Inflorescences—the arrangement of flowers on the plant |
An inflorescence can be thought of as a shoot system that is modified for the production of flowers. Familiarity with the main types of inflorescences can significantly aid species recognition. Some are particularly distinctive and characteristic of whole families, such as the umbel in the Apiaceae (Carrot and Parsley Family, sometimes called Umbelliferae, e.g. Pl. 34b, c), and the head in the Asteraceae (Daisy Family, also called Compositae, e.g. Pl. 32c, h). The older names of these two families reflect their general inflorescence types.
Solitary flowers
Flowers are said to be solitary when the plant bears only one, or when single flowers appear on the ends of lateral branches that are remote from one another (Pls. 4a, 8j). In practice, the inflorescence of a shrub with one flower in each leaf axil on many shoots is often described as ‘flowers solitary in the axils of the leaves’.
Multi-flowered inflorescences
Inflorescences of more than one flower can be relatively simple or develop into quite complex flowering-branch systems. They are usually quite distinct from the vegetative parts of the plant.
Differences in their overall appearance from one species to another are mostly determined by variations in a small number of features including the pattern of branching, the relative elongation of branches and flower stalks, the sequence of flower development (i.e. the position in the inflorescence of the older and younger flowers), and the repetition of basic flowering ‘units’ within a more complex inflorescence.
One important aspect of inflorescence structure concerns the nature of the main axis. A determinate inflorescence, sometimes called a definite inflorescence, is one in which the main axis ends in a flower. In contrast, the inflorescence is indeterminate (indefinite) if the main axis continues growth without a definite conclusion, giving rise to a smaller or larger number of lateral flowers. The later flowers may remain incompletely developed, and atrophy along with the end of the axis.
There are two main forms of branching, racemose and cymose, and some inflorescences are a combination of both.
Racemose branching
A racemose pattern of branching has traditionally been defined on the basis of an indeterminate main axis that continues apical growth, producing lateral buds which become flowers or flowering shoots (Fig. 10). This pattern of branching may also be called monopodial. The youngest buds or flowers are nearest the apex. Members of the widespread family Brassicaceae (Pls. 23, 24a–e), which includes numerous cultivated genera such as Brassica (Cabbage), Eruca (Rocket) and Raphanus (Radish) produce inflorescences of this nature.
Cymose branching
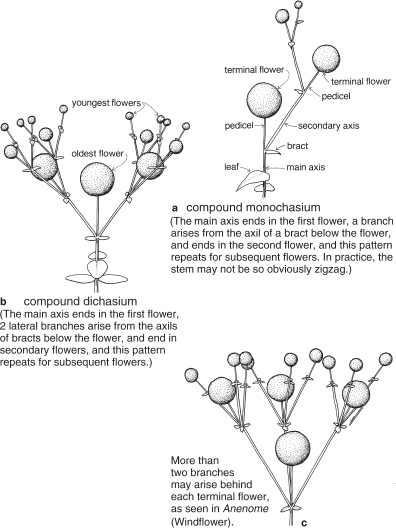
In contrast, inflorescences with cymose branching have growth of the main axis terminated by a flower. One or more lateral buds behind that flower then develop until their growth in turn concludes with the production of a flower. This branching pattern is also considered sympodial. While there are a number of different types of such inflorescences recognised (some illustrated in Figure 11), flowers may simply be described as being in ‘cymes’ indicating an inflorescence of this general type but avoiding the need to be specific. Inflorescences with cymose branching are common in the family Caryophyllaceae (Pl. 25) which includes many familiar garden subjects such as Gypsophila (Baby’s Breath) and Silene (Catchfly and Campion), and common weeds in the genera Cerastium and Stellaria (Chickweeds).
Some inflorescence types, such as the head and umbel, can occur with either racemose or cymose branching. In these types, one indication of cymose branching is the youngest flowers being towards the outside of the inflorescence, instead of towards the centre as shown in Figures 10d and 10f. When the two forms of branching are combined in one inflorescence, it often appears as a racemose main axis and cymose lateral branches in a form called a thyrse (Pl. 31b).
The names of some inflorescence types, such as spike, umbel and panicle, are often used as adjectival forms in descriptions of compound inflorescences. The term spicate is commonly used to describe a narrow spike-like form, and an inflorescence of branches radiating from the top of the stem could be referred to as umbellate. In practice, the term paniculate may be used to refer to a multi-branched inflorescence irrespective of branching pattern.
Fig. 10 Racemose inflorescences: the smallest spheres represent the youngest flowers.
Fig. 11 Some inflorescences with cymose branching.
The smallest spheres represent the youngest flowers. A simple monochasium is similar to that in a but consists of only the two lowest flowers. A simple dichasium consists of the first terminal flower and two lateral flowers only. Inflorescences with cymose branching are common in the family Caryophyllaceae (see Pl. 25).
A corymb is a flat-topped racemose inflorescence brought about by the lateral branches or flower stalks elongating to varying degrees such that the flowers are roughly in the same horizontal plane. The term corymbose may be used to describe any flat-topped inflorescence as in the compound form ‘corymbose panicle’ (Fig. 10g).
The informal term ‘cluster’ is used in a general sense and does not imply any particular branching pattern.
Figure and plate numbers of illustrations of some inflorescence types are set out in the following list.
| Flowers solitary | Caladenia Fig. 25; Chiloglottis Fig. 24; Epacris Fig. 103, Pl. 28h; Lagunaria Pl. 21a, b; Liriodendron Pl. 6a, c; Papaver Pl. 4a; Salpichroa Pl. 30a |
| Spike | Acacia floribunda Fig. 59; A. sophorae Pl. 14j; Callistemon Fig. 81; Melaleuca Pl. 19j; Stackhousia Fig. 75 |
| Raceme | Brassica Pl. 23b; Capsella Pl. 24a, b; Fuchsia Pl. 19d; Grevillea Figs. 48, 50, Pl. 3b; Lathyrus Fig. 64; Medicago Pl. 16h; Paraserianthes Pl. 14a; Persea Pl. 7j; Prunus Pl. 3c; Senna Fig. 56, Pl. 15b; Stylidium Fig. 114; Thomasia Fig. 100 |
| Panicle | Conospermum Fig. 46 |
| Umbel (or umbellate) | Agapanthus Fig. 34; Amaryllis Pl. 11a; Eucalyptus Fig. 85 |
| Compound umbel | Daucus Pl. 34b; Foeniculum Fig. 132 |
| Head | Acacia Figs. 58, 60–62; Pl. 14d, g, h; Asteraceae Figs. 116–125; Pls. 32, 33 (except 33k, l) Pimelea Fig. 101 |
| Compound head | Calocephalus Fig. 126; Pycnosorus Pl. 33k, l |
| Cyme | Cerastium Pl. 25f; Lophostemon Pl. 20c; Silene Pl. 25a |
| Cymose panicle | Crassula Pl. 1a; Lasiopetalum Pl. 22f; Zieria Fig. 97 |
A number of other terms are commonly found in descriptions of inflorescences. The stalk bearing an inflorescence is usually referred to as the peduncle. The stalk of an individual flower is called a pedicel. When a peduncle or pedicel is absent the structure is referred to as sessile, which is a general term for ‘stalkless’. Sometimes a stalk is very short, and the term subsessile is employed, removing the need to be specific about its presence or absence. The central axis of an inflorescence (effectively continuing the peduncle beyond the first inflorescence branch or flower) is termed the rachis.
In many cases inflorescence branches and flowers arise in the axil of a bract (Fig. 10a, b; Pls. 5e, 32a), the axil being the ‘angle’ between the bract and its stem. Another very common way of expressing this is to say the branches or flowers are ‘subtended by a bract’. Bracts often appear as smaller, reduced versions of the ‘normal’ leaves on the plant, but the term may also be used for structures that are much more obvious and sometimes brightly coloured such as the large white bract of Zantedeschia aethiopica (Arum Lily, Pl. 8d), in this case also sometimes called a spathe.
When examining an inflorescence, it can be helpful to note the relative positions of a flower and the bract below it. In racemose inflorescences a bract will often be found subtending the flower, and is borne on the same side of the axis as the flower. If the branching is cymose, a bract may be seen below, and subtending, the branch that is lateral to the terminal flower (and thus often appears to be on the opposite side of the axis to the flower, as in Figure 11a).
Bracts are not always present, and a number of plant groups produce inflorescences in which they are notably absent—the bracts may be suppressed early in inflorescence development, or be lacking completely.
As often defined, the term bracteole applies to a diminutive bract-like structure borne on the pedicel or calyx of a flower, in which position there is no further growth from its axil. In many plant groups bracteoles are borne in pairs (e.g. Fig. 42c; Pl. 28b). Some authors, however, use the term more broadly in referring to very small bract-like parts in other positions.
Common practice in applying inflorescence terminology
Within plant identification texts, established practice seems to allow a certain degree of flexibility and imprecision with respect to the application of inflorescence terminology. For example, some authors accept arrangements of subsessile flowers (sometimes quite clearly stalked) to fall within the definitions of ‘head’ (e.g. Pl. 16e) and ‘spike’. In these instances the stalks are often obscured by a dense cluster of flowers or bracts. Within some branching inflorescences it can be difficult to establish whether the branches are ‘true’ spikes or are in fact racemes, or the species itself may vary in this feature. Some texts navigate this issue by adopting less definite but still descriptive forms such as ‘spike-like branches’.
Another example is the breadth of application of the term ‘panicle’ to cover a wide variety of branching inflorescences.
In the genus Acacia (Wattle) flowers are often borne in heads, which themselves may then be arranged in more complex inflorescences, commonly referred to as racemes or panicles. This extension of the definitions of raceme and panicle, conventionally defined as an arrangement of flowers, to apply to heads, mirrors that adopted for the Grasses where the basic unit of the inflorescence is the spikelet. In this latter context the definition of the term pedicel is also ‘extended’ to refer to the stalk of the spikelet.
From time to time in specialist literature such as botanical journals, authors are challenging common practice, and proposing new, more accurate ways of understanding and comparing inflorescences, with a repertoire of new terminology. Numerous new terms have been proposed and existing definitions refined, modified or discarded. It remains to be seen how readily a new approach can be incorporated into succinct flora descriptions, or is taken up in more general texts. At present in current English-language floras, apart from one or two new terms seen recently, the inherited conventions appear to have been maintained. A selection of new terms has been included here in the glossary.
Tertiary level botanical text books such as Judd et al. (2016) generally provide an introduction to inflorescence structure. For a more detailed, scholarly account see Weberling (1989).