpels involved will usually be indicated either by a divided style and/or stigma (Fig. 5e), or by the presence of more than one placenta within the ovary (Fig. 5g). An example of this type is seen in Passiflora (Passion Flower), which has a gynoecium composed of three united carpels with three styles and stigmas and a unilocular ovary. When a passionfruit is cut open and the pulp removed, the stalks to which the seeds were attached are seen in three groups on the inside of the fruit wall, indicating the location of the three placentas.
Partial fusion of the carpels also occurs in some genera of the Rutaceae, for example Boronia (Fig. 92; Pl. 4i), and in these flowers the carpels are free at the base but united above so there is only one style and stigma (Fig. 4f).
In practice the term gynoecium is rarely used in literature directly concerned with plant identification. Features of the female part of the flower are covered by the terms ovary, style and stigma, as well as carpel. In botanical descriptions it is usual to read statements such as ‘the ovary is trilocular’, or ‘the ovary is 1–4 locular’. The latter statement also illustrates the extent of the variation in carpel number in some groups.
Fig, 4 The gynoecium—some possible combinations of free or united carpels: a structure of a single carpel; b–c free carpels; d–g united carpels.
How many carpels are in this flower?
Guidelines for finding the number of carpels when they are united
Cut a transverse section (T.S.) of the ovary and observe the number of loculi:
If the number of loculi > 1, then no. of carpels = no. of loculi
If only 1 loculus is present, then no. of carpels = no. of 
See e.g. Figs. 4d–g and 5d–h, and Pls. 4d, k, 11c and 20d, g. These guidelines will work in the majority of cases, but are not infallible. It is suggested that sections be cut at several different levels to cover the rare occasions where the septa are incomplete, and loculi number may differ, for example at the base or towards the top of an ovary. See also the accounts of the families Lamiaceae (no. 41) and Goodeniaceae (no. 43).
In some texts, often in American literature, the term ‘pistil’ may be used to refer to the female part of the flower and, depending on the context, it can have several shades of meaning. A flower with a single carpel could be described as possessing a ‘simple pistil’, or a flower with free carpels could be said to have ‘several free pistils’. When two or more carpels are united the flower could be said to have a ‘compound pistil’.
Placentation
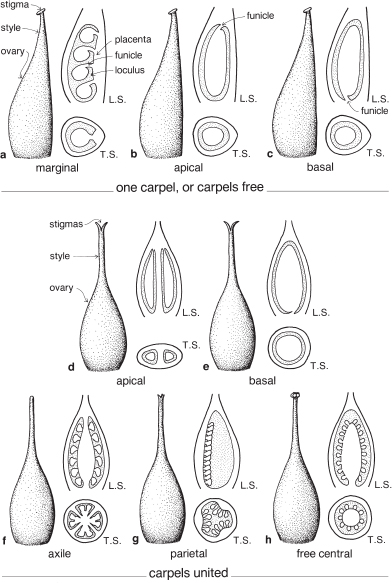
The placenta is the tissue in the ovary to which the ovules are attached. The arrangement of the ovules (and hence the position of the placenta) within the ovary is termed the placentation and its determination is sometimes required when identifying a plant.
In a single carpel with numerous ovules the placentation is marginal (also sometimes called lateral, Fig. 5a; Pl. 1f). If there is only one ovule (or few) it may be attached at the top of the loculus, apical placentation (Fig. 5b) and then the ovule is said to be pendulous, or at the base of the loculus, basal placentation (Fig. 5c; Pl. 2c).
If the gynoecium is syncarpous (made up of two or more united carpels) and there are two or more loculi within the ovary, the placentas are often situated on the central axis and the placentation is said to be axile (Figs. 4d–f, 5f; Pls. 11c, 28e, 29i). However, if there is only one ovule (or few) in each loculus the placentation may be apical or basal (Figs. 5d, e; Pl. 5c).
Fig. 5 Placentation—attachment of ovules in the ovary. Placentation types are not restricted to the examples shown here, for example, axile placentation (f) may occur with two or more united carpels.
Other less common types of placentation occur when the ovary is unilocular and a number of united carpels are involved. For example, if within the loculus the placenta is raised on a central column the placentation is described as free central (Fig. 5h; Pl. 25i), and is typical of the family Primulaceae, for example Anagallis (Pimpernel) and Primula (Primrose). Again, a unilocular ovary may contain two or more placentas attached to its inside wall. This type of placentation is called parietal (Fig. 5g; Pl. 4k) and is found, for example, in Passiflora (Passion Flower). Another type of parietal placentation is seen in Papaver (Poppy, Pl. 4d) in which plates of tissue carrying the placentas project inwards towards the centre of the loculus.
The style and stigma
The style is the tissue that links the top of the ovary with the stigma. It may be elongated (Figs. 5d–h; Pls. 9c, 31m), or relatively short (Figs. 5a–c; Pl. 20f), or occasionally absent (Pl. 4c). It is commonly inserted at the apex of the ovary (Fig. 6a), less frequently in a depression in the top of the ovary (Figs. 6b, 104b), or joined to the base of a deeply lobed ovary (Figs. 6c, 113c) when it is described as gynobasic.
If the gynoecium is syncarpous, the stigma may be lobed (Figs. 5g–h, 80) or the style may branch either close to the point of insertion on the ovary (Pl. 25d) or nearer to the end (Fig. 5d–e), and such branches are referred to as style-arms or style-branches. In the majority of cases the number of lobes or style-arms will equal the number of united carpels.
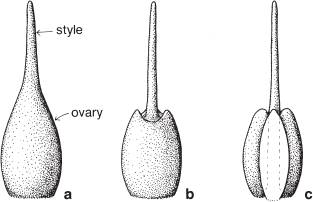
Fig. 6 Insertion of the style on the ovary: a terminal; b inserted in a depression; c inserted at the base of a deeply lobed ovary (gynobasic).
The stigma is usually located on or near the end of the style or stylearms, and often appears as a discernible individual ‘structure’ (Pls. 3e, 31d), although sometimes the boundary between the style and its stigmatic surface is obscure (Pls. 1f, 5b). The stigmatic surface may be covered with short glandular hairs or papillae (small rounded projections), which provide a receptive tissue on which compatible pollen may germinate. (See section on pollination, Chapter 4.) If the stigmatic region occupies most of the style-arm, for the purposes of identification, the terms style-arm and stigma may effectively be interchangeable.
While the end of the style of a single carpel is usually undivided, it may be expanded in various ways; for example, in Hakea and Grevillea (family Proteaceae) the style-end is either conical or discoid (Figs. 49, 50; Pl. 3a, b).
Discs and nectaries
Discs are not found in all species, but when present they are usually associated with the ovary, although not strictly part of it. A disc located at the base of an ovary is referred to as hypogynous (hypo = below, gynous = female), while one across the top is an epigynous disc (epi = upon). The hypogynous disc may be a continuous ring of tissue (Fig. 7a) or in the form of flat or rounded lobes (Fig. 7c), or it may appear as a swelling on one side of the ovary only (Fig. 7b). Discs are often green, but may be coloured (Pl. 30c) and, as they often produce nectar, may also be referred to as nectaries or nectary glands. Individual lobes may be referred to as nectariferous scales (Pl. 1c). The illustrations of Boronia (Fig. 92; Pl. 4i), Zieria (Fig. 98), Foeniculum (Fig. 132), Petroselinum (Pl. 5b) and Scrophularia (Pl. 31d) all include discs.
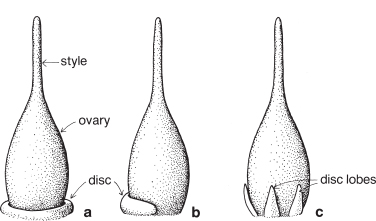
Fig. 7 Some common forms of disc occurring at the base of the ovary. The disc often produces nectar; it may also be referred to as a nectary (see also Figs. 92, 98, 132c; Pls. 1c, 3a, 4i, 30c, 31d).
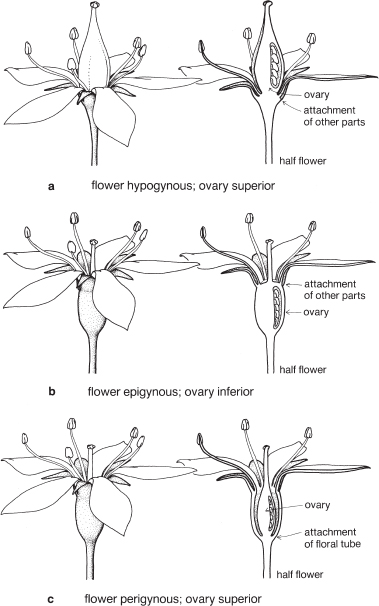
Fig. 8 Flower types and ovary position: three commonly recognised types of flower and the corresponding ovary position with respect to the attachment of the perianth and stamens or the floral tube.
Nectar is commonly seen in flowers and, depending on the species, may be produced by specialised tissue asscociated with any of the floral parts. For example, the tepals of many lily-like species are nectar-producing (usually from near the base), as are specialised zones on the petals in Ranunculus (Buttercup). The elongated spurs produced from sepals or petals in various species may secrete nectar, or function more simply as containers for nectar produced elsewhere in the flower. The floral tubes seen in Rosaceae (e.g. Prunus, Pl. 3e), Onagraceae (e.g. Fuchsia, Fig. 80), and Myrtaceae (e.g. Leptospermum, Pl. 20i) are often lined to some degree with nectar-producing tissue. Nectariferous glands may also be associated with stamens as in members of the Lauraceae (Laurel Family, Pls. 6h, 7).
With respect to the carpels, tissue in various locations may produce nectar, depending on the species. In some cases it may be secreted by the whole outer surface. When carpels are united, nectar may be produced by tissue within the ovary, for example associated with the septa between the loculi. Septal nectaries are recorded for numerous genera with lilylike flowers such as Agave spp., Amaryllis belladonna (Belladonna Lily, Pl. 11c) and Narcissus (Daffodil and Jonquil). In these particular cases the nectaries are visible in a T.S. of the ovary, forming part of each septum but a (slightly) different colour and readily oozing liquid.
Relationships of parts in the flower
In most flowers, the parts of adjacent whorls alternate; thus the sepals alternate with the petals, petals with stamens, and stamens with carpels (Fig. 1; Pl. 1). Another way of describing this arrangement is to say that the stamens are opposite the sepals and the carpels are opposite the petals. Recalling this typical pattern can assist with the interpretation of flowers with an unusual structure, for example, when one whorl of parts is absent.
The relative position of floral parts is one of the most important clues to the interpretation of floral structure.
Fig. 9 The floral tube.
When the flower parts are arranged on the receptacle so that the sepals occupy the lowest position, followed in order by the petals and stamens, with the carpels (whether free or united) at the top, the flower is said to be hypogynous, and the ovary is superior (Figs. 1, 8a; Pls. 1, 4).
Sometimes in a flower with a superior ovary, the receptacle appears to be cup-shaped so that the sepals, petals and stamens originate around the ovary. Such a flower is said to be perigynous (peri = around, Fig. 8c). The cup-like structure may be called the floral tube. Some genera of the family Rosaceae have perigynous flowers, including Prunus (Cherry, Fig. 72. Pl. 3e), Rosa (Rose) and Spiraea (Pl. 2g).
If the sepals, petals and stamens appear to come from the top of the ovary, the flower is said to be epigynous and the ovary is inferior (Figs. 8b, 132; Pl. 5a–c). Occasionally, flowers appear to have an inferior ovary but the top of the ovary protrudes above the point where sepals, petals and stamens arise. Such an ovary is considered to be semi-inferior, and is seen, for example, in some species of Pomaderris (Fig. 74c) and Leptospermum (Pl. 20i).
The ovary is superior (above) or inferior (below) with respect to the point of attachment of the other floral parts (perianth and stamens).
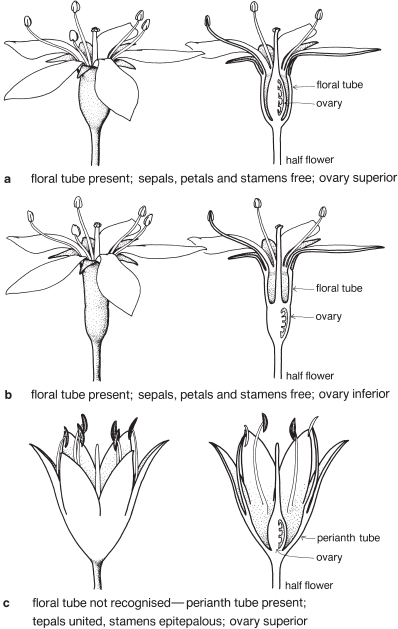
The floral tube
The floral tube may be defined as consisting of the fused bases of the perianth parts (both calyx and corolla if both are present) and the stamens (Figs. 9a–b). (Note that one of the perianth whorls may be absent, as in Pimelea—see Fig. 101.) It should be clearly distinguished from a corolla tube which involves the union of petals only, as shown in Epacris (Fig. 104) and Brugmansia (Pl. 29g, h).
The floral tube has been the subject of much debate, particularly with respect to its derivation, and has been given diferent names by various authors. The neutral term ‘floral tube’ is not widely used in the literature. The most common alternative is hypanthium, and others used include thalamus tube, receptacle tube and torus. Sometimes the term calyx tube is used in this sense but is not strictly correct. Note that the term ‘torus’ is also synonymous with ‘receptacle’, implying the interpretation that the floral tube is an outgrowth from, or part of, the receptacle. A flower may thus be described as having a ‘hollow cup-shaped torus’. This interpretation was used extensively in the older text Flora of South Australia by J.M. Black.
Sometimes the ovary is described as being ‘adnate to the floral tube’. This implies an inferior, or at least semi-inferior, ovary. In this sense the floral tube may not be evident as a separate structure, as the tissues of the ovary wall and floral tube are integrally fused. In the example of Fuchsia (Fig. 80; Pl. 19d), the floral tube may be described as ‘adnate to, and produced above the ovary’.
The presence of a floral tube should not be a problem to those identifying plants except that it may affect the decision about freedom or union of perianth parts. When a floral tube is present and the sepals and petals are distinctly different, and arise independently from the top of the tube, they are usually regarded as being free (Figs. 9a, 90; Pl. 20h, i).
When a floral tube is technically present but the perianth is not obviously differentiated into sepals and petals, the floral tube may be ignored and the tepals described as united. Then, the stamens can be regarded as joined to a perianth tube, and the term epitepalous stamens applies (Figs. 9c, 35; Pl. 11b). In these cases the tube usually appears to be a continuation of the perianth.
Floral tubes may be encountered in members of a number of families, such as:
| Celastraceae | Stackhousia (Fig. 75) |
| Myrtaceae | Eucalypts, Tea-trees, etc. (e.g. Fig. 90; Pl. 20f, i) |
| Onagraceae | Fuchsia (Fig. 80; Pl. 19d), Oenothera (Pl. 19b) |
| Rhamnaceae | Cryptandra (Fig. 73) |
| Rosaceae | Acaena (Fig. 70), Chaenomeles (Fig. 71), |
| Prunus (Fig. 72; Pl. 3d, e), Spiraea (Pl. 2g) | |
| Thymelaeaceae | Pimelea (Fig. 101). |
When a floral tube is present, that is, the bases of all perianth parts and stamens are united—decide about the freedom or union of parts:
• at the top of the tube if the sepals and petals are obviously different
• at the bottom of the tube if the sepals and petals are similar in size, colour and texture.
The number of parts in each whorl of the flower varies from species to species. The degree of variation is considerable, ranging from zero to numerous (the latter term is usually applied when the number of parts exceeds about 10). Parts commonly occur in multiples of the petal number, for example 5 sepals, 5 petals, 5 or 10 stamens and 5 carpels. Floral parts in fours or fives occur in the majority of Flowering Plants, but floral parts in threes are also associated with a significant number of families—such as all the Lilies and Irises, as well as the Magnolia and Laurel groups.
Reduction in the number of reproductive parts is common; for example, often there are only two carpels in a 4- or 5-partite flower, or fewer stamens than petals.
Unisexual flowers
Most flowers have both stamens and carpels and are therefore bisexual. However, some have no stamens and thus are unisexual and functionally female. Similarly, flowers without carpels are functionally male. Unisexual flowers may occur on the same plant, as in cucumbers, marrows and pumpkins and sometimes in Allocasuarina (She-oak). Such species are said to be monoecious (from Greek for ‘one house’). When the male and female flowers are borne on separate individuals the species is said to be dioecious; examples include Clematis (Fig. 42), Wurmbea dioica (Early Nancy, Fig. 23), and most species of Allocasuarina (Pl. 17d–g).
Further information on floral structure may be found in Bowes (1996), Judd et al. (2016), Lawrence (1951) and Weberling (1989).
A floral formula is a shorthand means of recording the basic structure of a flower. Standard symbols used for the floral organs are listed below (although some texts may vary these slightly). Formulae have been used here in most captions; they have been expressed as simply as possible to indicate what is readily visible in the flower.
| P | perianth—tepals |
| K | calyx —sepals |
| C | corolla —petals |
| A | androecium—stamens |
| G | gynoecium —carpels |
| G ovary superior | |
| –G– ovary semi-inferior | |
| G ovary inferior | |
| ∞ | parts numerous (>10) |
| ( ) | parts of a whorl united |
 |
used in this book to indicate parts of a whorl at first united and becoming free as the flower ages, or parts particularly tenuously united |
 |
links united parts of different whorls |
| + | indicates parts are in more than 1 whorl |
| ‚ | separates numbers of parts within a whorl |
| – | linking two numbers, indicates a possible range of parts within a whorl |
Examples
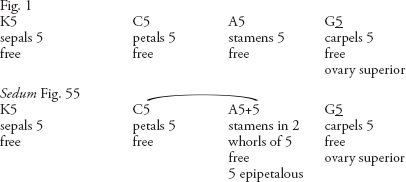
In a few cases, the form ‘G(loculi)’ has been used to indicate the number of loculi visible in a transverse section of the ovary, instead of noting the number of carpels in the gynoecium (e.g. Figs. 112, 113, 115).